Specific leaf area estimation model building based on leaf dry matter content of Cunninghamia lanceolata
PENG Xi, YAN Wen-De, WANG Feng-Qi, WANG Guang-Jun, YU Fang-Yong, ZHAO Mei-Fang,*Faculty of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China; Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem in Hunan Province, Huitong, Hunan 418307, China; and National Engineering Laboratory for Applied Forest Ecological Technology in Southern China, Changsha 410004, China
SupportedbytheNationalForestryIndustryResearchSpecialFundsforPublicWelfareProjects(201404316) the National Natural Science Foundation of China.(31600355)
Abstract Aims With progresses of leaf functional traits study, there is an increasing demand to explore the life history strategy and trade-offs in plants, as well as estimate stand productivity, by employing easy and simple leaf parameters. For instance, the interconversion between leaf dry matter content (LDMC) and specific leaf area (SLA) just fit the bill. Cunninghamia lanceolata serves as one of the most important afforestation evergreen needle species in subtropical zone. Building the SLA estimation model based on LDMC could provide a new approach to estimate SLA, and establish a connection path between mechanism explanation and productivity evaluation. Moreover, it could also build a bridge between individual level and large-scale, as well as between actuarial and estimation.Methods Leaf samples were collected from two sampling sites located in C. lanceolata growing region: Huitong County of Hunan Province and Xinyang City of Henan Province. The samples covered fundamentally different niches (aspect, slope position, and canopy depth), and different life history (stand age and leaf age). SLA and LDMC were determined along leaf age gradients, and their value distributions in linkage to different factors were discussed. A general model based on LDMC of C. lanceolata was built to estimate SLA, and the impact of leaf age on the model was explored. Important findings The SLA of C. lanceolata was (103.15 ± 69.54) cm 2·g -1, while LDMC was 0.39 ± 0.11. The LDMC and SLA of C. lanceolata can be estimated by nonlinear model (R2 = 0.718β4, p < 0.001), which meets the estimation requirements. One-year-old leaves showed the best fitting model (R2 = 0.889, p < 0.001), while old leaves (more than 2-year-old) showed the worst (R2 = 0.100β1, p < 0.001). Old leaves with a lower SLA (52.28-75.74 cm 2·g -1) might imply the relative independence among the variation of LDMC. The model based on LDMC to evaluate SLA is credible and effective. The effects on LDMC and SLA along leaf age gradients indicate leaf sensitivity, life history strategies and trade-offs. Keywords:Cunninghamia lanceolata;specific leaf area;leaf dry matter content;model estimation;leaf functional traits
PDF (1146KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 彭曦, 闫文德, 王凤琪, 王光军, 玉昉永, 赵梅芳. 基于叶干质量比的杉木比叶面积估算模型的构建. 植物生态学报, 2018, 42(2): 209-219 doi:10.17521/cjpe.2017.0132 PENG Xi, YAN Wen-De, WANG Feng-Qi, WANG Guang-Jun, YU Fang-Yong, ZHAO Mei-Fang. Specific leaf area estimation model building based on leaf dry matter content of Cunninghamia lanceolata. Chinese Journal of Plant Ecology, 2018, 42(2): 209-219 doi:10.17521/cjpe.2017.0132
本研究试验地设在中国杉木中心产区的湖南会同杉木林生态系统国家野外科学观测研究站(109.75° E, 26.83° N)和非中心产区的河南信阳南湾实验林场(113.97° E, 31.88° N), 两地海拔高度范围分别为270-400 m、300-500 m, 均系低山丘陵地貌。湖南会同与河南信阳同属于亚热带季风气候, 气候温和, 年平均气温15.1-16.8 ℃, 空气湿润, 年平均相对湿度均在77%以上, 雨热同期, 年降水量分别为1β304 mm和1β106 mm, 降水量年内分布不均, 集中在夏季, 日照充沛, 年日照时间均在1β900 h以上, 其光照和水热条件均适宜杉木生长。湖南会同林分密度约为2β400株·hm-2, 河南信阳为1β500-1β800株·hm-2。土壤类型均为山地黄壤。
1.2 研究方法
于杉木生长期4月在湖南会同、河南信阳的不同林龄(幼龄林、中龄林、成熟林、过熟林)各设置2块样方(10 m × 10 m), 共8块。在每个样方内随机选取生长状况良好的3株杉木。为保持叶龄序列, 分别在树冠上层、中层、下层以及冠层以下部位(即临近凋落但未凋落的枝叶)用高枝剪随机采集生长健康、无病虫害的杉木整枝作为标准枝; 同时记录其背景信息如坡位、坡向、海拔等。将采集样品加入湿纸巾放在自封袋内, 用不透光容器保存并尽快带回实验室。
Fig. 1Summary characteristics and degree of discretization of leaf dry matter content (LDMC) (A) and specific leaf area (SLA) (B) of Cunninghamia lanceolata along different factors gradients (n = 374).
Fig. 2Nonlinear fitting model of specific leaf area (SLA) based on leaf dry matter content (LDMC) (A) and SLA scatter plot the regression residuals of LDMC (B) of Cunninghamia lanceolata. Model selection y = a(1 + x)b; N, sampling size; R2, adjusted R-squared; p, significant difference degree.
Fig. 3Nonlinear fitting model of specific leaf area (SLA) based on leaf dry matter content (LDMC) of Cunninghamia lanceolata along different leaf age gradients. Model selection y = a(1 + x)b; N, sampling size; R2, adjusted R-squared; p, significant difference degree.
Fig. 4Specific leaf area (SLA) scatter plot the regression residuals of leaf dry matter content (LDMC) of Cunninghamia lanceolata along different leaf age gradients.
AthokpamFD, GarkotiSC, BorahN ( 2013). Periodicity of leaf growth and leaf dry mass changes in the evergreen and deciduous species of southern Assam, India Ecological Research, 29, 153-165. DOI:10.1007/s11284-013-1105-2URL [本文引用: 1] The study described patterns of leaf dry mass change, leaf mass per area (LMA), relative growth rate and leaf life span (LL) for 14 evergreen and 7 deciduous species of a tropical forest of Southern Assam, India. Leaf expansion in both the groups was, in general, completed before June (i.e. well before the onset of monsoon rains). Although leaf dry mass during leaf initiation phase was significantly higher ( P 02<020.01) in evergreen species than in deciduous species, at the time of full leaf expansion, average leaf dry mass relative to the peak leaf dry mass, realised by the evergreen species was lower (6602%) than for deciduous species (7602%). Leaf dry mass increase in both groups continued after leaf full expansion. Evergreen species had a longer leaf dry mass steady phase than deciduous species (2–6 vs 2–302months). Average LMA of mature leaves for evergreen species (77.4302g02m 612 ) was significantly greater than that of deciduous species (48.4302g02m 612 ). LL ranged from 16502days in Gmelina arborea (deciduous) to 50902days in Dipterocarpus turbinatus (evergreen). LMA was correlated positively with LL, indicating that evergreen species with higher leaf construction cost retain leaves for longer period to pay back. The average leaf dry mass loss before leaf shedding was greater ( P 02<020.01) for deciduous species (30.2902%) than for evergreen species (18.3102%). Although the cost of leaf construction in deciduous species was lower than for evergreen species, they replace leaves at a faster rate. Deciduous species perhaps compensate the cost involved in faster leaf replacement through higher reabsorption of dry mass during senescence, which they remobilise to initiate growth in the following spring when soil resources remain limiting.
BertinN ( 1998). Short and long term fluctuations of the leaf mass per area of tomato plants—Implications for growth models Annals of Botany, 82, 71-81. DOI:10.1006/anbo.1998.0647URL [本文引用: 3] The leaf mass per unit leaf area (LMA) is a key variable in many growth models, since it is often used to predict leaf area expansion from leaf dry weight increase, or vice versa . Influences of source-sink balance on leaf area, leaf dry weight, LMA, and leaf content in non-structural carbohydrates were investigated in glasshouse tomato crops. The source-sink balance was manipulated by artificial shading, CO 2 enrichment or fruit removal using different tomato cultivars. Leaf area was hardly affected by competition for assimilates except under extreme conditions. In contrast, leaf dry weight, and consequently LMA, underwent large and rapid fluctuations in response to any factor that changed source and sink activities. A 60% reduction of photosynthetically active radiation involved a 24% decrease in LMA after 10 d. Carbon dioxide enrichment and fruit removal induced about a 45% and 15% increase in LMA, respectively, on plants with two fruiting trusses, but hardly affected LMA of producing plants. No significant cultivar effect could be identified. Changes in starch and soluble sugar content in leaves accounted for only 29% of diurnal variations in LMA, suggesting regular fluctuations of other components. We propose that structural LMA varies between a maximum and a minimum value according to the ratio of assimilate supply and demand during leaf development. Leaf area is independent of the supply of assimilates when the minimum structural LMA is realised. When the maximum structural LMA is attained, a storage pool of assimilates may accumulate in leaves during periods of high supply and low demand. We present a model including these hypotheses, which predicts structural and non-structural LMA variations of plants with different source-sink ratios.
CantónY, Del BarrioG, Solé-BenetA, LázaroR ( 2004). Topographic controls on the spatial distribution of ground cover in the tabernas badlands of SE Spain Catena, 55, 341-365. DOI:10.1016/S0341-8162(03)00108-5URL [本文引用: 1] The relationships between the spatial distribution of ground-cover and terrain attributes were examined in the Tabernas badlands (SE Spain) in order to understand the terrain-dependent driving forces of the spatially heterogeneous ground cover. Ground cover was mapped in the field and terrain attributes were derived from a 1-m resolution Digital Elevation Model (DEM). The association of spatial distribution of the landforms resulting from a regionalisation (using a nonhierarchical classification of the topographic overlays) and the ground-cover pattern was proved. From the analysis of relationships between terrain attributes and proportional abundance of ground-cover types, it was found that ground cover is arranged along topographic gradients: plant-covered surfaces are more abundant on low slope angles, concave slopes, relatively large contributing areas and with low length slope factor values. Unvegetated surfaces show contrary trends and lichens are associated with intermediate conditions. Relationships with local terrain attributes, such as slope angle or elevation, are more pronounced than those with terrain attributes related to sediment and water transfer, such as contributing area, wetness index or length slope factor which could be explained by the heterogeneity of runoff that is usually shorter than the hillslope length. The relationships established between the spatial distribution of ground-cover types and terrain attributes provide the basis for future development of a tool for mapping spatial distribution of ground cover in similar areas from only topographic information.
CornelissenJHC, LavorelS, GarnierE ( 2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. DOI:10.1071/bt02124URL [本文引用: 3] There is growing recognition that classifying terrestrial plant species on the basis of their function ( into 'functional types') rather than their higher taxonomic identity, is a promising way forward for tackling important ecological questions at the scale of ecosystems, landscapes or biomes. These questions include those on vegetation responses to and vegetation effects on, environmental changes ( e. g. changes in climate, atmospheric chemistry, land use or other disturbances). There is also growing consensus about a shortlist of plant traits that should underlie such functional plant classifications, because they have strong predictive power of important ecosystem responses to environmental change and/or they themselves have strong impacts on ecosystem processes. The most favoured traits are those that are also relatively easy and inexpensive to measure for large numbers of plant species. Large international research efforts, promoted by the IGBP-GCTE Programme, are underway to screen predominant plant species in various ecosystems and biomes worldwide for such traits. This paper provides an international methodological protocol aimed at standardising this research effort, based on consensus among a broad group of scientists in this field. It features a practical handbook with step-by-step recipes, with relatively brief information about the ecological context, for 28 functional traits recognised as critical for tackling large-scale ecological questions.
DangJJ, ZhaoCZ, LiY, HouZJ, DongXG ( 2015). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China. Chinese Journal of Plant Ecology, 39, 23-31. URL [本文引用: 2]
DíazS, CabidoM ( 2009). Plant functional types and ecosystem function in relation to global change Journal of Vegetation Science, 8, 463-474. DOI:10.2307/3237198URL [本文引用: 1] Plant functional types (PFTs) bridge the gap between plant physiology and community and ecosystem processes, thus providing a powerful tool in climate change research. We aimed at identifying PFTs within the flora of central-western Argentina, and to explore their possible consequences for ecosystem function. We analyzed 24 vegetative and regenerative traits of the 100 most abundant species along a steep climatic gradient. Based on plant traits and standard multivariate techniques, we identified eight PFTs. Our results confirmed, over a wide range of climatic conditions, the occurrence of broad recurrent patterns of association among plant traits reported for other floras; namely trade-offs between high investment in photosynthesis and growth on the one hand, and preferential allocation to storage and defence on the other. Regenerative traits were only partially coupled with vegetative traits. Using easily-measured plant traits and individual species cover in 63 sites, we predicted main community-ecosystem processes along the regional gradient. We hypothesized likely impacts of global climatic change on PFTs and ecosystems in situ, and analysed their probabilities of migrating in response to changing climatic conditions. Finally, we discuss the advantages and limitations of this kind of approach in predicting changes in plant distribution and in ecosystem processes over the next century.
DíazS, CabidoM, FernandoC ( 1998). Plant functional traits and environmental filters at a regional scale Journal of Vegetation Science, 9, 113-122. DOI:10.2307/3237229URL [本文引用: 1] Abstract Abstract. Links between plant traits and the environment, i.e. sets of plant attributes consistently associated with certain environmental conditions, are the consequence of the filtering effect of climatic, disturbance and biotic conditions. These filters determine which components of a species pool are assembled into local communities. We aimed at testing for consistent association between plant traits and climatic conditions along a steep regional gradient, divided into 13 climatically homogeneous sectors, in central-western Argentina. We analyzed 19 vegetative and regeneration traits of the 100 most abundant species along the gradient. For each trait, we tested for homogeneity of frequencies of different categories between sectors and the regional species pool, using the 2 statistic. We rejected H 0 in 71% of the pair-wise comparisons, which strongly suggests a iltering effect of climatic factors on key plant functions. Vegetative traits were filtered more often than regeneration traits. Specific leaf area, life span, ramification, canopy height, leaf weight ratio, carbon investment into support tissue and pollination mode were the traits showing differences in the largest number of pair-wise comparisons. This is probably the first attempt to detect, on a quantitative, statistically conservative basis, consistent linkages between climatic factors and numerous plant traits, over a broad spectrum of environmental conditions and plant growth forms. We discuss the advantages and limitations of this approach in predicting vegetation structure and functioning under present environmental conditions, and those expected for the next century as a consequence of global change.
DuanYY, SongLJ, NiuSQ, HuangT, YangGH, HaoWF ( 2017). Variation of leaf functional traits of different aged Robinia pseudoacacia communities and relationships with soil nutrients. Chinese Journal of Applied Ecology, 28, 28-36. URL [本文引用: 1]
EllsworthDS, ReichPB ( 1993). Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest Oecologia, 96, 169-178. DOI:10.1007/BF00317729URLPMID:28313412 [本文引用: 5] Canopy structure and light interception were measured in an 18-m tall, closed canopy deciduous forest of sugar maple ( Acer saccharum ) in southwestern Wisconsin, USA, and related to leaf structural characteristics, N content, and leaf photosynthetic capacity. Light attenuation in the forest occurred primarily in the upper and middle portions of the canopy. Forest stand leaf area index (LAI) and its distribution with respect to canopy height were estimated from canopy transmittance values independently verified with a combined leaf litterfall and point-intersect method. Leaf mass, N and A max per unit area (LMA, N/area and A max /area, respectively) all decreased continuously by over two-fold from the upper to lower canopy, and these traits were strongly correlated with cumulative leaf area above the leaf position in the canopy. In contrast, neither N concentration nor A max per unit mass varied significantly in relation to the vertical canopy gradient. Since leaf N concentration showed no consistent pattern with respect to canopy position, the observed vertical pattern in N/area is a direct consequence of vertical variation of LMA. N/area and LMA were strongly correlated with A max /area among different canopy positions (r 2 =0.81 and r 2 =0.66, respectively), indicating that vertical variation in area-based photosynthetic capacity can also be attributed to variation in LMA. A model of whole-canopy photosynthesis was used to show that observed or hypothetical canopy mass distributions toward higher LMA (and hence higher N/area) in the upper portions of the canopy tended to increase integrated daily canopy photosynthesis over other LMA distribution patterns. Empirical relationships between leaf and canopy-level characteristics may help resolve problems associated with scaling gas exchange measurements made at the leaf level to the individual tree crown and forest canopy-level.
FieldC, MooneyHAJ ( 1983). Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a california shrub Oecologia, 56, 348-355. DOI:10.1007/BF00379711URL [本文引用: 2]
FrakE, RouxXL, MillardP, DreyedE, JaouenG, Saint-JoanisB, WendlerR ( 2001). Changes in total leaf nitrogen and partitioning of leaf nitrogen drive photosynthetic acclimation to light in fully developed walnut leaves Plant, Cell & Environment, 24, 1279-1288. DOI:10.1046/j.0016-8025.2001.00784.xURL [本文引用: 2] Abstract Comprehensive studies on the processes involved in photosynthetic acclimation after a sudden change in light regime are scarce, particularly for trees. We tested (i) the ability of photosynthetic acclimation in the foliage of walnut trees growing outdoors after low-to-high and high-to-low light transfers made early or late in the vegetation cycle, and (ii) the relative importance of changes in total leaf nitrogen versus changes in the partitioning of leaf nitrogen between the different photosynthetic functions during a 2 month period after transfer. Changes in maximum carboxylation rate, light-saturated electron transport rate, respiration rate, total leaf nitrogen, ribulose 1 5-bisphosphate carboxylase/oxygenase (Rubisco) and total chlorophylls were surveyed before and after the change in light regime. Respiration rate acclimated fully within 1 week of transfer, and full acclimation was observed 1 month after transfer for the amount of Rubisco. In contrast, total nitrogen and photosynthetic capacity acclimated only partially during the 2 month period. Changes in photosynthetic capacity were driven by changes in both total leaf nitrogen and leaf nitrogen partitioning. The extent of acclimation also depended strongly on leaf age at the time of the change in light regime.
GaoJL, LuoFM, ZhaoYM, ZhangJB, YuanWJ, SunF, HaoYG ( 2016). Specific leaf area and leaf dry matter content of three kinds of poplar in Ulan Buh desert oasis Journal of Northwest Forestry University, 31(1), 15-20. DOI:10.3969/j.issn.1001-7461.2016.01.03URL [本文引用: 1] 比叶面积(SLA)和叶干物质含量(LDMC)综合反映了植物利用自然资源的能力,也体现了植物对环境的适应性。以乌兰布和沙漠绿洲防护林体系3种杨树(新疆杨、二白杨和小叶杨)为研究对象,通过测定叶面积、叶饱和鲜重及干重,计算了SLA和LDMC,探讨了3种杨树的生态适应性。结果表明:3种杨树叶面积和叶重之间均为极显著线性相关关系(p0.000 1),可用叶面积与叶重的关系式来推算叶面积。其中,用叶面积与叶饱和鲜重的关系式推算的叶面积数据准确度更高。3种杨树的SLA、LDMC均存在极显著差异(p0.0001),新疆杨、二白杨、小叶杨的SLA和LDMC分别为(9.955±2.574)m~2·kg~(-1)和(420.38±68.046)mg·g~(-1),(12.964±2.465)m~2·kg~(-1)和(301.017±32.647)mg·g~(-1),(16.462±6.286)m~2·kg~(-1)和(287.678±72.055)mg·g~(-1)。SLA和LDMC之间呈极显著负相关关系(p0.0001),可用幂函数y=ax~(-b)来表述。SLA和LDMC较好地反映了3种杨树对乌兰布和沙漠贫瘠与干旱环境的适应能力,新疆杨的适应性优于二白杨和小叶杨,建议今后该区域防护林更新与重建过程中应适当加大新疆杨的数量比例。 [ 高君亮, 罗凤敏, 赵英铭, 张景波, 原伟杰, 孙非, 郝玉光 ( 2016). 乌兰布和沙漠绿洲3种杨树比叶面积和叶干物质含量研究 西北林学院学报, 31(1), 15-20.] DOI:10.3969/j.issn.1001-7461.2016.01.03URL [本文引用: 1] 比叶面积(SLA)和叶干物质含量(LDMC)综合反映了植物利用自然资源的能力,也体现了植物对环境的适应性。以乌兰布和沙漠绿洲防护林体系3种杨树(新疆杨、二白杨和小叶杨)为研究对象,通过测定叶面积、叶饱和鲜重及干重,计算了SLA和LDMC,探讨了3种杨树的生态适应性。结果表明:3种杨树叶面积和叶重之间均为极显著线性相关关系(p0.000 1),可用叶面积与叶重的关系式来推算叶面积。其中,用叶面积与叶饱和鲜重的关系式推算的叶面积数据准确度更高。3种杨树的SLA、LDMC均存在极显著差异(p0.0001),新疆杨、二白杨、小叶杨的SLA和LDMC分别为(9.955±2.574)m~2·kg~(-1)和(420.38±68.046)mg·g~(-1),(12.964±2.465)m~2·kg~(-1)和(301.017±32.647)mg·g~(-1),(16.462±6.286)m~2·kg~(-1)和(287.678±72.055)mg·g~(-1)。SLA和LDMC之间呈极显著负相关关系(p0.0001),可用幂函数y=ax~(-b)来表述。SLA和LDMC较好地反映了3种杨树对乌兰布和沙漠贫瘠与干旱环境的适应能力,新疆杨的适应性优于二白杨和小叶杨,建议今后该区域防护林更新与重建过程中应适当加大新疆杨的数量比例。
GarnierE, CordonnierP, GuillermJL ( 1997). Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in mediterranean old-fields Oecologia, 111, 490-498. DOI:10.1007/s004420050262URLPMID:28308109 [本文引用: 1] Specific leaf area (the ratio of leaf area to leaf dry mass) and leaf nitrogen concentration were measured on ten annual and nine perennial grass species growing in two old-fields of southern France, under a sub-humid Mediterranean climate. Specific leaf area (SLA) was found to be significantly higher in annuals than in perennials, but leaf nitrogen concentration expressed on a dry mass basis ($\text{LNC}_{\text{m}}$) was similar in both life-forms; expressed on an area basis, leaf nitrogen concentration ($\text{LNC}_{\text{a}}$) was significantly higher in perennials. The correlation between SLA and$\text{LNC}_{\text{m}}$was negative in annuals and positive in perennials, while that between the inverse of specific leaf area (1/SLA) and$\text{LNC}_{\text{a}}$was positive in annuals and not significant in perennials. It is hypothesized that these contrasting patterns depend on whether the two components of SLA -- leaf thickness and density -- vary in opposite directions. For nine of the species studied (six annuals and three perennials), relative growth rate data obtained in the laboratory under non-limiting nutrient supply were available; positive correlations were found between these values and both SLA and$\text{LNC}_{\text{m}}$obtained in the field, suggesting that the interspecific differences in structural and chemical characteristics of leaves are maintained under a wide range of growing conditions.
GarnierE, LaurentG, BellmannA, DebainS, BerthelierP, DucoutB, RoumetC, NavasML ( 2001 a). Consistency of species ranking based on functional leaf traits New Phytologist, 152, 69-83. DOI:10.1046/j.0028-646x.2001.00239.xURL [本文引用: 1] 61 Specific leaf area (leaf area to dry mass ratio), leaf dry matter content (leaf dry mass to saturated fresh mass ratio) and leaf nitrogen concentration (LNC) have been proposed as indicators of plant resource use in data bases of plant functional traits. 61 We tested whether species ranking based on these traits was repeatable by studying spatio-temporal variations in specific leaf area and leaf dry matter content of water-saturated leaves ( SLASATand LDMCSAT), as well as in LNC, for 57 herbaceous and woody species (or subsets thereof) growing under the Mediterranean climate of southern France. 61 Interseason and intersite variations were more pronounced than interannual variations, but species ranking for a given trait remained mostly consistent in space and time. Classifications based on LDMCSATwere generally more repeatable across years and sites, whereas those based on SLASATwere more stable over seasons. LNC usually gave the least repeatable classifications. 61 Species rankings were not completely similar for the three traits. Discussion of reproducibility, ease of trait measurement, as well as trait-function relationships led us to propose that measurements of the leaf traits, SLASATand/or LDMCSAT, were the most suitable in large screening programmes.
GarnierE, ShipleyB, RoumetC, LaurentG ( 2001 b). A standardized protocol for the determination of specific leaf area and leaf dry matter content Functional Ecology, 15, 688-695. DOI:10.1046/j.0269-8463.2001.00563.xURL [本文引用: 4] 1. The impact of sample preparation, rehydration procedure and time of collection on the determination of specific leaf area (SLA, the ratio of leaf area to leaf dry mass) and leaf dry matter content (LDMC, the ratio of leaf dry mass to fresh mass) of mature leaves was studied in three wild species growing in the field, chosen for their contrasting SLA and LDMC. 2. Complete rehydration was achieved 6 h after samples were placed into water, but neither of the procedures tested - preparation of samples before rehydration or temperature applied during rehydration - had a significant effect on the final values of SLA or LDMC. 3. As expected, water-saturated leaves had a lower LDMC than non-rehydrated leaves; more surprisingly, their SLA was also higher. The impact of rehydration on SLA was especially important when the SLA of the species was high. 4. There was no significant effect of time of sampling on either trait in any species over the time period covered (09.00-16.30 h). 5. These results suggest that SLA and LDMC obtained on water-saturated leaves (SLASATand LDMCSAT) can be used for species comparisons. We propose a standardized protocol for the measurement of these traits. This would allow for better consistency in data collection, a prerequisite for the constitution of large databases of functional traits.
GrataniL, BombelliA ( 2000). Correlation between leaf age and other leaf traits in three mediterranean maquis shrub species:Quercus ilex, Phillyrea latifolia and Cistus incanus. Environmental and Experimental Botany, 43, 141-153. DOI:10.1016/S0098-8472(99)00052-0URL [本文引用: 1] The anatomical and morphological leaf traits as well as leaf inclination and orientation per different leaf age cohort of Quercus ilex, Phillyrea latifolia and Cistus incanus growing in the Mediterranean maquis along Rome’s coast-line (Italy) were investigated. Specific leaf weight (SLW), total leaf thickness ( L), leaf density index and leaf inclination ( α) changed according to leaf age. The maximum values were measured at full leaf expansion, underlining the strong influence of α on the reduction of solar radiation incident on leaf surface and the importance of the received solar radiation by leaf structure during leaf age. C. incanus summer leaves had the lowest surface area, the highest SLW (15±2 mg cm 612) and L (244±15 μm) with respect to winter leaves, reducing the evaporative leaf surface during drought. Older leaves of 2–4 years Q. ilex and P. latifolia, shaded by new leaves had lower α than 1 year old leaves. α is a linear function of SLW. By the seasonal leaf dimorphism and the characteristic leaf folding the adjustment of leaf inclination angle from 6137° in winter leaves to +44° in summer leaves increased reduction of incident solar radiation during drought. Leaf folding may be related to the less xeromorphic leaf structure of C. incanus. The index of xeromorphism, measured at full leaf expansion and resulting from the surface area of the polygon plotted joining the value of the seven considered xeromorphic leaf traits in the radar graph, is the highest in P. latifolia (0.88), and the lowest in C. incanus (0.44).
GreenwoodMS, WardMH, DayME, AdamsSL, BondBJ ( 2008). Age-related trends in red spruce foliar plasticity in relation to declining productivity Tree Physiology, 28, 225-232. DOI:10.1093/treephys/28.2.225URL [本文引用: 1]
GrimeJP, ThompsonK, HuntR, HodgsonJG, CornelissenJHC, RorisonIH, HendryGAF, AshendenTW, AskewAP, BandSR, BoothRE, BossardCC, CampbellBD, CooperJEL, DavisonAW, GuptaPL, HallW, HandDW, HannahMA, HillierSH, HodkinsonDJ, JaliliA, LiuZ, MackeyJML, MatthewsN, MowforthMA, NealAM, ReaderRJ, ReilingK, Ross-FraserW, SpencerRE, SuttonF, TaskerDE, ThorpePC, WhitehouseJ ( 1997). Integrated screening validates primary axes of specialisation in plants Oikos, 79, 259-281. DOI:10.1002/(SICI)1520-6378(200006)25:33.0.CO;2-#URL [本文引用: 1] Standardised procedures have been used to measure 67 traits in 43 common plants of the British flora. This paper provides an interpretation of the most consistent patterns in the resulting matrix by means of correlation, ordination and classification analyses. Only a weak coupling was observed between attributes of the regenerative and established phases of the life history. However, within each phase, attributes were strongly aggregated into sets and a high proportion of the variation between species coincided with a single axis. Attributes of the established phase displayed remarkably consistent trends, with a strong 'Axis 1' being identified by three different multivariate methods. There was a marked correlation between foliar concentrations of N, P, K, Ca and Mg, high concentrations of which coincided with the capacity for rapid growth in productive conditions and an inability to sustain yield under limiting supplies of nutrients. A diverse array of other traits, less immediately involving mineral nutrients, were also entrained in Axis 1; these included life history, root and shoot foraging, the morphology, longevity, tensile strength and palatability of leaves, and the decomposition rate of leaf litter. This pattern occurred in both monocotyledons and dicotyledons and appeared to reflect a tradeoff between attributes conferring an ability for high rates of resource acquisition in productive habitats and those responsible for retention of resource capital in unproductive conditions. The second axis of variation evident in the established phase was related to phylogeny and distinguished between monocotyledons and dicotyledons on the basis of a diverse set of traits including genome size, cell size, root and shoot foraging characteristics and vascular tissues. A third axis was detected in which ephemerals and perennials were separated by differences in attributes such as breeding system, leaf decomposition rate and a set of traits reflecting the small stature of m
HodgsonJG, Montserrat-MartíG, CharlesM, JonesG, WilsonP, ShipleyB, SharafiM, CeraboliniBEL, CornelissenJHC, BandSR, BogardA, Castro-DíezP, Guerrero- CampoJ, PalmerC, Pérez-RontoméMC, CarterG, HyndA, Romo-DíezA, de Torres EspunyL, Royo PlaF ( 2011). Is leaf dry matter content a better predictor of soil fertility than specific leaf area? Annals of Botany, 108, 1337-1345. DOI:10.1093/aob/mcr225URLPMID:3197453 [本文引用: 2] Abstract BACKGROUND AND AIMS: Specific leaf area (SLA), a key element of the 'worldwide leaf economics spectrum', is the preferred 'soft' plant trait for assessing soil fertility. SLA is a function of leaf dry matter content (LDMC) and leaf thickness (LT). The first, LDMC, defines leaf construction costs and can be used instead of SLA. However, LT identifies shade at its lowest extreme and succulence at its highest, and is not related to soil fertility. Why then is SLA more frequently used as a predictor of soil fertility than LDMC? METHODS: SLA, LDMC and LT were measured and leaf density (LD) estimated for almost 2000 species, and the capacity of LD to predict LDMC was examined, as was the relative contribution of LDMC and LT to the expression of SLA. Subsequently, the relationships between SLA, LDMC and LT with respect to soil fertility and shade were described. KEY RESULTS: Although LD is strongly related to LDMC, and LDMC and LT each contribute equally to the expression of SLA, the exact relationships differ between ecological groupings. LDMC predicts leaf nitrogen content and soil fertility but, because LT primarily varies with light intensity, SLA increases in response to both increased shade and increased fertility. CONCLUSIONS: Gradients of soil fertility are frequently also gradients of biomass accumulation with reduced irradiance lower in the canopy. Therefore, SLA, which includes both fertility and shade components, may often discriminate better between communities or treatments than LDMC. However, LDMC should always be the preferred trait for assessing gradients of soil fertility uncoupled from shade. Nevertheless, because leaves multitask, individual leaf traits do not necessarily exhibit exact functional equivalence between species. In consequence, rather than using a single stand-alone predictor, multivariate analyses using several leaf traits is recommended.
HouZJ, ZhaoCZ, LiY, ZhangQ, MaXL ( 2014). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland. Chinese Journal of Plant Ecology, 38, 281-288. DOI:10.3724/SP.J.1258.2014.00025URL [本文引用: 1] 权衡关系是生活史对策理论的基础,株高和枝条数的权衡关系对理解植物在不同生境下的表型可塑性有重要意义。该研究选择祁连山北坡高寒退化草地,利用ArcGIS建立研究区域的数字高程模型(DEM),并提取样地坡向数据,采用广义相加模型(GAM)与偏相关分析相结合的方法,分析了不同坡向影响下狼毒(Stellera chamaejasme)种群株高和枝条数的关系。结果表明:随着坡向由北坡转向东坡、南坡、西坡,草地群落地上生物量和盖度呈"减小—增大—减小"的变化趋势,群落高度则先增大后减小;坡向是影响狼毒株高和枝条数空间分异的主要地形因子;随着坡向由北、东转向西、南,狼毒种群株高呈下降趋势,而枝条数呈上升趋势,二者表现出此消彼长的权衡关系,狼毒植株比叶面积先增大后减小。不同坡向狼毒株高和枝条数的权衡关系,反映了异质生境中资源多重竞争下狼毒生物量分配机制和提高种群适应性的种群更新策略。 [ 侯兆疆, 赵成章, 李钰, 张茜, 马小丽 ( 2014). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系 植物生态学报, 38, 281-288.] DOI:10.3724/SP.J.1258.2014.00025URL [本文引用: 1] 权衡关系是生活史对策理论的基础,株高和枝条数的权衡关系对理解植物在不同生境下的表型可塑性有重要意义。该研究选择祁连山北坡高寒退化草地,利用ArcGIS建立研究区域的数字高程模型(DEM),并提取样地坡向数据,采用广义相加模型(GAM)与偏相关分析相结合的方法,分析了不同坡向影响下狼毒(Stellera chamaejasme)种群株高和枝条数的关系。结果表明:随着坡向由北坡转向东坡、南坡、西坡,草地群落地上生物量和盖度呈"减小—增大—减小"的变化趋势,群落高度则先增大后减小;坡向是影响狼毒株高和枝条数空间分异的主要地形因子;随着坡向由北、东转向西、南,狼毒种群株高呈下降趋势,而枝条数呈上升趋势,二者表现出此消彼长的权衡关系,狼毒植株比叶面积先增大后减小。不同坡向狼毒株高和枝条数的权衡关系,反映了异质生境中资源多重竞争下狼毒生物量分配机制和提高种群适应性的种群更新策略。
HuYS, YaoXY, LiuYH ( 2015). Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains Acta Ecologica Sinica, 35, 1480-1487. DOI:10.5846/stxb201310132459URL [本文引用: 1] 比叶面积(SLA)是植物功能性状的重要指标,反映了植物生长过程中资源收获策略。以长白山森林演替过程中4个阶段的典型群落为研究对象,分析了不同演替阶段比叶面积的差异及影响因素。结果表明:比叶面积随演替的发生进行呈现明显增加趋势,且差异显著(P〈0.05)。相关分析结果表明,比叶面积与海拔、坡位和土壤氮含量均呈显著正相关关系,与其他影响因子无显著相关关系。进一步的通径分析结果表明,演替前期坡位是主要的影响因素,演替中期模型解释度不高,各通径系数较小,演替后期土壤氮含量对比叶面积的作用凸显。 [ 胡耀升, 么旭阳, 刘艳红 ( 2015). 长白山森林不同演替阶段比叶面积及其影响因子 生态学报, 35, 1480-1487.] DOI:10.5846/stxb201310132459URL [本文引用: 1] 比叶面积(SLA)是植物功能性状的重要指标,反映了植物生长过程中资源收获策略。以长白山森林演替过程中4个阶段的典型群落为研究对象,分析了不同演替阶段比叶面积的差异及影响因素。结果表明:比叶面积随演替的发生进行呈现明显增加趋势,且差异显著(P〈0.05)。相关分析结果表明,比叶面积与海拔、坡位和土壤氮含量均呈显著正相关关系,与其他影响因子无显著相关关系。进一步的通径分析结果表明,演替前期坡位是主要的影响因素,演替中期模型解释度不高,各通径系数较小,演替后期土壤氮含量对比叶面积的作用凸显。
HuangHX, YangXD, SunBW, ZhangZH, YanER ( 2014). Variability and association of leaf traits between current-?year and former-year leaves in evergreen trees in Tiantong, Zhejiang, China Chinese Journal of Plant Ecology, 37, 912-921. DOI:10.3724/SP.J.1258.2013.00094URL [本文引用: 2] AimsMethods), specific leaf area () and leaf dry matter content ()) in each of current-year leaves and former-year leaves were measured for 2 277 evergreen trees belonging to 41 species. Variability and correlations for each of three leaf traits were compared between current-year and former-year leaves at both individual and species levels.Important findings) were highest in (individual: 79.5%; species: 66.5%), intermediate in (individual: 28.1%; species: 24.7%) and lowest in (individual: 17.0%; species: 14.1%). Variability coefficients among the three leaf traits were greater in current-year leaves than in former-year leaves. Former-year leaves had significantly greater ( =38.53, ( =9.71, ( = 45.30, , and, and in former-year leaves. More significantly, at the species level 97%, 83% and 85% of the total variation in each of , and, and from current-year leaves. Variation in between differently aged leaves demonstrated that, at a given unit investment of biomass, a relatively larger leaf area can be structured by current-year leaves than by former-year leaves, thus having a low cost in constructing leaf area for current-year leaves. In conclusion, plant leaf traits vary and connect significantly with change of leaf age. Trade-offs between biomass construction and cost in leaf area production might affect plant leaf development. [ 黄海侠, 杨晓东, 孙宝伟, 张志浩, 阎恩荣 ( 2014). 浙江天童常绿植物当年生与往年生叶片性状的变异与关联 植物生态学报, 37, 912-921.] DOI:10.3724/SP.J.1258.2013.00094URL [本文引用: 2] AimsMethods), specific leaf area () and leaf dry matter content ()) in each of current-year leaves and former-year leaves were measured for 2 277 evergreen trees belonging to 41 species. Variability and correlations for each of three leaf traits were compared between current-year and former-year leaves at both individual and species levels.Important findings) were highest in (individual: 79.5%; species: 66.5%), intermediate in (individual: 28.1%; species: 24.7%) and lowest in (individual: 17.0%; species: 14.1%). Variability coefficients among the three leaf traits were greater in current-year leaves than in former-year leaves. Former-year leaves had significantly greater ( =38.53, ( =9.71, ( = 45.30, , and, and in former-year leaves. More significantly, at the species level 97%, 83% and 85% of the total variation in each of , and, and from current-year leaves. Variation in between differently aged leaves demonstrated that, at a given unit investment of biomass, a relatively larger leaf area can be structured by current-year leaves than by former-year leaves, thus having a low cost in constructing leaf area for current-year leaves. In conclusion, plant leaf traits vary and connect significantly with change of leaf age. Trade-offs between biomass construction and cost in leaf area production might affect plant leaf development.
HuoH, WangCK ( 2007). Effects of canopy position and leaf age on photosynthesis and transpiration of Pinus koraiensis. Chinese Journal of Applied Ecology, 18, 1181-1186. URL [本文引用: 1] 利用Li-6400便携式CO2/H2O红外气体分析仪测定了红松不同冠层部位和叶龄针叶的光合蒸腾特性及其环境影响因子.结果表明:冠层部位和叶龄显著地影响最大净光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、表观最大量子效率(α)、蒸腾速率(Tr)和比叶面积(SLA),但对水分利用效率(WUE)影响不显著.随着冠层部位的下降和叶龄的增加,红松针叶的Pmax逐渐下降,其平均值变动在6.55~9.05 μmol·m-2·s-1之间.不同冠层部位和叶龄针叶的LSP和LCP的差异很大,以树冠中部针叶对弱光和强光的利用能力最大.Tr随着冠层部位的下降而降低;不同叶龄针叶的Tr在1.37~1.59 mmol·m-2·s-1之间变化.不同部位和叶龄红松针叶的Tr和光合有效辐射存在极显著正相关关系(R2=0.967).红松的WUE与净光合速率紧密相关(R2=0.860).随冠层部位的上升和叶龄的增大,红松针叶的SLA递减,分别在6.61~8.41 m2·kg-1和6.65~8.38 m2·kg-1之间波动. [ 霍宏, 王传宽 ( 2007). 冠层部位和叶龄对红松光合蒸腾特性的影响 应用生态学报, 18, 1181-1186.] URL [本文引用: 1] 利用Li-6400便携式CO2/H2O红外气体分析仪测定了红松不同冠层部位和叶龄针叶的光合蒸腾特性及其环境影响因子.结果表明:冠层部位和叶龄显著地影响最大净光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、表观最大量子效率(α)、蒸腾速率(Tr)和比叶面积(SLA),但对水分利用效率(WUE)影响不显著.随着冠层部位的下降和叶龄的增加,红松针叶的Pmax逐渐下降,其平均值变动在6.55~9.05 μmol·m-2·s-1之间.不同冠层部位和叶龄针叶的LSP和LCP的差异很大,以树冠中部针叶对弱光和强光的利用能力最大.Tr随着冠层部位的下降而降低;不同叶龄针叶的Tr在1.37~1.59 mmol·m-2·s-1之间变化.不同部位和叶龄红松针叶的Tr和光合有效辐射存在极显著正相关关系(R2=0.967).红松的WUE与净光合速率紧密相关(R2=0.860).随冠层部位的上升和叶龄的增大,红松针叶的SLA递减,分别在6.61~8.41 m2·kg-1和6.65~8.38 m2·kg-1之间波动.
JamesSA, BellDT ( 2000). Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. Globulus provenances. Tree Physiology, 20, 1007-1018. [本文引用: 1]
JiangAP, JiangJM, LiuJ ( 2016). Response of leaf traits of Sassafras tsumu (Hemsl.) Hemsl.along an altitudinal gradient. . Chinese Journal of Ecology, 25, 1467-1474. DOI:10.13292/j.1000-4890.201606.017URL [本文引用: 1] 植物叶片性状是植物与环境长期互作的结果,对环境变化具有敏锐的捕捉力,而且植物叶片性状影响植物的“投资-收益”关系.本文以天目山不同海拔的檫木叶片为材料,分析檫木叶面积、叶长宽比、叶片干重和比叶面积等叶片形态性状以及叶绿素含量和叶氮含量(Narea)等叶片生理性状对海拔的响应.结果表明,不同海拔亚群体间叶面积具有显著差异,比叶面积、叶长宽比和叶片干重存在极显著差异;叶长宽比和叶片干重在亚群体内的变异系数小于叶面积和比叶面积,说明檫木叶片形态性状在亚群体间存在着不同程度的变异.比叶面积与叶面积和叶长宽比具有显著正相关,与叶片干重呈显著负相关;叶绿素含量与Narea呈显著正相关;通过檫木叶片性状与温度的线性回归表明,叶面积、比叶面积、叶绿素含量和Narea随着温度下降而下降,与之相反,叶片干重和叶长宽比则逐渐增加. [ 蒋艾平, 姜景民, 刘军 ( 2016). 檫木叶片性状沿海拔梯度的响应特征 生态学杂志, 25, 1467-1474.] DOI:10.13292/j.1000-4890.201606.017URL [本文引用: 1] 植物叶片性状是植物与环境长期互作的结果,对环境变化具有敏锐的捕捉力,而且植物叶片性状影响植物的“投资-收益”关系.本文以天目山不同海拔的檫木叶片为材料,分析檫木叶面积、叶长宽比、叶片干重和比叶面积等叶片形态性状以及叶绿素含量和叶氮含量(Narea)等叶片生理性状对海拔的响应.结果表明,不同海拔亚群体间叶面积具有显著差异,比叶面积、叶长宽比和叶片干重存在极显著差异;叶长宽比和叶片干重在亚群体内的变异系数小于叶面积和比叶面积,说明檫木叶片形态性状在亚群体间存在着不同程度的变异.比叶面积与叶面积和叶长宽比具有显著正相关,与叶片干重呈显著负相关;叶绿素含量与Narea呈显著正相关;通过檫木叶片性状与温度的线性回归表明,叶面积、比叶面积、叶绿素含量和Narea随着温度下降而下降,与之相反,叶片干重和叶长宽比则逐渐增加.
JullienA, AllirandJ-M, MathieuA, AndrieuB, NeyB ( 2009). Variations in leaf mass per area according to N nutrition, plant age, and leaf position reflect ontogenetic plasticity in winter oilseed rape (Brassica napus l.). Field Crops Research, 114, 188-197. DOI:10.1016/j.fcr.2009.07.015URL [本文引用: 1] Leaf mass per area (LMA) is a parameter the variability of which contributes to architectural plasticity. However its variations according to environment remain not completely understood. We hypothesise that LMA variations of winter oilseed rape (WOSR) follow an ontogenetic trajectory that can be modified by the environment. This was assessed at plant and leaf levels comparing four genotypes field-grown under different environmental conditions characterised by year and nitrogen nutrition. Leaves of the main stem of plants sampled between emergence and harvest were surfaced, oven-dried and weighed to calculate LMA of each individual leaf. At the plant level, LMA presented a common ontogenetic trajectory, which increases from seedling emergence to bolting and from then on decreases. Variations in LMA were related to the variations in plant demand, LMA decreased when demand increased. The trajectory was modified by low N nutrition that increased LMA but differently according to year and genotype, reflecting the plant plasticity. At the leaf level, variations for each individual leaf were related to the variations at the plant level. Plant plasticity and genotypic variability of the responses of LMA to N deficiency seemed to be related to differences in biomass allocation between leaves and stems.
KaravinN ( 2013). Effects of leaf and plant age on specific leaf area in deciduous tree species Quercus cerris L. var. cerris. Bangladesh Journal of Botany, 42, 301-306. DOI:10.3329/bjb.v42i2.18034URL [本文引用: 2] In Quercus cerris L. var. cerris , four diameters at breast height (BDH) groups were selected as a surrogate for approximate plant age groups to study the effects of plant and leaf age on leaf dry mass, leaf area and specific leaf area. The forms of fluctuations in specific leaf area (SLA) of all DBH groups were similar during the growth period. The minimum SLA was calculated at the leaves mature phase. Leaf area did not differ significantly according to leaf age. Significant differences caused by leaf age were determined in mean leaf dry mass and SLA. No significant differences in mean SLA caused by plant age were found. Leaf area and leaf dry mass significantly varied among DBH groups. Leaf area was positively correlated with leaf dry mass, and SLA was negatively correlated with leaf dry mass, while there was no significant relationship with leaf area. DOI: http://dx.doi.org/10.3329/bjb.v42i2.18034 Bangladesh J. Bot. 42 (2): 301-306, 2013 (December)
KattgeJ, DíazS, LavorelS, PrenticeIC, LeadleyP, B?nischG, GarnierE, WestobyM, ReichPB, WrightIJ, CornelissenJHC, ViolleC, HarrisonSP, Van BodegomPM, ReichsteinM, EnquistBJ, SoudzilovskaiaNA, AckerlyDD, AnandM, AtkinO, BahnM, BakerTR, BaldocchiD, BekkerR, BlancoCC, BlonderB, BondWJ, BradstockR, BunkerDE, CasanovesF, Cavender-BaresJ, ChambersJQ, Chapin IIIFS, ChaveJ, CoomesD, CornwellWK, CraineJM, DobrinBH, DuarteL, DurkaW, ElserJ, EsserG, EstiarteM, FaganWF, FangJ, Fernández-MéndezF, FidelisA, FineganB, FloresO, FordH, FrankD, FreschetGT, FyllasNM, GallagherRV, GreenWA, GutierrezAG, HicklerT, HigginsSI, HodgsonJG, JaliliA, JansenS, JolyCA, KerkhoffAJ, KirkupD, KitajimaK, KleyerM, KlotzS, KnopsJMH, KramerK, KühnI, KurokawaH, LaughlinD, LeeTD, LeishmanM, LensF, LenzT, LewisSL, LloydJ, LlusiàJ, LouaultF, MaS, MahechaMD, ManningP, MassadT, MedlynBE, MessierJ, MolesAT, MüllerSC, NadrowskiK, NaeemS, Niinemetsü, N?llertS, NüskeA, OgayaR, OleksynJ, OnipchenkoVG, OnodaY, Ordo?EzJ, OverbeckG, OzingaWA, Pati?oS, PaulaS, PausasJG, Pe?uelasJ, PhillipsOL, PillarV, PoorterH, PoorterL, PoschlodP, PrinzingA, ProulxR, RammigA, ReinschS, ReuB, SackL, Salgado-NegretB, SardansJ, ShioderaS, ShipleyB, SiefertA, SosinskiE, SoussanaJF, SwaineE, SwensonN, ThompsonK, ThorntonP, WaldramM, WeiherE, WhiteM, WhiteS, WrightSJ, YguelB, ZaehleS, ZanneAE, WirthC ( 2011). Try—A global database of plant traits Global Change Biology, 17, 2905-2935. DOI:10.1111/j.1365-2486.2011.02451.xURLPMID:3627314 [本文引用: 1] Plant traits – the morphological, anatomical, physiological, biochemical and phenological characteristics of plants and their organs – determine how primary producers respond to environmental factors, affect other trophic levels, influence ecosystem processes and services and provide a link from species richness to ecosystem function
KattgeJ, KnorrW, RaddatzT, WirthC ( 2009). Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models Global Change Biology, 15, 976-991. DOI:10.1111/j.1365-2486.2008.01744.xURL [本文引用: 1] Abstract Photosynthetic capacity and its relationship to leaf nitrogen content are two of the most sensitive parameters of terrestrial biosphere models (TBM) whose representation in global-scale simulations has been severely hampered by a lack of systematic analyses using a sufficiently broad database. Here, we use data of qualitative traits, climate and soil to subdivide the terrestrial vegetation into functional types (PFT), and then assimilate observations of carboxylation capacity, (723 data points), and maximum photosynthesis rates, (776 data points), into the Cphotosynthesis model proposed by Farquhar to constrain the relationship of (normalised to 25) to leaf nitrogen content per unit leaf area for each PFT. In a second step, the resulting functions are used to predict per PFT from easily measurable values of leaf nitrogen content in natural vegetation (1966 data points). Mean values of thus obtained are implemented into a TBM (BETHY within the coupled climate egetation model ECHAM5/JSBACH) and modelled gross primary production (GPP) is compared with independent observations on stand scale. Apart from providing parameter ranges per PFT constrained from much more comprehensive data, the results of this analysis enable several major improvements on previous parameterisations. (1) The range of mean between PFTs is dominated by differences of photosynthetic nitrogen use efficiency (NUE, defined as divided by leaf nitrogen content), while within each PFT, the scatter of values is dominated by the high variability of leaf nitrogen content. (2) We find a systematic depression of NUE on certain tropical soils that are known to be deficient in phosphorous. (3) of tropical trees derived by this study is substantially lower than earlier estimates currently used in TBMs, with an obvious effect on modelled GPP and surface temperature. (4) The root-mean-squared difference between modelled and observed GPP is substantially reduced.
KayamaM, SasaK, KoikeT ( 2002). Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan. Tree Physiology, 22, 707-716. [本文引用: 1]
LandsbergJJ, GowerST ( 1997). Applications of physiological ecology to forest management Tree Physiology, 17, 601-602. DOI:10.2307/2405244URL [本文引用: 1] This book is aimed at undergraduates in final courses for degrees in forestry and forest ecology, and postgraduate students in the same disciplines. It discusses the effects of the environment, humans and their interactions on the growth of forests, and the use of this knowledge in forest management decision making. There are 10 chapters covering a wide range of topics: Introduction; Forest bio...
LavorelS, GarnierE ( 2002). Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the holy grail Functional Ecology, 16, 545-556. DOI:10.1046/j.1365-2435.2002.00664.xURL [本文引用: 1] Summary 1 The concept of plant functional type proposes that species can be grouped according to common responses to the environment and/or common effects on ecosystem processes. However, the knowledge of relationships between traits associated with the response of plants to environmental factors such as resources and disturbances (response traits), and traits that determine effects of plants on ecosystem functions (effect traits), such as biogeochemical cycling or propensity to disturbance, remains rudimentary. 2 We present a framework using concepts and results from community ecology, ecosystem ecology and evolutionary biology to provide this linkage. Ecosystem functioning is the end result of the operation of multiple environmental filters in a hierarchy of scales which, by selecting individuals with appropriate responses, result in assemblages with varying trait composition. Functional linkages and trade-offs among traits, each of which relates to one or several processes, determine whether or not filtering by different factors gives a match, and whether ecosystem effects can be easily deduced from the knowledge of the filters. 3 To illustrate this framework we analyse a set of key environmental factors and ecosystem processes. While traits associated with response to nutrient gradients strongly overlapped with those determining net primary production, little direct overlap was found between response to fire and flammability. 4 We hypothesize that these patterns reflect general trends. Responses to resource availability would be determined by traits that are also involved in biogeochemical cycling, because both these responses and effects are driven by the trade-off between acquisition and conservation. On the other hand, regeneration and demographic traits associated with response to disturbance, which are known to have little connection with adult traits involved in plant ecophysiology, would be of little relevance to ecosystem processes. 5 This framework is likely to be broadly applicable, although caution must be exercised to use trait linkages and trade-offs appropriate to the scale, environmental conditions and evolutionary context. It may direct the selection of plant functional types for vegetation models at a range of scales, and help with the design of experimental studies of relationships between plant diversity and ecosystem properties.
LiK, XiangWH ( 2011). Comparison of specific leaf area, SPAD value and seed mass among subtropical tree species in hilly area of central Hunan, China Journal of Central South University of Forestry & Technology, 31, 213-218. URL [本文引用: 3]
LiXR, LiuQJ, CaiZ, MaZQ ( 2007). Specific leaf area and leaf area index of conifer plantations in Qianyanzhou station of subtropical China Journal of Plant Ecology (Chinese Version), 31, 93-101. DOI:10.17521/cjpe.2007.0012URL [本文引用: 3] 根据实测数据计算了湿地松(Pinuselliotii)、马尾松(P.massoniana)和杉木(Cunninghamialanceolata)不同年龄、不同类型叶片的生物量和比叶面积,并结合样地调查数据和相对生长方程计算了中国科学院千烟洲试验站20年生湿地松林、马尾松林、杉木林和针叶混交林的叶面积指数。根据拟合结果,选择如下方程计算3个树种的叶生物量湿地松W=12.0741D2.1515、马尾松W=6.9727D2.1973和杉木W=5.2619D2.3027。湿地松林的叶生物量(0.822kg.m-2)最大,其次为针叶混交林(0.679kg.m-2),马尾松林和杉木林相差不大(分别为林0.528和0.572kg.m-2)。不同树种、不同年龄、不同类型叶片的比叶面积比较发现,新叶的比叶面积大于老叶,三针一束叶的比叶面积略大于两针一束叶,马尾松的平均半比表面积(8.62m2.kg-1)大于湿地松(6.04m2.kg-1)和杉木(7.91m2.kg-1)。胸径与单木叶片半表面积之间的经验方程为湿地松LA=0.073D2.1515、马尾松LA=0.060D2.1973和杉木LA=0.042D2.3027。据此计算湿地松林的叶面积指数为5.03,马尾松林和杉木林为4.31,针叶混交林为4.77,该结果比利用CI_110植被冠层数字图像仪测得的结果偏大。 [ 李轩然, 刘琪璟, 蔡哲, 马泽清 ( 2007). 千烟洲针叶林的比叶面积及叶面积指数 植物生态学报, 31, 93-101.] DOI:10.17521/cjpe.2007.0012URL [本文引用: 3] 根据实测数据计算了湿地松(Pinuselliotii)、马尾松(P.massoniana)和杉木(Cunninghamialanceolata)不同年龄、不同类型叶片的生物量和比叶面积,并结合样地调查数据和相对生长方程计算了中国科学院千烟洲试验站20年生湿地松林、马尾松林、杉木林和针叶混交林的叶面积指数。根据拟合结果,选择如下方程计算3个树种的叶生物量湿地松W=12.0741D2.1515、马尾松W=6.9727D2.1973和杉木W=5.2619D2.3027。湿地松林的叶生物量(0.822kg.m-2)最大,其次为针叶混交林(0.679kg.m-2),马尾松林和杉木林相差不大(分别为林0.528和0.572kg.m-2)。不同树种、不同年龄、不同类型叶片的比叶面积比较发现,新叶的比叶面积大于老叶,三针一束叶的比叶面积略大于两针一束叶,马尾松的平均半比表面积(8.62m2.kg-1)大于湿地松(6.04m2.kg-1)和杉木(7.91m2.kg-1)。胸径与单木叶片半表面积之间的经验方程为湿地松LA=0.073D2.1515、马尾松LA=0.060D2.1973和杉木LA=0.042D2.3027。据此计算湿地松林的叶面积指数为5.03,马尾松林和杉木林为4.31,针叶混交林为4.77,该结果比利用CI_110植被冠层数字图像仪测得的结果偏大。
LiYL, CuiJY, SuYZ ( 2005). Specific leaf area and leaf dry matter content of some plants in different dune habitats Acta Ecologica Sinica, 25, 304-311. DOI:10.3321/j.issn:1000-0933.2005.02.019URL [本文引用: 3] 研究了生长在不同沙丘生境中 (流动沙丘 ,半固定沙丘和固定沙丘 ) 2 0个植物种 (10个 1年生植物种和 10个多年生植物种 )的比叶面积 (SL A)和叶干物质含量 (L DMC)的变化 ,并且分析了各个沙丘生境的土壤养分特征。结果表明 ,各个植物种的平均 SL A和 L DMC在植物种之间差异显著 ;多数在两种或 3种沙丘生境均有分布的植物其 SL A在不同沙丘生境之间差异显著 ,但是仅有 6个植物种的 L DMC在不同沙丘生境之间表现出差异 (p0 .0 5 )。与许多研究结果类似 ,1年生植物的 SL A显著大于多年生植物的 SL A,而且两者之间 L DMC存在一定的差异。 1年生植物 SL A和 L DMC之间相关性不显著 ,但多年生植物SL A和 L DMC之间呈显著负相关。综合所有 2 0个植物种可以发现 ,SL A增大时 ,L DMC有下降的趋势 [ 李玉霖, 崔建垣, 苏永中 ( 2005). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较 生态学报, 25, 304-311.] DOI:10.3321/j.issn:1000-0933.2005.02.019URL [本文引用: 3] 研究了生长在不同沙丘生境中 (流动沙丘 ,半固定沙丘和固定沙丘 ) 2 0个植物种 (10个 1年生植物种和 10个多年生植物种 )的比叶面积 (SL A)和叶干物质含量 (L DMC)的变化 ,并且分析了各个沙丘生境的土壤养分特征。结果表明 ,各个植物种的平均 SL A和 L DMC在植物种之间差异显著 ;多数在两种或 3种沙丘生境均有分布的植物其 SL A在不同沙丘生境之间差异显著 ,但是仅有 6个植物种的 L DMC在不同沙丘生境之间表现出差异 (p0 .0 5 )。与许多研究结果类似 ,1年生植物的 SL A显著大于多年生植物的 SL A,而且两者之间 L DMC存在一定的差异。 1年生植物 SL A和 L DMC之间相关性不显著 ,但多年生植物SL A和 L DMC之间呈显著负相关。综合所有 2 0个植物种可以发现 ,SL A增大时 ,L DMC有下降的趋势
LiuZM ( 2014). Research on the Nutrient Dynamics and Internal Absorption of Cunninghamia lanceolata Foliar. Master degree dissertation, Fujian Agriculture and Forestry University Fuzhou. URL [本文引用: 1] 杉木(Cunninghamia lanceolata)是我国南方重要的用材树种,在福建省占有十分重要的地位。近年来随着杉木栽种技术的不断发展,我国杉木人工林经营取得了十分显著的成效。突出表现在单位面积杉木出材率大幅提高,杉木人工林经营管理日趋规范。但是,杉木连栽导致的地力衰退问题也正日益的凸显。杉木养分内循环是杉木养分保持的重要机制,而杉木针叶养分内循环又是杉木养分内循环中非常重要的组成部分。杉木针叶养分内循环的实质是:杉木针叶在衰老的过程中会将大部分的养分元素转移到新鲜的杉木针叶中。这种杉木新老叶之间的养分转移关系会降低杉木营养元素的损失,进而可以提高杉木养分的利用效率。 本研究课题以南平峡阳林场五个年龄段的杉木林为研究对象,将所选标准木的针叶分为四个龄级测定其养分含量,以了解各针叶龄级之间养分的差异及养分转移的动态。旨在揭示同一年龄段杉木针叶各龄级之间的养分状况及转移关系;不同年龄段杉木同一针叶龄级的养分差异;不同年龄段杉木林土壤养分状况等。其主要的研究成果如下: 1.对不同年龄段杉木而言,杉木针叶可转移元素氮、磷、钾、镁含量基本上随着杉木针叶龄级的增加而减少。从养分含量随针叶龄级变化的趋势上看,杉木针叶磷含量的变化最为显著。杉木针叶的氮含量在针叶为当年生和一年生上变化不显著,随着针叶龄级的增加,下降的速度变快;杉木针叶的磷养分在杉木针叶的各个龄级上变化趋势大体一致;杉木针叶的钾含量和镁含量在前两个针叶龄级上变化显著,随着针叶龄级增加,下降的趋势趋于和缓。对不可转移元素钙而言,杉木针叶钙含量随着针叶龄级的增加而增加,钙养分明显的在老叶累积。杉木针叶钙含量在针叶龄级为当年生和一年生上增幅较快,随着针叶龄级的增加,养分累积的速度变慢。 2.杉木林龄与杉木针叶各养分含量的相关关系随针叶龄级而发生明显变化。当杉木针叶龄级为当年生时,杉木针叶的氮含量、磷含量与杉木林龄呈极显著的负相关关系,杉木针叶的钾含量与杉木林龄呈极显著的正相关关系,杉木针叶的氮含量与杉木针叶的钾含量呈极显著的负相关关系,与杉木针叶的磷含量呈显著的正相关关系;当杉木针叶龄级为一年生时,杉木针叶的磷含量、氮含量与杉木林龄呈显著的负相关关系,当杉木针叶龄级为二年生时,杉木针叶氮含量、磷含量与杉木林龄呈显著的负相关关系,杉木针叶钙含量、镁含量与杉木林龄呈显著的正相关关系,杉木针叶氮含量与钙含量呈显著的负相关关系,与杉木针叶的磷含量呈显著的正相关关系,杉木针叶的磷含量与钙含量呈显著的负相关关系,杉木针叶的钾含量与镁含量呈显著的负相关关系;当杉木针叶龄级为三年生时,杉木针叶钾含量、钙含量与杉木叶龄呈显著的正相关关系,杉木针叶磷含量与杉木林龄呈显著的负相关关系,杉木针叶钙含量与磷含量呈显著的负相关关系。 3.对不同年龄杉木林而言,土壤养分含量基本上随着土层的不断加深而逐渐的变小。研究地的土壤养分状况良好,适宜杉木的生长。就各土壤养分的相互关系而言,土壤全氮含量与土壤全磷、全钾、全钙、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤全钾含量与土壤全钙、全镁、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤全钙含量与土壤全镁、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤交换性钙与土壤交换性镁、速效钾含量呈显著的正相关关系;土壤交换性镁与土壤速效钾含量呈显著的正相关关系。 4.不同年龄杉木凋落针叶养分含量在4-7月的比较:杉木凋落针叶氮含量中,14年生杉木21年生杉木8年生杉木46年生杉木;杉木凋落针叶磷含量中,14年生杉木8年生杉木21年生杉木46年生杉木:杉木凋落针叶钾含量中,46年生杉木21年生杉木14年生杉木8年生杉木;杉木凋落针叶镁含量中,21年生杉木14年生杉木46年生杉木8年生杉木;杉木凋落针叶钙含量中,46年生杉木14年生杉木8年生杉木21年生杉木。 5.就杉木养分内吸收率的相互关系而言,杉木针叶各养分内吸收率之间的相关关系不显著。就杉木针叶养分内吸收率的综合比较而言,杉木针叶钾内吸收率杉木针叶磷内吸收率杉木针叶氮内吸收率杉木针叶镁内吸收率杉木针叶钙内吸收率。 [ 刘桌明 ( 2014). 杉木针叶养分含量动态及其内吸收率研究 硕士学位论文, 福建农林大学, 福州.] URL [本文引用: 1] 杉木(Cunninghamia lanceolata)是我国南方重要的用材树种,在福建省占有十分重要的地位。近年来随着杉木栽种技术的不断发展,我国杉木人工林经营取得了十分显著的成效。突出表现在单位面积杉木出材率大幅提高,杉木人工林经营管理日趋规范。但是,杉木连栽导致的地力衰退问题也正日益的凸显。杉木养分内循环是杉木养分保持的重要机制,而杉木针叶养分内循环又是杉木养分内循环中非常重要的组成部分。杉木针叶养分内循环的实质是:杉木针叶在衰老的过程中会将大部分的养分元素转移到新鲜的杉木针叶中。这种杉木新老叶之间的养分转移关系会降低杉木营养元素的损失,进而可以提高杉木养分的利用效率。 本研究课题以南平峡阳林场五个年龄段的杉木林为研究对象,将所选标准木的针叶分为四个龄级测定其养分含量,以了解各针叶龄级之间养分的差异及养分转移的动态。旨在揭示同一年龄段杉木针叶各龄级之间的养分状况及转移关系;不同年龄段杉木同一针叶龄级的养分差异;不同年龄段杉木林土壤养分状况等。其主要的研究成果如下: 1.对不同年龄段杉木而言,杉木针叶可转移元素氮、磷、钾、镁含量基本上随着杉木针叶龄级的增加而减少。从养分含量随针叶龄级变化的趋势上看,杉木针叶磷含量的变化最为显著。杉木针叶的氮含量在针叶为当年生和一年生上变化不显著,随着针叶龄级的增加,下降的速度变快;杉木针叶的磷养分在杉木针叶的各个龄级上变化趋势大体一致;杉木针叶的钾含量和镁含量在前两个针叶龄级上变化显著,随着针叶龄级增加,下降的趋势趋于和缓。对不可转移元素钙而言,杉木针叶钙含量随着针叶龄级的增加而增加,钙养分明显的在老叶累积。杉木针叶钙含量在针叶龄级为当年生和一年生上增幅较快,随着针叶龄级的增加,养分累积的速度变慢。 2.杉木林龄与杉木针叶各养分含量的相关关系随针叶龄级而发生明显变化。当杉木针叶龄级为当年生时,杉木针叶的氮含量、磷含量与杉木林龄呈极显著的负相关关系,杉木针叶的钾含量与杉木林龄呈极显著的正相关关系,杉木针叶的氮含量与杉木针叶的钾含量呈极显著的负相关关系,与杉木针叶的磷含量呈显著的正相关关系;当杉木针叶龄级为一年生时,杉木针叶的磷含量、氮含量与杉木林龄呈显著的负相关关系,当杉木针叶龄级为二年生时,杉木针叶氮含量、磷含量与杉木林龄呈显著的负相关关系,杉木针叶钙含量、镁含量与杉木林龄呈显著的正相关关系,杉木针叶氮含量与钙含量呈显著的负相关关系,与杉木针叶的磷含量呈显著的正相关关系,杉木针叶的磷含量与钙含量呈显著的负相关关系,杉木针叶的钾含量与镁含量呈显著的负相关关系;当杉木针叶龄级为三年生时,杉木针叶钾含量、钙含量与杉木叶龄呈显著的正相关关系,杉木针叶磷含量与杉木林龄呈显著的负相关关系,杉木针叶钙含量与磷含量呈显著的负相关关系。 3.对不同年龄杉木林而言,土壤养分含量基本上随着土层的不断加深而逐渐的变小。研究地的土壤养分状况良好,适宜杉木的生长。就各土壤养分的相互关系而言,土壤全氮含量与土壤全磷、全钾、全钙、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤全钾含量与土壤全钙、全镁、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤全钙含量与土壤全镁、交换性钙、交换性镁、速效钾含量有显著的正相关关系;土壤交换性钙与土壤交换性镁、速效钾含量呈显著的正相关关系;土壤交换性镁与土壤速效钾含量呈显著的正相关关系。 4.不同年龄杉木凋落针叶养分含量在4-7月的比较:杉木凋落针叶氮含量中,14年生杉木21年生杉木8年生杉木46年生杉木;杉木凋落针叶磷含量中,14年生杉木8年生杉木21年生杉木46年生杉木:杉木凋落针叶钾含量中,46年生杉木21年生杉木14年生杉木8年生杉木;杉木凋落针叶镁含量中,21年生杉木14年生杉木46年生杉木8年生杉木;杉木凋落针叶钙含量中,46年生杉木14年生杉木8年生杉木21年生杉木。 5.就杉木养分内吸收率的相互关系而言,杉木针叶各养分内吸收率之间的相关关系不显著。就杉木针叶养分内吸收率的综合比较而言,杉木针叶钾内吸收率杉木针叶磷内吸收率杉木针叶氮内吸收率杉木针叶镁内吸收率杉木针叶钙内吸收率。
MatsuzakiJ, NorisadaM, KodairaJ, SuzukiM, TangeT (2004). Shoots grafted into the upper crowns of tall Japanese cedar ( Cryptomeria japonica D. Don) show foliar gas exchange characteristics similar to those of intact shoots. Trees, 19, 198-203. [本文引用: 1]
MencucciniM, Martínez-VilaltaJ, VanderkleinD, HamidHA, KorakakiE, LeeS, MichielsB ( 2005). Size-mediated ageing reduces vigour in trees Ecology Letter, 8, 1183-1190. DOI:10.1111/j.1461-0248.2005.00819.xURLPMID:21352442 [本文引用: 1] There is increasing interest in understanding the costs and benefits of increased size and prolonged lifespan for plants. Some species of trees can grow more than 100 m in height and can live for several millennia, however whether these achievements are obtained at the cost of some other physiological functions is currently unclear. As increases in size are usually associated with ageing, it is also unclear whether observed reductions in growth rates and increased mortality rates are a function of size or of age per se. One theory proposes that reduced growth after the start of the reproductive phase is caused by cellular senescence. A second set of theories has focussed instead on plant size and the increased respiratory burdens or excessive height. We report on experimental manipulations to separate the effects of extrinsic factors such as size from those of intrinsic factors such as age for four tree species of contrasting phylogeny and life history. For each species, we measured growth, gas exchange and leaf biochemical properties for trees of different ages and sizes in the field and on propagated material obtained from the same genetic individuals but now all of small similar size in our common gardens. For all species, evidence indicated that size, not cellular senescence, accounted for the observed age-related declines in relative growth rates and net assimilation rates. Two species exhibited evidence of genetic control on leaf characters such as specific leaf area, although size also exerted an independent, and stronger, effect. We found partial support for the theory of hydraulic limitations to tree growth. The lack of a marked separation of soma and germline, an unlimited proliferation potential of meristem cells and the exponential increase in reproductive effort with size all help explain the lack of a senescence-induced decline in trees. It is possible that trees much older than the ones we sampled exhibit senescence symptoms.
MiaoYM, LüJZ, BiRC ( 2012). Distribution pattern of leaf traits in canopy of typical broad-leaved trees in Taiyue Mountain Guihaia, 32, 483-486. URL [本文引用: 1]
NicotraAB, LeighA, BoyceCK, JonesCS, NiklasKJ, RoyerDL, TsukayaH ( 2011). The evolution and functional significance of leaf shape in the angiosperms Functional Plant Biology, 38, 535-552. DOI:10.1071/FP11057URL [本文引用: 1]
OnoK, NishiY, WatanabeA, TerashimaI ( 2001). Possible mechanisms of adaptive leaf senescence Plant Biology, 3, 234-243. DOI:10.1055/s-2001-15201URL [本文引用: 1] Abstract: Availability of nitrogen almost always limits plant growth. Therefore, efficient use of nitrogen is essential for the plants. In upright plants, especially when they form dense plant stands, old, lower leaves are shaded by young, upper leaves. Nitrogenous compounds in such shaded leaves are degraded and re-allocated to the developing young, upper leaves. These processes raise efficiency of nitrogen use in photosynthetic production of the plant. For this to occur in the most effective way, leaves would need to sense their photosynthetic status in a plant and increase, maintain or decrease their photosynthetic capacity accordingly. Hypotheses that explain how a leaf can sense its photosynthetic status in the plant are reviewed. They include systems involving phytochrome, sugar-sensing, or cytokinin. Our experimental results with Helianthus annuus and Phaseolus vulgaris plants, which were subject to various shading treatments, are examined in the light of these hypotheses. Our experimental results favoured the sugar-sensing hypothesis: A leaf can sense demand of other plant parts for photosynthates produced by it and nitrogen abundance or deficiency by monitoring its sugar concentration. Problems that are to be challenged in the near future are also pointed out.
OsnasJL, LichsteinJW, ReichPB, PacalaSW ( 2013). Global leaf trait relationships: Mass, area, and the leaf economics spectrum Science, 340, 741-744. DOI:10.1126/science.1231574URL [本文引用: 1]
OuXL, LiuYH ( 2017). Age, slope aspectsand diameter classes affect the leaf functional traits of Pinus tabulaeformis in Songshan, Beijing. Journal of Nanjing Forestry University (Natural Sciences Edition), 41(4), 80-88. [本文引用: 1]
QinXH ( 2015). Spatial heterogeneity of specific leaf area and leaf construction cost of Quercus liaotungensis canopy. Forest Resources Management, ( 4), 145-150. URL [本文引用: 1]
ReichPB, UhlC, WaltersMB, EllsworthDS (1991a). Leaf lifespan as a determinant of leaf structure and function among 23 amazonian tree species Oecologia, 86, 16-24. DOI:10.1007/BF00317383URLPMID:28313152 [本文引用: 2] The relationships between resource availability, plant succession, and species' life history traits are often considered key to understanding variation among species and communities. Leaf lifespan is one trait important in this regard. We observed that leaf lifespan varies 30-fold among 23 species from natural and disturbed communities within a 1-km radius in the northern Amazon basin, near San Carlos de Rio Negro, Venezuela. Moreover, leaf lifespan was highly correlated with a number of important leaf structural and functional characteristics. Stomatal conductance to water vapor (g) and both mass and area-based net photosynthesis decreased with increasing leaf lifespan (r2= 0.74, 0.91 and 0.75, respectively). Specific leaf area (SLA) also decreased with increasing leaf lifespan (r2= 0.78), while leaf toughness increased (r2= 0.62). Correlations between leaf lifespan and leaf nitrogen and phosphorus concentrations were moderate on a weight basis and not significant on an area basis. On an absolute basis, changes in SLA, net photosynthesis and leaf chemistry were large as leaf lifespan varied from 1.5 to 12 months, but such changes were small as leaf lifespan increased from 1 to 5 years. Mass-based net photosynthesis (A/mass) was highly correlated with SLA (r2= 0.90) and mass-based leaf nitrogen (N/mass) (r2= 0.85), but area-based net photosynthesis (A/area) was not well correlated with any index of leaf structure or chemistry including N/area. Overall, these results indicate that species allocate resources towards a high photosynthetic assimilation rate for a brief time, or provide resistant physical structure that results in a lower rate of carbon assimilation over a longer time, but not both.
ReichPB, WaltersMB, EllsworthDS (1991 b). Leaf age and season influence the relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees Plant, Cell & Environment, 14, 251-259. [本文引用: 3]
ReichPB, WaltersMB, EllsworthDS ( 1992). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems Ecological Monographs, 62, 365-392. DOI:10.2307/2937116URL [本文引用: 1] Variation in leaf life-span has long been considered of ecological significance.Despite this, quantitative evaluation of the relationships between leaf life-span and other plant and ecosystem characteristics has been rare. In this paper we ask whether leaf life-span is related to other leaf, plant, and stand traits of species from diverse ecosystems and biomes. We also examine the interaction between leaf, plant, and stand traits and their relation to productivity and ecological patterns. Among all species, both mass- (A"m"a"s"s) and area-based (A"a"r"e"a) maximum net photosynthesis decreased with increasing leaf life-span, but the relationship was stronger on a mass (P <.001, r^2 = 0.70) than an area (P <.05, r^2 = 0.24) basis. Similarly, mass-based leaf nitrogen (leaf N"m"a"s"s) decreased (P <.001, r^2 = 0.52) with leaf life-span, but area-based leaf N (leaf N"a"r"e"a) did not (P >.25, r^2 = 0.01). Specific leaf area (SLA, leaf area/leaf dry mass) and leaf diffusive conductance also decreased with increasing leaf life-span. Decreasing A"m"a"s"s with increasing leaf life-span results from the impact of decreasing N"m"a"s"s and SLA on A"m"a"s"s. Variation in leaf traits as a function of leaf life-span was similar for broad-leaved and needle-leaved subsets of the data. These leaf-scale data from several biomes were compared to a data set from a single biome, Amazonia. For several leaf traits (e.g., SLA, N"m"a"s"s, and A"m"a"s"s) the quantitative relationship with leaf life-span was similar in the two independent data sets, suggesting that these are fundamental relations applicable to all species. A"m"a"s"s was a linear function of N"m"a"s"s (P .001, r^2 = 0.74) with a regression similar to previous analyses, while A"a"r"e"a was not significantly related to N"a"r"e"a. These results suggest that the photosynthesis-leaf N relationship among species should be considered universal when expressed on a mass, but not on a leaf area, basis. Relative growth rates (RGR) and leaf area ratio (LAR, the whole-plant ratio of leaf area to total dry mass) of seedlings decreased with increasing leaf life-span (P < .001, r^2 = 0.61 and 0.89, respectively). LAR was positively related to both RGR and A"m"a"s"s (r^2 = 0.68 and 0.84, respectively), and A"m"a"s"s and RGR were also positively related (r^2 = 0.55). Absolute height growth rates of young trees decreased with increasing leaf life-span (P < .001, r^2 = 0.72) and increased with A"m"a"s"s (P < .001, r^2 = 0.78). It appears that a suite of traits including short leaf life-span and high leaf N"m"a"s"s, SLA, LAR, and A"m"a"s"s interactively contribute to high growth rates in open-grown individuals. These traits interact similarly at the stand level, but stands differ from individuals in one key trait. In closed-canopy forests, species with longer lived foliage (and low LAR as seedlings) have greater foliage mass per unit ground area (P < .001, r^2 = 0.74) and a greater proportion of total mass in foliage. The aboveground production efficiency (ANPP/foliar biomass) of forest stands decreased markedly with increasing leaf life-span or total foliage mass (P < .001, r^2 = 0.78 and 0.72, respectively), probably as a result of decreasing A"m"a"s"s, N"m"a"s"s, and SLA, all of which were positively related with production efficiency and negatively related to total foliage mass. However, high foliage mass of species with extended leaf life-spans appears to compensate for low production per unit foliage, since aboveground net primary production (ANPP, in megagrams per hectare per year) of forest stands was not related to leaf life-span. Extended leaf life-span also appears to compensate for lower potential production per unit leaf N per unit time, with the result that stand-level N use efficiency is weakly positively related to leaf life-span. We hypothesize that co-variation among species in leaf life-span, SLA, leaf N"m"a"s"s, A"m"a"s"s, and growth rate reflects a set of mutually supporting traits that interact to determine plant behavior and production, and provide a useful conceptual link between processes at short-term leaf scales and longer term whole plant and stand-level scales. Although this paper has focused on leaf life-span, this trait is so closely interrelated with several others that this cohort of leaf traits should be viewed as casually interrelated. Generality in the relationships between leaf life-span and other plant traits across diverse communities and ecosystems suggests that they are universal in nature and thus can provide a quantitative link and/or common currency for ecological comparisons among diverse systems.
ReichPB, WaltersMB, EllsworthDS, VoseJM, VolinJC, GreshamC, BowmanWD ( 1998). Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups Oecologia, 114, 471-482. DOI:10.1007/s004420050471URLPMID:28307896 [本文引用: 1] Based on prior evidence of coordinated multiple leaf trait scaling, we hypothesized that variation among species in leaf dark respiration rate ( R d ) should scale with variation in traits such as leaf nitrogen (N), leaf life-span, specific leaf area (SLA), and net photosynthetic capacity ( A max ). However, it is not known whether such scaling, if it exists, is similar among disparate biomes and plant functional types. We tested this idea by examining the interspecific relationships between R d measured at a standard temperature and leaf life-span, N, SLA and A max for 69 species from four functional groups (forbs, broad-leafed trees and shrubs, and needle-leafed conifers) in six biomes traversing the Americas: alpine tundra/subalpine forest, Colorado; cold temperate forest/grassland, Wisconsin; cool temperate forest, North Carolina; desert/shrubland, New Mexico; subtropical forest, South Carolina; and tropical rain forest, Amazonas, Venezuela. Area-based R d was positively related to area-based leaf N within functional groups and for all species pooled, but not when comparing among species within any site. At all sites, mass-based R d ( R d-mass ) decreased sharply with increasing leaf life-span and was positively related to SLA and mass-based A max and leaf N (leaf N mass ). These intra-biome relationships were similar in shape and slope among sites, where in each case we compared species belonging to different plant functional groups. Significant R d-mass 61 N mass relationships were observed in all functional groups (pooled across sites), but the relationships differed, with higher R d at any given leaf N in functional groups (such as forbs) with higher SLA and shorter leaf life-span. Regardless of biome or functional group, R d-mass was well predicted by all combinations of leaf life-span, N mass and/or SLA ( r 2 ≥ 0.79, P65 <650.0001). At any given SLA, R d-mass rises with increasing N mass and/or decreasing leaf life-span; and at any level of N mass , R d-mass rises with increasing SLA and/or decreasing leaf life-span. The relationships between R d and leaf traits observed in this study support the idea of a global set of predictable interrelationships between key leaf morphological, chemical and metabolic traits.
RenY, LuQ, WuB, LiMH ( 2015). Specific leaf area and leaf dry matter content of Nitraria tangutorum in the artificially simulated precipitation. Acta Ecologica Sinica, 35, 4707-4715. URL [本文引用: 1]
RocheP, Díaz-BurlinsonN, GachetS ( 2004). Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecology, 174, 37-48. DOI:10.1023/B:VEGE.0000046056.94523.57URL [本文引用: 2] Nine leaf traits (area, fresh weight, dry weight, volume, density, thickness, specific leaf area (SLA), dry matter content (LDMC), leaf nitrogen content (LNC)) from ten plant species at eight sites in southern mediterranean France were investigated in order to assess their variability along a climatic gradient and their ranking congruency power. After examination of trait correlation patterns, we reduced the nine initial leaf traits to four traits, representative of three correlation groups: allometric traits (dry weight), functional traits (SLA and dry matter percentage) and Leaf Thickness. We analysed the variability of these four leaf traits at species and site level. We observed that between species variation (between 64.5 for SLA and 91% for LDMC) is higher than within species variation. Allowing a good congruency of species ranking assessed by spearman rank correlation () and a good reallocation of individuals to species by discriminant analysis. A site level variability (between 0.7% for Dry weight and 6.9% for SLA) was identified and environmental parameters (altitude, temperature, precipitation, nitrogen, pH) were considered as probable control factors. We found significant correlation between SLA, LDMC and the average minimum temperature (respectively r=0.87 and r=0,9) and no correlation for the other traits or environmental parameters. Furthermore, we conclude that two leaf traits appear to be central in describing species: specific leaf area (SLA), percentage of dry matter (LDMC. While, SLA and LDMC are strongly correlated, LDMC appears to be less variable than SLA. According to our results the Dry Matter Content (or its reversal Leaf Water Content) appears the best leaf trait to be quantified for plant functional screening. Leaf thickness appeared to be rather uncorrelated with other leaf traits and show no environmental contingency; its variability could not have been explained in this study. Further studies should focus on this trait.
ShiYC, ZhaoCZ, SongQH, DuJ, ChenJ, WangJW ( 2015). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of Lanzhou. Chinese Journal of Plant Ecology, 39, 362-370. [本文引用: 1]
ShipleyB, LechowiczMJ, WrightIJ ( 2006). Fundamental trade-offs generating the worldwide leaf economics spectrum Ecology, 87, 535-541. DOI:10.1890/05-1051URL [本文引用: 1]
SimsDA, PearcyRW ( 1992). Response of leaf anatomy and photosynthetic capacity in Alocasia macrorrhiza(Araceae) to a transfer from low to high light. American Journal of Botany, 79, 449-455. [本文引用: 1]
SmartSM, GlanvilleHC, BlanesMDC, MercadoLM, EmmettBA, JonesDL, CosbyBJ, MarrsRH, ButlerA, MarshallMR, ReinschS, Herrero-JáureguiC, HodgsonJG, FieldK ( 2017). Leaf dry matter content is better at predicting above-ground net primary production than specific leaf area Functional Ecology, 31, 1336-1344. DOI:10.1111/1365-2435.12832URL [本文引用: 3] 1. Reliable modelling of above-ground net primary production (aNPP) at fine resolution is a significant challenge. A promising avenue for improving process models is to include response and effect trait relationships. However, uncertainties remain over which leaf traits are correlated most strongly with aNPP. 2. We compared abundance-weighted values of two of the most widely used traits from the leaf economics spectrum (specific leaf area and leaf dry matter content) with measured aNPP across a temperate ecosystem gradient. 3. We found that leaf dry matter content (LDMC) as opposed to specific leaf area (SLA) was the superior predictor of aNPP (R2 = 0 55). 4. Directly measured in situ trait values for the dominant species improved estimation of aNPP significantly. Introducing intraspecific trait variation by including the effect of replicated trait values from published databases did not improve the estimation of aNPP. 5. Our results support the prospect of greater scientific understanding for less cost because LDMC is much easier to measure than SLA.
SterckFJ, PoorterL, SchievingF ( 2006). Leaf traits determine the growth-survival trade-off across rain forest tree species The American Naturalist, 167, 756-765. DOI:10.1086/503056URLPMID:16671019 [本文引用: 1] A dominant hypothesis explaining tree species coexistence in tropical forest is that trade‐offs in characters allow species to adapt to different light environments, but tests for this hypothesis are scarce. This study is the first that uses a theoretical plant growth model to link leaf trade‐offs to whole‐plant performances and to differential performances across species in different light environments. Using data of 50 sympatric tree species from a Bolivian rain forest, we observed that specific leaf area and photosynthetic capacity codetermined interspecific height growth variation in a forest gap; that leaf survival rate determined the variation in plant survival rate under a closed canopy; that predicted height growth and plant survival rate matched field observations; and that fast‐growing species had low survival rates for both field and predicted values. These results show how leaf trade‐offs influence differential tree performance and tree species' coexistence in a heterogeneous light environment.
TorrezV, J?rgensenPM, ZanneAE ( 2013). Specific leaf area: A predictive model using dried samples Australian Journal of Botany, 61, 350-357. DOI:10.1071/BT12236URL [本文引用: 1] Specific leaf area (SLA; fresh-leaf area/dry mass) describes the amount of leaf area for light capture per unit of biomass invested. The standard protocol is simple; however, it requires recently collected sun-exposed leaves to determine fresh-leaf area, limiting where and which samples can be studied. A protocol to predict SLA for fresh leaves from herbarium-dried leaves was developed from samples collected in a dry forest in Bolivia. Leaf area was measured both fresh and dried on the same leaf samples to generate two general mixed-effects models, varying in their inclusion of the position in the crown where the leaf developed. As a test of the potential generality of the models for other systems, we applied them to samples collected in an oak-hickory forest in Missouri, USA. Both models performed well. A recommended protocol for studies predicting SLA from dry leaves was developed. These predictive models and protocols can extend the temporal, geographic, ecological and taxonomic scope of SLA studies.
WangXH, ZhangJ, ZhangZX ( 2000). Leaf longevity of evergreen broad-leaved species of Tiantong National Forest Park, Zhejiang Province Acta Phytoecologica Sinica, 24, 625-629. DOI:10.1088/0256-307X/17/9/008URL [本文引用: 1] 用生命表法对天童国家森林公园主要常绿阔叶树种叶子的寿命进行了研究,结果表明:大多数常绿阔叶树种叶子的寿命在1-3年之间,且变化幅度较大,就平均值来看,小乔木及灌木的叶子寿命较长,中乔木叶子的寿命较短;在演替过程中,组成演替早期群落的物种叶子寿命较短,而后期群落的植物叶子的寿命较长。文中还就叶子寿命和常绿阔叶林的结构及动态机理进行了讨论。 [ 王希华, 张婕, 张正祥 ( 2000). 浙江天童国家森林公园主要常绿阔叶树种叶子寿命的研究 植物生态学报, 24, 625-629.] DOI:10.1088/0256-307X/17/9/008URL [本文引用: 1] 用生命表法对天童国家森林公园主要常绿阔叶树种叶子的寿命进行了研究,结果表明:大多数常绿阔叶树种叶子的寿命在1-3年之间,且变化幅度较大,就平均值来看,小乔木及灌木的叶子寿命较长,中乔木叶子的寿命较短;在演替过程中,组成演替早期群落的物种叶子寿命较短,而后期群落的植物叶子的寿命较长。文中还就叶子寿命和常绿阔叶林的结构及动态机理进行了讨论。
WestobyM, FalsterDS, MolesAT, VeskPA, WrightIJ ( 2002). Plant ecological strategies: Some leading dimensions of variation between species Annual Review of Ecology and Systematics, 33, 125-159. DOI:10.1146/annurev.ecolsys.33.010802.150452URL [本文引用: 2]
WilsonPJ, ThompsonKEN, HodgsonJG ( 1999). Specific leaf area and leaf dry matter content as alternative predictors of plant strategies New Phytologist, 143, 155-162. DOI:10.1046/j.1469-8137.1999.00427.xURL [本文引用: 6] Abstract A key element of most recently proposed plant strategy schemes is an axis of resource capture, usage and availability. In the search for a simple, robust plant trait (or traits) that will allow plants to be located on this axis, specific leaf area is one of the leading contenders. Using a large new unpublished database, we examine the variability of specific leaf area and other leaf traits, the relationships between them, and their ability to predict position on the resource use axis. Specific leaf area is found to suffer from a number of drawbacks; it is both very variable between replicates and much influenced by leaf thickness. Leaf dry-matter content (sometimes referred to as tissue density) is much less variable, largely independent of leaf thickness and a better predictor of location on an axis of resource capture, usage and availability. However, it is not clear how useful dry matter content will be outside northwest Europe, and in particular in dry climates with many succulents.
WirthC, LichsteinJW ( 2009). The imprint of species turnover on old-growth forest carbon balances—Insights from a trait-based model of forest dynamics Old-Growth Forests, 81-113 DOI:10.1007/978-3-540-92706-8_5URL [本文引用: 1] This chapter presents a modelling approach to quantify and understand successional patterns in aboveground carbon stock changes, with a focus on the old-growth stage. The model integrates extensive da
WitkowskiETF, LamontBB ( 1991). Leaf specific mass confounds leaf density and thickness Oecologia, 88, 486-493. DOI:10.1007/BF00317710URLPMID:28312617 [本文引用: 2] We explored the relationship between leaf specific mass (LSM) and its two components, leaf density and thickness. These were assessed on the leaves of (a) the moderately sclerophyllous tree Arbutus menziesii distributed along a natural nutrient/moisture gradient in California, (b) eight sclerophyllous shrub species on four substrates in south-western Australia, and (c) seedlings of two morphologically contrasting Hakea species grown under varying soil nutrient, moisture and light regimes in a glasshouse experiment. Leaf area, mass, LSM, density and thickness varied greatly between leaves on the same plant, different species, and with different nutrient, moisture and light regimes. In some cases, variations in LSM were due to changes in leaf density in particular or thickness or both, while in others, density and thickness varied without a net effect on LSM. At lower nutrient or moisture availabilities or at higher light irradiances, leaves tended to be smaller, with higher LSM, density and thickness. Under increased stress, the thickness (diameter) of needle leaves decreased despite an increase in LSM. We concluded that, while LSM is a useful measure of sclerophylly, its separation into leaf density and thickness may be more appropriate as they often vary independently and appear to be more responsive to environmental gradients than LSM.
WykaT, RobakowskiP, ZytkowiakR ( 2008). Leaf age as a factor in anatomical and physiological acclimative responses of Taxus baccata L. needles to contrasting irradiance environments. Photosynthesis Research, 95, 87-99. [本文引用: 2]
YangM, WangSL, ZhangWD, WangQK ( 2010). Dynamics of biomass and nutrient accumulation in a Chinese-fir plantation Chinese Journal of Applied Ecology, 21, 1674-1680. URL [本文引用: 1]
YuYF, SongTQ, ZengFP, PengWX, WenYG, HuangCB, WuQB, ZengZX, YuY ( 2013). Dynamics changes of biomass and its allocation in Cunninghamia lanceolata plantations of different stand ages. Chinese Journal of Ecology, 32, 1660-1666. URL [本文引用: 1]
ZaehleS, FriendAD ( 2010). Carbon and nitrogen cycle dynamics in the O-CN land surface model: 1. Model description, site-scale evaluation, and sensitivity to parameter estimates Global Biogeochemical Cycles, 24, 1468-1470. DOI:10.1029/2009GB003521URL [本文引用: 1] [1] Nitrogen (N) availability plays a key role in terrestrial biosphere dynamics. To understand and quantify the role of terrestrial N in the Earth system, we developed an advanced terrestrial biogeochemical model O-CN that mechanistically couples terrestrial energy, water, carbon, and nitrogen fluxes in terrestrial ecosystems. We evaluate this new model against observations from intensive forest monitoring plots at temperate and boreal locations in Europe. O-CN simulates realistic foliage N concentrations and N cycling rates and reproduces observed diurnal and seasonal cycles of C fluxes as well as observed gradients in vegetation productivity with N availability for the forest sites studied. A sensitivity test reveals that these results are reasonably robust against uncertainties in model parameter estimates. Using this model we quantify the likely contribution of anthropogenic N deposition to present ecosystem C sequestration as 36 (range: 209000979) g C g0908081 N in agreement with ecosystem manipulation studies.
ZhangL, LuoTX, DengKM, LiWH ( 2008). Vertical variations in specific leaf and leaf dry matter content with canopy height in Pinus yunnanensis. Journal of Beijing Forestry University, 30(1), 40-44. DOI:10.3321/j.issn:1000-1522.2008.01.007URL [本文引用: 3] 比叶面积(SLA)和叶干物质含量(LDMC)综合反应了植物利用资源的能力,是植物适应环境所体现出的关键叶性状。为深入了解SLA和LDMC沿冠层高度的垂直变化规律,该文对云南省永仁县万马乡2株云南松(树高14.4m,树龄29年;树高30.2m,树龄138年)不同冠层高度叶片SLA和LDMC进行调查研究,分析了不同年龄树木、不同叶龄SLA和LDMC的差异,初步探讨了SLA和LDMC在云南松冠层中的垂直空间分布。结果表明:①对于SLA,29年生云南松当年叶和1年叶比138年生云南松分别高27.66%和16.71%;对于LDMC,138年生比29年生分别高14.67%和5.10%;②SLA与LDMC成负相关;③各龄叶SLA随冠层高度的增加均具有对数递减趋势;④约在22m高度以下,LDMC随冠层高度的增加而增加;22m以上,当年叶LDMC呈现下降趋势,但1年叶变化不明显。研究结果指出,光照和水分资源在冠层不同高度的分配,共同导致SLA和LDMC沿冠层垂直方向发生变化。 [ 张林, 罗天祥, 邓坤枚, 李文华 ( 2008). 云南松比叶面积和叶干物质含量随冠层高度的垂直变化规律 北京林业大学学报, 30(1), 40-44.] DOI:10.3321/j.issn:1000-1522.2008.01.007URL [本文引用: 3] 比叶面积(SLA)和叶干物质含量(LDMC)综合反应了植物利用资源的能力,是植物适应环境所体现出的关键叶性状。为深入了解SLA和LDMC沿冠层高度的垂直变化规律,该文对云南省永仁县万马乡2株云南松(树高14.4m,树龄29年;树高30.2m,树龄138年)不同冠层高度叶片SLA和LDMC进行调查研究,分析了不同年龄树木、不同叶龄SLA和LDMC的差异,初步探讨了SLA和LDMC在云南松冠层中的垂直空间分布。结果表明:①对于SLA,29年生云南松当年叶和1年叶比138年生云南松分别高27.66%和16.71%;对于LDMC,138年生比29年生分别高14.67%和5.10%;②SLA与LDMC成负相关;③各龄叶SLA随冠层高度的增加均具有对数递减趋势;④约在22m高度以下,LDMC随冠层高度的增加而增加;22m以上,当年叶LDMC呈现下降趋势,但1年叶变化不明显。研究结果指出,光照和水分资源在冠层不同高度的分配,共同导致SLA和LDMC沿冠层垂直方向发生变化。
ZhaoMF, XiangWH, PengCH, TianDL ( 2009). Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model Forest Ecology and Management, 257, 1520-1531. DOI:10.1016/j.foreco.2008.12.025URL [本文引用: 1] Chinese fir [( Cunninghamia lanceolata (Lamb.) Hook (Taxodiaceae)] plantations are helping to meet China's increasing demands for timber, while, at the same time, sequestering carbon (C) above and belowground. The latter function is important as a means of slowing the rate that CO 2 is increasing in the atmosphere. Available data are limited, however, and even if extensive, would necessitate consideration of future changes in climatic conditions and management practices. To evaluate the contribution of Chinese fir plantations under a range of changing conditions a dynamic model is required. In this paper, we report successful outcome in parameterizing a process-based model (3-PG) and validating its predictions with recent and long-term field measurements acquired from different ages of Chinese fir plantations at the Huitong National Forest Ecosystem Research Station. Once parameterized, the model performed well when simulating leaf area index (LAI), net primary productivity (NPP), biomass of stems ( W S), foliage ( W F) and roots ( W R), litterfall, and shifts in allocation over a period of time. Although the model does not specifically include heterotrophic respiration, we made some attempts to estimate changes in root C storage and decomposition rates in the litterfall pool as well as in the total soil respiration. Total C stored in biomass increased rapidly, peaking at age 21 years in unthinned stands. The predicted averaged above and belowground NNP (13.81 t ha 611 a 611) of the Chinese fir plantations between the modeling period (from 4 to 21-year-old) is much higher than that of Chinese forests (4.8–6.22 t ha 611 a 611), indicating that Chinese fir is a suitable tree species to grow for timber while processing the potential to act as a C sequestration sink. Taking into account that maximum LAI occurs at the age of 15 years, intermediate thinning and nutrient supplements should, according to model predictions, further increase growth and C storage in Chinese fir stands. Predicted future increases (approximately 0–2 °C) in temperature due to global warming may increase plantation growth and reduce the time required to complete a rotation, but further increases (approximately 2–6 °C) may reduce the growth rate and prolong the rotational age.
ZhengYP, ZhuH, XuHN, XuFH, ZhouYQ ( 1991). The characteristics of growing processes of Chinese fir and division of its growth stages Journal of Zhejiang Forestry College, 8, 219-226. URL [本文引用: 1]
ZhouJY, HeJJ, GuoZY, WangBB, ZhangXW, GuoCY ( 2013). A study on specific leaf area and leaf dry matter content of five dominant species in Xiangshan Mountain, Huaibei City, Anhui Province Journal of Huaibei Normal University (Natural Science), 34(3), 51-54. DOI:10.3969/j.issn.2095-0691.2013.03.013URL [本文引用: 1] 植物的功能性状是植物生态学领域近年来研究的热点内容之一.其中,植物叶功能性状与植株生物量、植物对环境资源的获得及其利用的关系最为密切.研究淮北相山5种优势植物叶干物质含量、比叶面积及其相互关系,显示不同物种间的叶干物质含量、比叶面积存在显著差异,两种叶性状间存在明显的负相关关系.依据比叶面积的大小,初步判断5种植物对相山的适应性顺序为柘树扁担木牡荆酸枣构树. [ 周济源, 何俊洁, 郭治远, 王贝贝, 张兴旺, 郭传友 ( 2013). 淮北相山主要优势物种比叶面积与叶干物质含量初步研究 淮北师范大学学报(自然科学版), 34(3), 51-54.] DOI:10.3969/j.issn.2095-0691.2013.03.013URL [本文引用: 1] 植物的功能性状是植物生态学领域近年来研究的热点内容之一.其中,植物叶功能性状与植株生物量、植物对环境资源的获得及其利用的关系最为密切.研究淮北相山5种优势植物叶干物质含量、比叶面积及其相互关系,显示不同物种间的叶干物质含量、比叶面积存在显著差异,两种叶性状间存在明显的负相关关系.依据比叶面积的大小,初步判断5种植物对相山的适应性顺序为柘树扁担木牡荆酸枣构树. Periodicity of leaf growth and leaf dry mass changes in the evergreen and deciduous species of southern Assam, India 1 2013
Correlation between leaf age and other leaf traits in three mediterranean maquis shrub species:Quercus ilex, Phillyrea latifolia and Cistus incanus. 1 2000
Variations in leaf mass per area according to N nutrition, plant age, and leaf position reflect ontogenetic plasticity in winter oilseed rape (Brassica napus l.). 1 2009
... id="C3">随着叶片经济谱(LES) (Wright et al., 2004)研究的不断深入, 叶片功能性状从生产估算到个体的适应和进化的量化研究、从区域经营到全球生物功能地理参考和预测的相互转换成为近年来的研究热点(Lavorel & Garnier, 2002).其中叶干质量比(LDMC)和比叶面积(SLA)作为易于测量的两个叶片性状, 具有一定的样本普适性, 是一对相互关联的重要指标.LDMC是叶干质量(LDM)和叶鲜质量(LFM)的比值, SLA是指单位干质量的鲜叶表面积大小.联合两个指标, 不仅能探索和预测个体行为、群落结构以及生态系统应对气候和环境变化的协调机制和权衡策略(Díaz et al., 1998; Wilson et al., 1999; Díaz & Cabido, 2009), 而且能用于从单株叶片的光合能力指示个体净生产能力、冠层光合能力, 从区域生产经营扩展到全球生产力的变化和验证(Kattge et al., 2009; Wirth & Lichstein, 2009; Zaehle & Friend, 2010; Osnas et al., 2013), 实现从小区域到大尺度、从精算到估算的转换(Kattge et al., 2011). ...
Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models 1 2009
... id="C3">随着叶片经济谱(LES) (Wright et al., 2004)研究的不断深入, 叶片功能性状从生产估算到个体的适应和进化的量化研究、从区域经营到全球生物功能地理参考和预测的相互转换成为近年来的研究热点(Lavorel & Garnier, 2002).其中叶干质量比(LDMC)和比叶面积(SLA)作为易于测量的两个叶片性状, 具有一定的样本普适性, 是一对相互关联的重要指标.LDMC是叶干质量(LDM)和叶鲜质量(LFM)的比值, SLA是指单位干质量的鲜叶表面积大小.联合两个指标, 不仅能探索和预测个体行为、群落结构以及生态系统应对气候和环境变化的协调机制和权衡策略(Díaz et al., 1998; Wilson et al., 1999; Díaz & Cabido, 2009), 而且能用于从单株叶片的光合能力指示个体净生产能力、冠层光合能力, 从区域生产经营扩展到全球生产力的变化和验证(Kattge et al., 2009; Wirth & Lichstein, 2009; Zaehle & Friend, 2010; Osnas et al., 2013), 实现从小区域到大尺度、从精算到估算的转换(Kattge et al., 2011). ...
Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan. 1 2002
... id="C30">杉木叶片生活周期较长, 其LDMC和SLA在不同叶龄下的分配策略能表现叶片生活史弹性适应策略(Reich et al., 1991b; Frak et al., 2001; 霍宏和王传宽, 2007; 黄海侠等, 2014).老叶往往表现出最小的SLA ((64.01 ± 11.73) cm2·g-1)和最大的LDMC (0.45 ± 0.06), 以及最小的变异程度(0.18和0.13), 这是由于随着叶龄增加, 叶片内次生代谢物积累, 叶片为了防虫食和提高外界干扰的防御能力, 会减小叶面积、增加保卫构造组织, 叶片变厚、含水率降低, 以减少失水(Witkowski & Lamont, 1991; Reich et al., 1998; Kayama et al., 2002), 随着光合能力的减弱, 生长发育受到限制, 老叶的形态可塑性最小(Field & Mooney, 1983; Ono et al., 2001).而新生叶、一年生叶和二年生叶都呈现出高SLA低LDMC, 以及较大的形态可塑性(Frak et al., 2001; Wyka et al., 2008). ...
Applications of physiological ecology to forest management 1 1997
... id="C4">LDMC指示植物含水状况、养分和物质的储存能力和分配策略, 与水分利用效率密切相关(Cornelissen et al., 2003).例如在干旱和贫瘠的环境下, LDMC可能更适用于单独预测个体植物行为和生活史对策(Garnier et al., 2001a, 2001b; Hodgson et al., 2011; Smart et al., 2017), 被认为是鉴别和定位植物资源分配单元的最佳变量(Wilson et al., 1999).SLA预示着叶片光截取和碳获取能力(Westoby, 1998), 与光合、呼吸、蒸腾等生理过程密切相关.SLA作为主导变量(Westoby et al., 2002), 能灵敏地反映植物在不同环境下的光合过程和适应特征(Garnier et al., 2001b), 预测冠层光环境变化下的植物潜在光合固碳能力(Ellsworth & Reich, 1993).SLA常被用来估算作物产量、森林生物量(Bertin, 1998)和生态系统净生产力(Smart et al., 2017), 促进农林生产经营(Landsberg & Gower, 1997; Bertin, 1998).同时, LDMC和SLA是一对相互联系的重要指标, 它们能共同指示植物在不同生长阶段的生长策略.例如, 在叶片生长早期, 叶片需要获得较强的光资源获取能力、充足的含水率支持较强的代谢活动, 会表现出较高的SLA和较低的LDMC (Field & Mooney, 1983; Sims & Pearcy, 1992). ...
Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the holy grail 1 2002
... id="C3">随着叶片经济谱(LES) (Wright et al., 2004)研究的不断深入, 叶片功能性状从生产估算到个体的适应和进化的量化研究、从区域经营到全球生物功能地理参考和预测的相互转换成为近年来的研究热点(Lavorel & Garnier, 2002).其中叶干质量比(LDMC)和比叶面积(SLA)作为易于测量的两个叶片性状, 具有一定的样本普适性, 是一对相互关联的重要指标.LDMC是叶干质量(LDM)和叶鲜质量(LFM)的比值, SLA是指单位干质量的鲜叶表面积大小.联合两个指标, 不仅能探索和预测个体行为、群落结构以及生态系统应对气候和环境变化的协调机制和权衡策略(Díaz et al., 1998; Wilson et al., 1999; Díaz & Cabido, 2009), 而且能用于从单株叶片的光合能力指示个体净生产能力、冠层光合能力, 从区域生产经营扩展到全球生产力的变化和验证(Kattge et al., 2009; Wirth & Lichstein, 2009; Zaehle & Friend, 2010; Osnas et al., 2013), 实现从小区域到大尺度、从精算到估算的转换(Kattge et al., 2011). ...
Carbon and nitrogen cycle dynamics in the O-CN land surface model: 1. Model description, site-scale evaluation, and sensitivity to parameter estimates 1 2010
... id="C3">随着叶片经济谱(LES) (Wright et al., 2004)研究的不断深入, 叶片功能性状从生产估算到个体的适应和进化的量化研究、从区域经营到全球生物功能地理参考和预测的相互转换成为近年来的研究热点(Lavorel & Garnier, 2002).其中叶干质量比(LDMC)和比叶面积(SLA)作为易于测量的两个叶片性状, 具有一定的样本普适性, 是一对相互关联的重要指标.LDMC是叶干质量(LDM)和叶鲜质量(LFM)的比值, SLA是指单位干质量的鲜叶表面积大小.联合两个指标, 不仅能探索和预测个体行为、群落结构以及生态系统应对气候和环境变化的协调机制和权衡策略(Díaz et al., 1998; Wilson et al., 1999; Díaz & Cabido, 2009), 而且能用于从单株叶片的光合能力指示个体净生产能力、冠层光合能力, 从区域生产经营扩展到全球生产力的变化和验证(Kattge et al., 2009; Wirth & Lichstein, 2009; Zaehle & Friend, 2010; Osnas et al., 2013), 实现从小区域到大尺度、从精算到估算的转换(Kattge et al., 2011). ...
云南松比叶面积和叶干物质含量随冠层高度的垂直变化规律 3 2008
... id="C5">然而, SLA不仅易受叶片自身大小、厚度、形状的影响(Witkowski & Lamont, 1991), 而且受限于叶片干质量和叶面积两个指标.目前对植物SLA的测定局限性主要表现在两方面: 一是叶面积测量技术瓶颈, 二是样本的局限性.例如, 叶面积的测定经常受限于测量技术上的困难(Cornelissen et al., 2003), 长久以来都不是一项简单的工作, 总是伴随着人力、物力和时间的高花费(Wilson et al., 1999; Roche et al., 2004), 且并不适用于所有样本, 一些针叶甚至无叶植物的叶面积测定至今很困难(Wilson et al., 1999).也有大量研究探讨SLA和LDMC的关联(李玉霖等, 2005; 张林等, 2008; 任昱等, 2015), 其理解主要针对SLA和LDMC对环境因子的响应, 常探究指标与环境因子间的关联而不是关系, 但用特定 环境下的性状相关性来指导现实问题, 往往收效 甚微. ...
... id="C27">SLA和LDMC暗示着植物快速扩展光截取能力和高效存储物质用于叶结构建成这两种资源分配策略的转变(Reich et al., 1992; Grime et al., 1997; Shipley et al., 2006; Hodgson et al., 2011).叶片SLA和LDMC不仅会对冠层深度(Ellsworth & Reich, 1993; 张林等, 2008)、坡向(党晶晶等, 2015; 欧晓岚和刘艳红, 2017)等小生境的环境梯度变化做出响应, 还会因自身的生长发育(如林龄和叶龄)(Wyka et al., 2008; Karavin, 2013; 黄海侠等, 2014)而不断调整生活史策略.本研究中杉木的SLA范围在96.08-110.22 cm2·g-1, 已有研究中杉木SLA在74-104 cm2·g-1(李轩然等, 2007; 李凯和项文化, 2011), 本研究的抽样样本既包含不同产区杉木, 又包含不同林龄级和叶龄级杉木, 以及不同小生境梯度(冠层、坡向、坡位等), 而已有研究样本受限于特定的林龄级等(李轩然等, 2007; 李凯和项文化, 2011).杉木的SLA较小(Garnier et al., 1997), 已有研究认为低SLA的物种更能适应干旱和贫瘠的环境(Wilson et al., 1999; Westoby et al., 2002), 这预示着杉木作为亚热带最重要的用材树种, 能很好地面对和适应未来可能出现的降水减少带来干旱胁迫增加的环境变化. ...
 ,*中南林业科技大学生命科学与技术学院, 长沙 410004; 湖南会同杉木林国家重点野外科学观测研究站, 湖南会同 418307; 南方林业生态应用技术国家工程实验室, 长沙 410004
,*中南林业科技大学生命科学与技术学院, 长沙 410004; 湖南会同杉木林国家重点野外科学观测研究站, 湖南会同 418307; 南方林业生态应用技术国家工程实验室, 长沙 410004
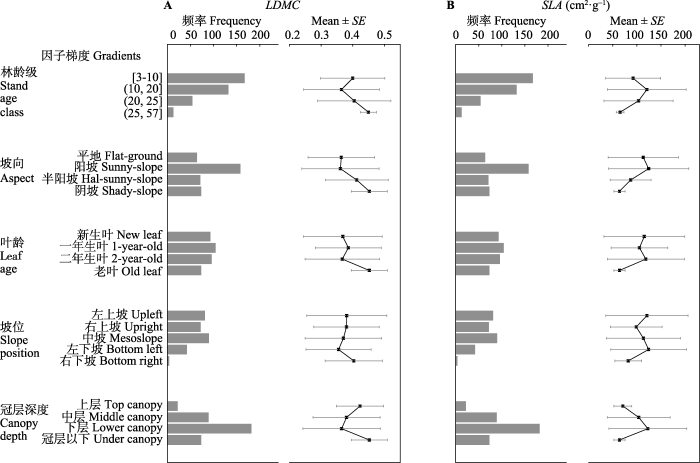 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT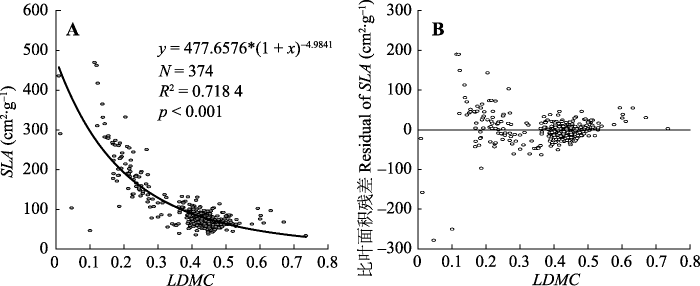 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT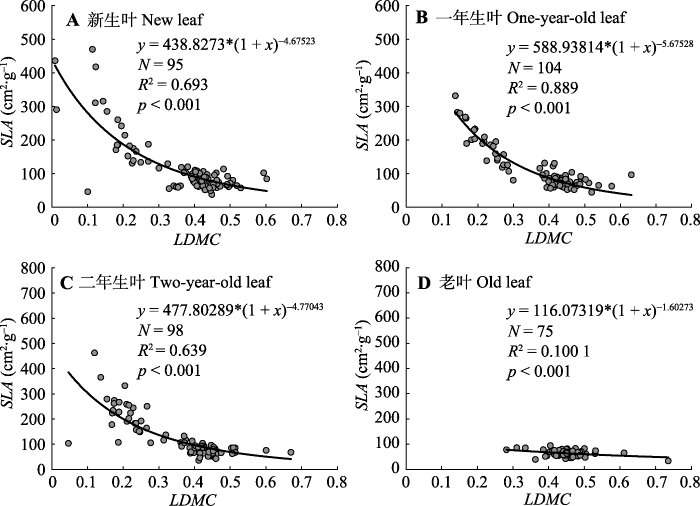 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT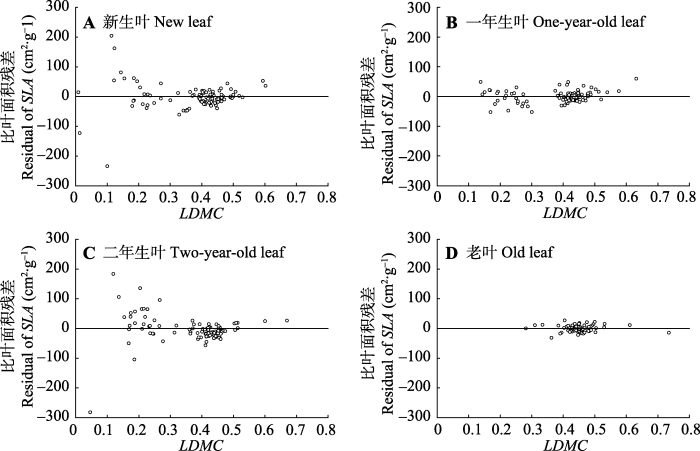 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}