 ,*四川农业大学生态林业研究所, 长江上游林业生态工程四川省重点实验室, 高山森林生态系统定位研究站, 水土保持与荒漠化防治省级重点实验室, 成都 611130; 长江上游生态安全协同创新中心, 成都 611130
,*四川农业大学生态林业研究所, 长江上游林业生态工程四川省重点实验室, 高山森林生态系统定位研究站, 水土保持与荒漠化防治省级重点实验室, 成都 611130; 长江上游生态安全协同创新中心, 成都 611130Effects of forest gap size on initial decomposition of twig litter in the subalpine forest of western Sichuan, China
GUO Cai-Hong, YANG Wan-Qin, WU Fu-Zhong, XU Zhen-Feng, YUE Kai, NI Xiang-Yin, YUAN Ji, YANG Fan, TAN Bo,*Institute of Ecology & Forestry, Sichuan Agricultural University, Forestry Ecological Engineering in Upper Reaches of Yangtze River Key Laboratory of Sichuan Province, Alpine Forest Ecosystem Research Station, Soil and Water Conservation and Desertification Control Key Laboratory of Sichuan Province, Chengdu 611130, China; and Collaborative Innovation Center of Ecological Security in the Upper Reaches of Yangtze River, Chengdu 611130, China通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (2811KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
郭彩虹, 杨万勤, 吴福忠, 徐振锋, 岳楷, 倪祥银, 袁吉, 杨帆, 谭波. 川西亚高山森林林窗对凋落枝早期分解的影响. 植物生态学报[J], 2018, 42(1): 28-37 doi:10.17521/cjpe.2017.0186
GUO Cai-Hong.
植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(Aerts, 1997; Berg & McClaugherty, 2014)。已有的研究表明, 凋落叶约占森林生态系统凋落物年归还总量的70%, 凋落枝约占26% (Berg & McClaugherty, 2014)。现有的研究大多关注凋落叶分解过程, 分解模型也大多基于凋落叶开展, 对凋落枝的认识较少(Parton et al., 2007; Adair et al., 2008)。Cha等(2017)通过40个月的分解实验发现, 6种温带落叶树种的凋落物枝条质量均损失了75%以上, 且夏季凋落叶和凋落枝具有最大的分解速率(Gosz et al., 1973)。Shorohova和Kapitsa (2016)对欧洲北方针叶林粗木质残体的研究表明, 凋落枝比凋落叶具有更长的分解周期, 分解速率(k)与树种密切相关。因此, 在森林生态系统的凋落物养分释放方面, 凋落枝会比凋落叶缓慢。
林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(McCarthy, 2001; 吴庆贵等, 2013), 可能通过两方面的机制影响高山森林凋落物的分解过程: (1)形成不同于林下的微环境条件及土壤生物群落结构(McCarthy, 2001; 管云云等, 2016); (2)直接增加了降水对地表凋落物的淋洗作用, 加速了凋落物的质量损失和养分释放(Zhang & Zak, 1995; Prescott et al., 2003)。例如, Arunachalam和Arunachalam (2000)在亚热带湿润森林的研究指出, 上层林冠疏开增加了土壤温度和湿度, 林下微环境的改变进一步提高了土壤微生物生物量碳含量和雨季的真菌群落数量; He等(2015, 2016)在高山森林的研究发现, 林窗对冬季雪被环境的改变增加了凋落物木质素和纤维素的降解以及微生物生物量。同时, 林窗面积大小驱动的水热变化会对高山森林凋落物分解施加强烈影响(Baptist et al., 2010; Zhu et al., 2015; He et al., 2016)。相比于郁闭林下, 在冬季期间, 林窗内积雪的隔热保温作用能使土壤微生物维持相对较高的活性(Baptist et al., 2010; Christenson et al., 2010), 林窗可能促进冬季凋落物分解, 且不同大小的林窗对雪被厚度和冻融循环格局的改变也可能不同程度地作用于凋落物分解(Aerts, 2006; Wu et al., 2014); 在生长季节, 林窗内相对更高的光照强度和光辐射效率可导致地表温度升高, 水分蒸发增加以及降雨淋洗增强(Denslow et al., 1998; Varhola et al., 2010; Devi et al., 2012), 林窗可能抑制或促进生长季节凋落物的分解。然而, 迄今为止有关林窗及其面积变化对凋落物分解的研究结论仍不甚清晰。例如, Prescott等(2000)研究发现, 随着林窗面积增大, 淋溶和光降解作用增大, 促进了凋落物的分解; 相反, Zhang和Zak (1995)研究认为, 具有更高温湿度条件的林下(或小林窗)的凋落物损失率比大林窗更高; Denslow等(1998)研究表明, 林窗大小对凋落物分解无显著影响。值得注意的是, 这些研究结论大多基于凋落叶分解实验得出, 有关林窗大小对凋落枝分解的影响尚不清楚, 这限制了对高山森林物质循环过程的全面认识。
位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(刘庆和吴彦, 2002; 吴庆贵等, 2016)。该区森林冬季具有明显的季节性雪被和土壤冻融过程(Wu et al., 2010), 森林群落的自然更新以林窗更新为主(刘庆和吴彦, 2002; 吴庆贵等, 2013), 是研究亚高山林窗对凋落物分解影响的天然实验平台。前期研究表明, 林窗位置和大小对川西亚高山森林冬季和生长季节凋落叶质量损失、元素释放以及腐殖化过程产生显著作用(Wu et al., 2014; He et al., 2015, 2016)。同时, 亚高山森林每年有371.28 kg·hm-2凋落物以枝条的形式归还到地表(Fu et al., 2017)。凋落枝普遍具有比凋落叶更高的碳和难降解组分含量(如木质素), 其分解特征及其对林窗面积变化的响应可能不同于凋落叶。然而, 关于该区森林凋落枝分解特征及其对林窗变化响应的研究未见报道。因此, 我们以川西亚高山/高山地区普遍分布的岷江冷杉(Abies faxoniana)凋落枝为对象, 研究林窗大小对岷江冷杉凋落枝质量损失的影响。我们假设: (1)冬季是亚高山森林凋落枝分解的重要时期, 冬季凋落枝质量损失随林窗面积增大而增加; (2)林窗中的亚高山森林凋落枝质量损失过程随林窗面积和分解持续时间具有变异性。研究结果可为全面地认识川西亚高山森林物质循环过程提供参考。
1 材料和方法
1.1 研究区域与样地概况
研究区域位于四川省阿坝藏族羌族自治州理县的四川农业大学高山森林生态系统定位研究站(102.88°-102.95° E, 31.23°-31.32° N, 海拔2 458-4 619 m)。研究站坐落于米亚罗自然保护区, 地处青藏高原东缘与四川盆地的过渡带, 区域内年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1 ℃, 年降水量约850 mm, 季节性冻融期长达5-6个月(吴庆贵等, 2013)。森林植被随海拔由低到高依次为针阔混交林、高寒针叶林、高山灌丛和草甸, 乔木树种主要有岷江冷杉、红桦(Betula albo-sinensis)、方枝柏(Sabina saltuaria)、粗枝云杉(Picea asperata)和四川红杉(Larix mastersiana)等, 林下灌木以康定柳(Salix paraplesia)、箭竹(Fargesia spathacea)、三颗针(Berberis julianae)、红毛花楸(Sorbus rufopilosa)、沙棘(Hippophae rhamnoides)等为主, 草本以蟹甲草(Cacalia spp.)、冷蕨(Cystopteris montana)、薹草属(Carex)和莎草属(Cyperus)等为主(吴庆贵等, 2016)。土壤为发育在灰岩、页岩、板岩的棕壤和暗棕壤, 土壤有机层厚度约为12.5 cm, pH值约为6.5。1.2 实验设计和样品处理
在高山森林生态系统定位研究站建立的1 hm2岷江冷杉原始林样地(坡向NE 38°, 坡度 24°)进行分解实验, 海拔3 598 m。林窗设置均指扩展林窗, 其边缘以边界木树干基部为准(Schliemann & Bockheim, 2011), 计算四周边界木围成的近似椭圆的面积作为扩展林窗的面积。前期调查发现, 该区域内最大的林窗面积约为280 m2 (吴庆贵等, 2013), 基于调查结果, 本实验选取的3种林窗面积大小分别为255-290 m2 (FG1)、153-176 m2 (FG2)和38-46 m2 (FG3)。同时, 在岷江冷杉郁闭林下(CC)设置3个10 m × 10 m的样方作为对照。林窗调查于2012年生长季节进行, 林窗的基本特征见表1。Table 1
表1
表1川西亚高山森林林窗基本性质
Table 1
| 林窗类型 Gap type | 林窗面积(m2) Area of gap | 林窗形成木 Species of gap maker | 林窗边界木 Species of gap border | 林窗形成方式 Causes of gap |
|---|---|---|---|---|
| FG1 | 255-290 | 岷江冷杉, 高度以20-30 m为主, 胸径以40-70 cm居多 Abies faxoniana with height and diameter at breast height ranging between 20 and 30 m and between 40 and 70 cm, respectively | 岷江冷杉、红桦, 高度以20-30 m为主, 胸径以40-60 cm居多 Abies faxoniana and Betula albo-sinensis with height and diameter at breast height ranging between 20 and 30 m and between 40 and 60 cm, respectively | 折干, 枯立 Breakage at trunk, standing death |
| FG2 | 153-176 | |||
| FG3 | 38-46 |
新窗口打开|下载CSV
2012年5-9月, 采集岷江冷杉地表新鲜凋落枝(直径为0.5-1.5 cm), 45 ℃烘干, 统一剪切为5 cm长度, 混匀后分装于大小为20 cm × 20 cm、孔径为1.0 mm的尼龙网凋落物袋中, 每袋10 g备用。凋落枝初始质量见表2。
Table 2
表2
表2岷江冷杉凋落枝初始质量特征(平均值±标准误差, n = 5)
Table 2
| 有机碳(g·kg-1) Organic carbon (C) | 总氮(g·kg-1) Total nitrogen (N) | 总磷(g·kg-1) Total phosphorus (P) | 木质素(%) Lignin | 纤维素(%) Cellulose | 碳:氮 C:N | 碳:磷 C:P | 氮:磷 N:P | 木质素:氮 Lignin:N |
|---|---|---|---|---|---|---|---|---|
| 441.8 ± 10.7 | 8.11 ± 0.20 | 0.7 ± 0.1 | 32.4 ± 1.7 | 23.8 ± 1.2 | 54.4 ± 0.9 | 657.4 ± 21.4 | 12.1 ± 0.4 | 41.7 ± 0.8 |
新窗口打开|下载CSV
2012年11月将制备的分解袋埋设于样地, 分解实验为期4年, 共埋设960个分解袋, 每次取样(10) ×林窗(4) ×重复(3) × 8次。于分解袋埋设后的6个月间隔周期回收分解袋, 每一间隔代表冬季或者生长季节分解变化特征, 具体时间为2013年4月24日(第1年冬季, 1W)、2013年11月13日(第1年生长季, 1GS)、2014年4月24日(第2年冬季, 2W)、2014年10月29日(第2年生长季, 2GS)、2015年4月23日(第3年冬季, 3W)、2015年11月2日(第3年生长季, 3GS)、2016年4月25日(第4年冬季, 4W)、2016年10月30日(第4年生长季, 4GS)。采样时, 随机从每个林窗内采集岷江冷杉枝条分解袋各10袋, 小心去除泥土杂物及新生根系, 置于有冰袋的保鲜盒中24 h内运回实验室, 放入4 ℃冰箱保存备用。采用纽扣式温度记录器DS1923-F5# (Maxim/Dallas Semiconductor, Sunnyvale, USA)连续监测各林窗下凋落袋下方地表温度, 设定每2 h记录一次温度数据, 每3个月下载一次数据。每年冬季采用钢尺测量林窗及林下雪被厚度, 具体时间为: 2012年11和12月, 2013、2014和2015年3、4、11和12月, 2016年3、4和11月。
将采集的样品分为两部分。一部分(4袋)用于枝条持水能力测定, 具体方法为: 将烘干至恒质量的凋落物称量, 然后装入凋落袋中, 浸入清水中24 h, 水面高度以浸过凋落袋, 凋落袋不露出水面为度, 将内装凋落物的凋落袋从清水中取出并悬挂在空中静置约5 min, 当无水滴滴下时立刻称量, 最后将袋中的凋落物烘干(65 ℃)称量, 计算最大持水率(陈百灵等, 2015)。将24 h的持水率作为其最大持水率; 剩余部分(6袋) 65 ℃烘干至恒质量后, 用于计算质量损失和测定元素含量。
1.3 数据处理与统计方法
在温度监测的任何时段, 将凋落袋下方地表温度低于0 ℃持续至少3 h, 随后回升到0 ℃至少持续3 h (或高于0 ℃持续至少3 h, 随后跌至0 ℃以下持续至少3 h)的温度动态过程记作一次冻融循环(Konestabo et al., 2007)。最大持水率=[(24 h湿质量-干质量)/干质量]× 100%
凋落物分解指数模型为: y = ae-kt
式中y为凋落物的月残留率(%), a为拟合参数, e为自然对数底, t为分解时间(月), k为凋落物分解系数(g·g-1) (Olson, 1963)。
分解时间: 半分解时间(T50%)、95%分解时间(T95%)计算式:
50%分解时间= -ln(1-0.50)/k (吴庆贵等, 2016)
95%分解时间= -ln(1-0.95)/k
质量损失率和质量损失速率(Baptist et al., 2010; 吴庆贵等, 2016)按如下公式计算:
凋落物质量损失率: Lt (%) = (M0- Mt)/M0 × 100%
每30天质量损失速率: Vt (%) = Lt (%)/DΔt × 30
凋落物质量损失4年时间各阶段所占比例(何伟等, 2013): P (%) = (Mt-1- Mt)/(M0- MT) × 100%
式中: Mt为t时刻凋落叶干质量(g), M0为初始干质量(g), (Mt-1- Mt)为相邻采样时间凋落叶残留量差(t = 1, 2, 3…8), DΔt为相邻采样时间间隔天数, MT为最后一次采样凋落物袋内凋落物残留量。
重复测量方差分析和最小显著差异法(LSD)比较检验不同面积林窗和林下环境特征(雪被、温度和冻融循环)、持水能力、质量损失速率的差异。比较不同面积林窗和林下质量残留率的差异, 采用指数回归分析拟合凋落枝质量损失与分解时间的关系。以上分析通过SPSS 22.0 (IBM SPSS Statistics, Chicago, USA)软件包完成, 指数拟合在Origin Pro9.0 (OriginLab, Northampton, USA)中进行。
2 结果
2.1 环境特征
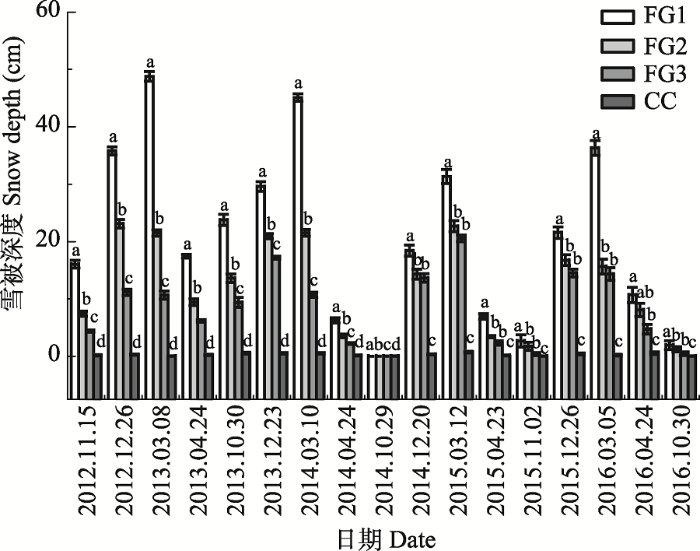
从图1可见, 亚高山森林不同面积林窗和林下具有明显的雪被过程。整个研究期间, 雪被深度呈现随林窗面积减小逐渐降低的变化趋势, FG1、FG2、FG3和CC 4年雪被厚度分别为36.3-48.7、15.7-22.7、10.7-20.5和0-0.6 cm, 且林窗显著影响(F = 14.41, p < 0.001)岷江冷杉枝条分解过程中的积雪深度。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1川西亚高山森林不同面积林窗和郁闭林下的雪被厚度(平均值±标准误差)。不同小写字母代表林窗间差异显著。FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。
Fig. 1Snow depths associated with the forest gaps and the closed canopy in the subalpine forest of Western Sichuan (mean ± SE). Different lowercase letters indicate significant differences among the three forest gaps and the closed canopy. FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy.
整个研究期间, 岷江冷杉枝条分解过程的地表平均温度和冻融循环频次随林窗面积大小和季节变化表现出明显的动态变化特征(图2)。FG1在冬季和生长季节均具有更高的地表平均温度, 冬季和生长季节地表平均温度分别为-0.6-0.9 ℃和8.8-9.8 ℃, 且显著高于FG3和林下(F = 40.78, p < 0.001), 而FG3与CC凋落枝的地表平均温度差异不显著(F = 4.232, p = 0.052), 冬季和生长季节地表平均温度为-0.6-0.9 ℃和7.0-7.8 ℃ (图2A)。冬季的冻融循环频次高于生长季节, 林窗并未显著影响(F = 2.36, p > 0.102)枝条分解过程中的冻融循环频次, 各林窗第1和第3年冬季冻融循环频次变化规律表现为FG2 > FG3 > FG1, 第2和第4年冬季融循环频次变化规律表现为随着林窗面积的增加而减少(图2B)。
图2
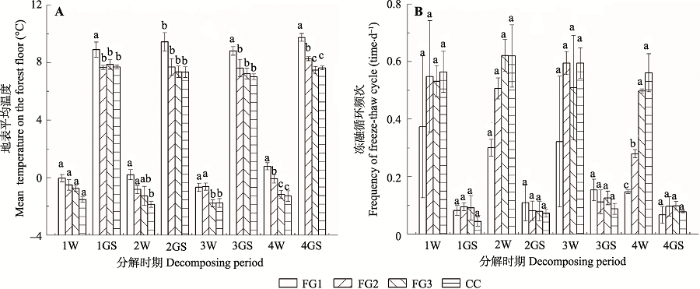 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2川西亚高山森林不同面积林窗和郁闭林下地表平均温度和冻融循环频次(平均值±标准误差)。不同小写字母代表林窗间差异显著。FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。1W, 第1年冬季; 1GS, 第1年生长季节; 2W, 第2年冬季; 2GS, 第2年生长季节; 3W, 第3年冬季; 3GS, 第3年生长季节; 4W, 第4年冬季; 4GS, 第4年生长季节。
Fig. 2Mean surface temperature and frequency of freeze-thaw cycles in the litterbag associated with the forest gaps and the closed canopy in the subalpine forest of Western Sichuan (mean ± SE). Different lowercase letters indicate significant differences among the three forest gaps and the closed canopy. FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy. 1W, the first winter; 1GS, the first growing season; 2W, the second winter; 2GS, the second growing season; 3W, the third winter; 3GS, the third growing season; 4W, the fourth winter; 4GS, the fourth growing season.
2.2 持水能力
由图3可见, 林窗和林下岷江冷杉枝条的持水能力在整个研究期间表现出先降低后升高, 且冬季和生长季节各林窗中枝条的持水能力变化显著(F = 3.805, p = 0.007)。在第1年冬季, FG2枝条的持水能力最大(46.8%), CC最小(36.2%), 而FG1枝条的持水能力在第1年生长季节最高(42.6%), FG3最低(25.3%)。林窗和林下枝条的持水能力随分解的进行逐渐增高, 枝条的持水能力到分解的第4年均达到了100%以上, 且林下枝条的持水能力显著高于林窗。图3
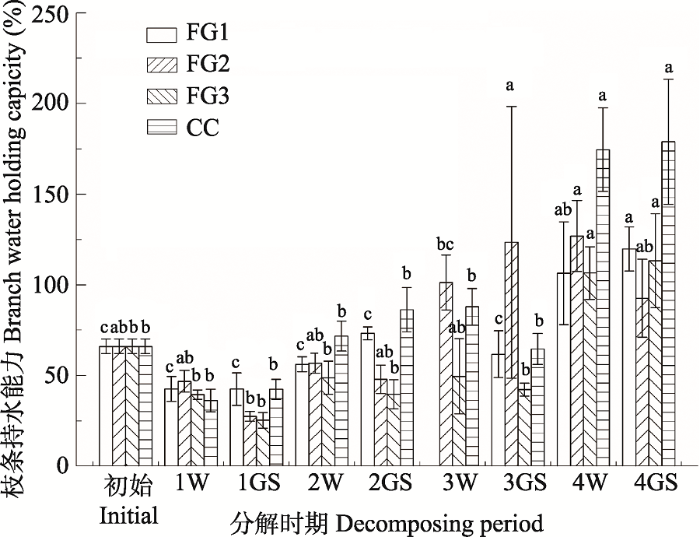 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3川西亚高山森林不同面积林窗和郁闭林下凋落枝持水能力(平均值±标准误差)。不同小写字母代表林窗间差异显著。FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。1W, 第1年冬季; 1GS, 第1年生长季节; 2W, 第2年冬季; 2GS, 第2年生长季节; 3W, 第3年冬季; 3GS, 第3年生长季节; 4W, 第4年冬季; 4GS, 第4年生长季节。
Fig. 3Water holding capacity of twig litter associated with the forest gaps and the closed canopy in the subalpine forest of Western Sichuan (mean ± SE). Different lowercase letters indicate significant differences among the three forest gaps and the closed canopy. FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy. 1W, the first winter; 1GS, the first growing season; 2W, the second winter; 2GS, the second growing season; 3W, the third winter; 3GS, the third growing season; 4W, the fourth winter; 4GS, the fourth growing season.
2.3 质量残留率
经过4年的分解, FG1、FG2、FG3和CC的岷江冷杉枝条质量残留率分别为59.9%、59.5%、62.1%、55.3% (图4)。冬季总累积质量损失量(17.1%-28.2%)高于生长季节总累积质量损失量(16.6%-20.1%), 冬季总累积质量损失量以林下最高(28.2%)、FG3最低(17.1%), 生长季节总累积质量损失量以FG2最大(20.1%)、FG1最小(16.6%)。凋落物分解指数模型拟合显示, 分解系数k值表现为CC (0.135) > FG2 (0.131) > FG1 (0.127) > FG3 (0.120), FG1、FG2、FG3和CC岷江冷杉枝条半分解时间分别需要5.5、5.3、5.8和5.1 a, 95%分解时间分别为23.6、22.7、25.0和22.2 a (表3)。与林下相比, 林窗并未显著增加(F = 0.601, p = 0.615)岷江冷杉枝条质量损失量, 而FG2在3种面积的林窗中具有更大的质量损失量。图4
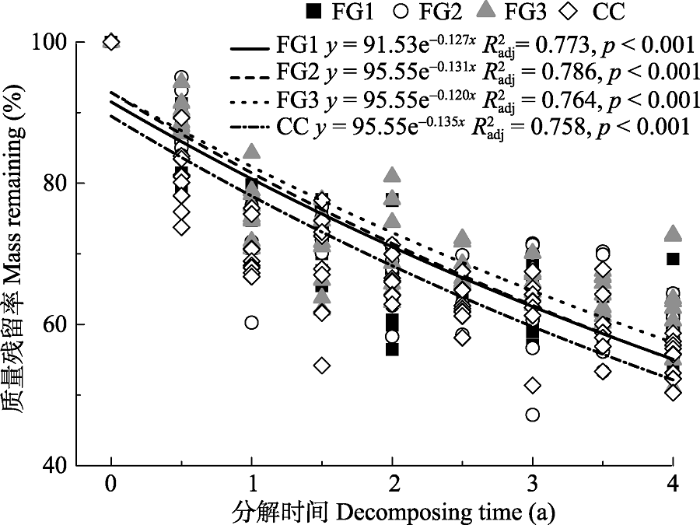 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4川西亚高山森林不同面积林窗和郁闭林下凋落枝质量残留率。不同小写字母代表林窗间差异显著。FG1, 255- 290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。R2adj, 调整决定系数。
Fig. 4Mass remaining of twig litter associated with the forest gaps and the closed canopy in the subalpine forest of Western Sichuan. Different lowercase letters indicate significant differences among the three forest gaps and the closed canopy. FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy. R2adj, adjusted coefficient of determination.
Table 3
表3
表3不同面积林窗和郁闭林下凋落枝分解特征分解系数、相关系数、半分解和95%分解时间
Table 3
| 林窗类型 Gap type | 回归方程 Regression equation | 分解系数k Decomposition constant k | 调整决定系数 Adjusted coefficient of determination | 半分解时间 Time of half decomposition (a) | 95%分解时间 Time of 95% decomposition (a) | 显著性 Significance |
|---|---|---|---|---|---|---|
| FG1 | y = 91.53e-0.127t | 0.127 | 0.773 | 5.458 | 23.588 | p < 0.001 |
| FG2 | y = 95.55e-0.131t | 0.131 | 0.786 | 5.291 | 22.868 | p < 0.001 |
| FG3 | y = 95.55e-0.120t | 0.120 | 0.764 | 5.776 | 24.964 | p < 0.001 |
| CC | y = 95.55e-0.135t | 0.135 | 0.758 | 5.134 | 22.191 | p < 0.001 |
新窗口打开|下载CSV
2.4 质量损失速率
整个研究期间, 林窗和林下岷江冷杉枝条质量损失速率随分解过程总体呈现降低的趋势(图5), FG1质量损失速率以第1年冬季最高(2.5%), 第3年生长季节最低(-0.2%), FG2和FG3则以第1年生长季节最高(分别为2.4%和2.2%), 第4年生长季节最低(均为0.3%)。与林下相比, 林窗显著增加了第1年和第2年生长季节的质量损失速率, 降低了第1年和第4年冬季的枝条质量损失速率。林窗在不同的分解时期对质量损失速率的影响不同, 质量损失速率在第1年和第3年冬季随林窗面积增大而增大, 在第3年生长季节随林窗面积增大而降低, 其余时期无明显规律。图5
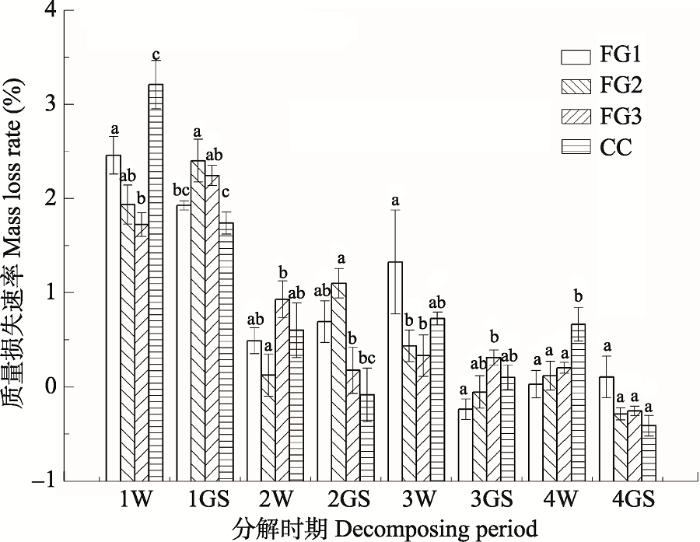 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5川西亚高山森林不同面积林窗和郁闭林下凋落枝每30天的质量损失速率(平均值±标准误差)。FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。1W, 第1年冬季; 1GS, 第1年生长季节; 2W, 第2年冬季; 2GS, 第2年生长季节; 3W, 第3年冬季; 3GS, 第3年生长季节; 4W, 第4年冬季; 4GS, 第4年生长季节。
Fig. 5Mass loss rate per 30 days of twig litter associated with the forest gaps and the closed canopy in the subalpine forest of Western Sichuan (mean ± SE). FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy. 1W, the first winter; 1GS, the first growing season; 2W, the second winter; 2GS, the second growing season; 3W, the third winter; 3GS, the third growing season; 4W, the fourth winter; 4GS, the fourth growing season.
2.5 各时期质量损失比例
由图6可知, 整个研究期间, 林窗和林下岷江冷杉枝条质量损失主要发生在第1年, FG1、FG2、FG3和CC质量损失比例分别占整个分解时期的67.6%、64.3%、62.9%和59.7%。其中, FG1、FG2、FG3和CC第1年冬季质量损失所占比例分别为37.9%、35.5%、35.7%和30.2%, 第1年生长季节质量损失所占比例分别为29.7%、28.8%、27.2%和29.5%, 且第1年冬季和生长季节质量损失所占比例随林窗面积增加而增加。图6
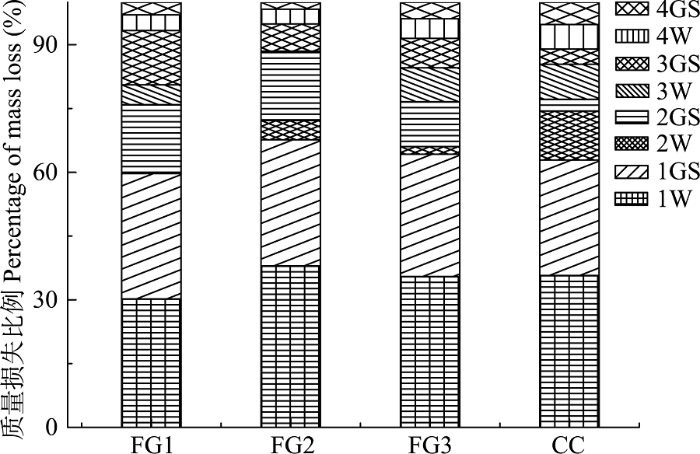 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6川西亚高山森林不同面积林窗和郁闭林下各关键时期凋落枝质量损失比例。FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, 郁闭林下。1W, 第1年冬季; 1GS, 第1年生长季节; 2W, 第2年冬季; 2GS, 第2年生长季节; 3W, 第3年冬季; 3GS, 第3年生长季节; 4W, 第4年冬季; 4GS, 第4年生长季节。
Fig. 6Percentage of mass loss at different sampling stages associated with different forest gaps and the closed canopy in the subalpine forest of Western Sichuan. FG1, 255-290 m2; FG2, 153-176 m2; FG3, 38-46 m2; CC, closed canopy. 1W, the first winter; 1GS, the first growing season; 2W, the second winter; 2GS, the second growing season; 3W, the third winter; 3GS, the third growing season; 4W, the fourth winter; 4GS, the fourth growing season.
3 讨论
林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(臧润国等, 1999), 而且能改变影响凋落物分解的微环境条件, 调控森林生态系统物质循环过程(Sariyildiz, 2008; 管云云等, 2016)。本研究中, 雪被深度总体呈现随林窗面积减小至林下逐渐降低的变化趋势, 冻融循环频次则随林窗面积增大而减少。然而与林下相比, 255-290 m2林窗(FG1)在整个分解期间都具有更高的分解温度和更少的冻融循环频次(图2)。这与随林窗面积增大、土壤平均温度降低的结果(Zhang & Zak, 1995; Sariyildiz, 2008)相反。FG1冬季深厚雪被的绝热保温作用促使凋落物分解温度维持在较高水平, 而由于研究样地海拔(3 598 m)较高, 长期面临低温环境, 林窗并未明显提升生长季节内凋落物的分解温度。由此可见, 林窗面积变化明显改变了冬季凋落枝的分解环境。目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(Shorohova & Kapitsa, 2016)。本研究中, 岷江冷杉凋落枝分解4年后的质量损失率为37.9%-44.7%, 其中, 第1年的质量损失率为23.8%-29.7%。枝条4年的质量损失量明显小于加拿大北方森林凋落叶(Prescott et al., 2000)和温带落叶森林粗枝(Cha et al., 2017), 但枝条第1年质量损失量与同区域岷江冷杉凋落叶的质量损失量相当(何伟等, 2013; 吴庆贵等, 2016), 且显著高于温带落叶森林(Cha et al., 2017)和亚热带森林枝条(Zhang & Zak, 1995)。造成这种变化的原因是: (1)新近凋落物常常具有较高的可溶性有机物质含量(可溶性碳和氮等), 雪被融化的淋洗作用和冻融循环的物理破碎能显著地促进寒冷季节凋落枝条相关组分的快速降解(Baptist et al., 2010; Wu et al., 2010); (2)雪被的绝热保温作用维持耐寒生物种群活动和生物活性同样能提高冬季凋落叶和枝条降解率(Baptist et al., 2010; Christenson et al., 2010); (3)冬季冻融作用机械破碎促使凋落枝条在水热条件和生物活性更为适宜的生长季节更容易被淋洗和降解(Aerts, 2006); (4)较低的C:N和木质素:N更有利于凋落枝分解初期质量的快速损失(Gavazov, 2010; Shorohova & Kapitsa, 2016; Cha et al., 2017)。当然, 研究持续时间和林地环境的差异也可能造成质量损失速率的变异。此外, 随着分解持续进行, 林窗和林下的质量损失率在第1年后开始显著降低, 这是由于淋溶作用减弱以及木质素、半纤维素等难降解物质的相对含量上升(Aerts, 1997; He et al., 2015, 2016)。由此可见, 亚高山森林冬季和生长季节明显的季节变化对亚高山森林物质循环有重要影响。
川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(Wu et al., 2010), 森林群落的自然更新以林窗更新为主(吴庆贵等, 2016)。一般情况下, 冬季冻结环境下凋落物分解等过程处于“停滞”状态(Olson, 1963); 但越来越多的研究表明, 高寒地区凋落物分解主要发生在冬季(Hobbie, 1996; Aerts, 1997, 2006; Gavazov, 2010)。Hobbie和Chapin (1996)研究发现, 寒冷地区凋落物分解主要发生在第1年, 第2年以后以冬季分解为主。而川西亚高山和高山区域岷江冷杉新鲜凋落叶第1年质量损失的53%-64%出现在冬季(Wu et al., 2010; 吴庆贵等, 2016)。与我们的假设一致, 4年的分解结果显示, 岷江冷杉枝条第1年质量的总损失量为59.7%-67.6% (图6), 且枝条冬季总质量损失比例明显高于生长季节。这主要是由于亚高山和高山森林季节性雪被和冻融循环特征及耐寒生物种群活动明显促进了寒冷季节的凋落物分解(Aerts, 2006; Wu et al., 2010)。与此同时, 林窗对土壤冻融格局、雪被环境(Varhola et al., 2010)以及耐寒微生物活性的改变也影响非生长季节枝条的分解。另一方面, Olson模型对凋落物分解系数(k)的拟合结果显示, 林窗和林下枝凋落物分解系数(k = 0.120-0.135)显著低于同区域叶凋落物(k = 0.201-0.328)和温带落叶森林枝凋落物(Cha et al., 2017), 高于亚热带森林林窗枝条(Zhang & Zak, 1995)和欧洲北方森林粗枝(Shorohova & Kapitsa, 2016)。这是因为: 与凋落叶相比, 尽管亚高山森林冬季和生长季节截然不同的环境特征同样有利于凋落枝易分解组分的快速降解, 但由于其木质素含量以及C:N和木质素:N等初始质量特征明显高于凋落叶, 导致枝条的分解更加依赖于土壤微生物的酶促降解过程(Berg & McClaugherty, 2014)。可见, 冬季是亚高山森林凋落物分解的重要阶段, 凋落枝在亚高山森林系统分解缓慢。
凋落物的分解是一个整体而连续的过程, 林窗对凋落物分解的影响常常随林窗面积大小和方位而发生变化, 这导致迄今为止的研究结果仍具有很大的不确定性(Zhu et al., 2015)。Sariyildiz (2008)在土耳其北部高山森林的分解实验发现, 林下或小林窗下凋落物的分解速率显著高于大林窗下凋落物的分解速率; Denslow等(1998)在湿润热带森林的实验显示, 林窗面积大小与凋落物分解速率之间并没有显著的相关关系; 吴庆贵等(2016)在同区域1年的分解实验表明, 冷杉凋落叶在生长季节的质量损失速率随林窗面积增加而减小, 在非生长季节(冬季)的质量损失速率随林窗面积增加而增加。4年的分解实验显示, 林窗并未促进枝条凋落物的分解, 林下凋落物的质量残留量显著低于林窗, 林下枝条具有更高的分解系数(图4), 分解速率至实验结束无显著差异(图5)。与我们的假设一致, 冷杉凋落枝质量损失速率同样在第1年分解的生长季节随林窗面积增加而减小, 在非生长季节(冬季)随林窗面积增加而增加。值得注意的是, 林窗和林下凋落枝条残留量和分解速率并未表现出与Sariyildiz (2008)以及Zhang和Zak (1995)研究结果一致的变化规律(林下> FG3> FG2> FG1), 枝条残留量和分解速率随分解持续不断变化, 林下与小林窗在不同分解时期表现出明显差异。一方面, 林窗面积越大, 冬季形成雪被的厚度比生长季节的积雪雪被厚, 林窗内雪被覆盖的时间也越长, 季节性雪被具有保温的作用, 使得较大林窗下凋落物的平均温度比其他林窗下要高, 这就为凋落物分解提供了绝热保温且相对稳定的分解环境, 加速了凋落枝的分解。另一方面, 在生长季节, 由于光线可更大程度地照射到地面使得凋落物平均温度升高, 促进了大林窗内凋落物水分的蒸发, 干湿交替过程可能不利于土壤无脊椎动物和微生物对凋落物的分解(Devi et al., 2012), 进而抑制凋落叶分解过程。这进一步说明林窗调控的环境变化特征深刻影响亚高山森林冬季和生长季节凋落物分解过程。
综上所述, 冬季是高海拔森林物质循环的重要时期, 林窗面积大小能改变林窗和林下凋落枝分解的雪被厚度、温度和冻融循环频次。川西亚高山森林林窗和林下凋落枝初期(4年)的质量损失量以第1年最高, 且以冬季损失为主, 林窗面积大小对亚高山森林凋落枝质量损失的影响随分解时间而有所不同。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/j.1365-2486.2008.01674.xURL [本文引用: 1]

As atmospheric CO 2 increases, ecosystem carbon sequestration will largely depend on how global changes in climate will alter the balance between net primary production and decomposition. The response of primary production to climatic change has been examined using well-validated mechanistic models, but the same is not true for decomposition, a primary source of atmospheric CO 2 . We used the Long-term Intersite Decomposition Experiment Team (LIDET) dataset and model-selection techniques to choose and parameterize a model that describes global patterns of litter decomposition. Mass loss was best represented by a three-pool negative exponential model, with a rapidly decomposing labile pool, an intermediate pool representing cellulose, and a recalcitrant pool. The initial litter lignin/nitrogen ratio defined the size of labile and intermediate pools. Lignin content determined the size of the recalcitrant pool. The decomposition rate of all pools was modified by climate, but the intermediate pool's decomposition rate was also controlled by relative amounts of litter cellulose and lignin (indicative of lignin-encrusted cellulose). The effect of climate on decomposition was best represented by a composite variable that multiplied a water-stress function by the Lloyd and Taylor variable Q 10 temperature function. Although our model explained nearly 70% of the variation in LIDET data, we observed systematic deviations from model predictions. Below- and aboveground material decomposed at notably different rates, depending on the decomposition stage. Decomposition in certain ecosystem-specific environmental conditions was not well represented by our model; this included roots in very wet and cold soils, and aboveground litter in N-rich and arid sites. Despite these limitations, our model may still be extremely useful for global modeling efforts, because it accurately ( R 2 =0.6804) described general patterns of long-term global decomposition for a wide array of litter types, using relatively minimal climatic and litter quality data.
DOI:10.1039/b612546hURL [本文引用: 3]
Litter decomposition is an important component of the global carbon budget. Due to the strong climatic control of litter decomposition, climate change may significantly affect this pathway. This review quantifies the climatic influences on litter decomposition rates, both directly and indirectly through effects on litter chemistry. To this end, I analysed first-year leaf litter decomposition data from 44 locations, ranging from cool temperate sites to humid tropical sites. Actual evapotranspiration (AET) was used as an index for the climatic control on decomposition. As litter chemistry parameters I included N and P concentrations, C/N and C/P ratios, lignin concentrations, and lignin/N and lignin/P ratios. At a global scale, climate (expressed as AET) is the best predictor for the decomposition constants (ic-values) of the litter, whereas litter chemistry parameters have much lower predictive values. Path analysis showed that the control of AET on litter decomposability is partly mediated through an indirect effect of AET on litter chemistry. Thus, the relation between climate, leaf litter chemistry and leaf litter decomposition is a triangular relationship. Mean AET in the humid tropical region is three times as high as in both the temperate and the Mediterranean region and this results in a more than six-fold increase in mean k-values. However, due to the large variability in k-values within each region there is a considerable overlap in k-values between the tropics and the other climatic regions. Within a particular climatic region litter chemistry parameters are the best predictors of k-values, especially in the tropics, whereas the percentage of variance in k-values explained by AET is low or absent. In general, litters from the tropical sites have higher N concentrations and lower lignin/N ratios than litters from other climatic regions. In both the tropics and in the Mediterranean region, the lignin/N ratio is the best chemical predictor of litter decomposab
DOI:10.1111/j.1365-2745.2006.01142.xURL [本文引用: 4]
Summary Top of page Summary Introduction Direct effects of temperature Indirect temperature effects on litter chemistry Indirect effects on detritivore and decomposer communities How to proceed? Acknowledgements References 1 Decomposition of plant litter, a key component of the global carbon budget, is hierarchically controlled by the triad: climate > litter quality > soil organisms. Given the sensitivity of decomposition to temperature, especially in cold biomes, it has been hypothesized that global warming will lead to increased litter decomposition rates, both through direct temperature effects and through indirect effects on litter quality and soil organisms. 2 A meta-analysis of experimental warming studies in cold biomes (34 site-species combinations) showed that warming resulted in slightly increased decomposition rates. However, this response was strongly dependent on the method used: open top chambers reduced decomposition rates, whereas heating lamps stimulated decomposition rates. The low responsiveness was mainly due to moisture-limited decomposition rates in the warming treatments, especially at mesic and xeric sites. This control of litter decomposition by both temperature and moisture was corroborated by natural gradient studies. 3 Interspecific differences in litter quality and decomposability are substantially larger than warming-induced phenotypic responses. Thus, the changes in the species composition and structure of plant communities that have been observed in medium-term warming studies in cold biomes will have a considerably greater impact on ecosystem litter decomposition than phenotypic responses. 4 Soil fauna communities in cold biomes are responsive to climate warming. Moreover, temperature-driven migration of the, hitherto absent, large comminuters to high-latitude sites may significantly increase decomposition rates. However, we do not know how far-reaching the consequences of changes in the species composition and structure of the soil community are for litter decomposition, as there is a lack of data on functional species redundancy and the species dispersal ability. 5 Global warming will lead to increased litter decomposition rates only if there is sufficient soil moisture. Hence, climate scenario and experimental studies should focus more on both factors and their interaction. As interspecific differences in potential decomposability and litter chemistry are substantially larger than phenotypic responses to warming, the focus of future research should be on the former. In addition, more light should be shed on the below-ground arkness to evaluate the ecological significance of warming-induced soil fauna community changes for litter decomposition processes in cold biomes.
DOI:10.1023/A:1004828221756URL
We examined the effects of treefall gap size and soil properties on microbial biomass dynamics in an undisturbed mature-phase humid subtropical broadleaved forest in north-east India. Canopy gaps had low soil moisture and low microbial biomass suggesting that belowground dynamics accompanied changes in light resources after canopy opening. High rainfall in the region causes excessive erosion/leaching of top soil and eventually soil fertility declines in treefall gaps compared to understorey. Soil microbial population was less during periods when temperature and moisture conditions are low, while it peaked during rainy season when the litter decomposition rate is at its peak on the forest floor. Greater demand for nutrients by plants during rainy season (the peak vegetative growth period) limited the availability of nutrients to soil microbes and, therefore, low microbial C, N and P. Weak correlations were also obtained for the relationships between microbial C, N and P and soil physico hemical properties. Gap size did influence the microbial nutrients and their contribution to soil organic carbon, total Kjeldhal nitrogen and available-P. Contribution of microbial C to soil organic carbon, microbial N to total nitrogen were similar in both treefall gaps and understorey plots, while the contribution of microbial P to soil available-P was lower in gap compared to the understorey. These results indicate that any fluctuation in microbial biomass related nutrient cycling processes in conjunction with the associated microclimate variation may affect the pattern of regeneration of tree seedlings in the gaps and hence be related with their size.
DOI:10.1007/s11104-009-0119-6URL [本文引用: 4]
We assessed direct and indirect effects of snow cover on litter decomposition and litter nitrogen release in alpine tundra. Direct effects are driven by the direct influence of snow cover on edaphoclimatic conditions, whereas indirect effects result from the filtering effect of snow cover on species abundance and traits. We compared the in situ decomposition of leaf litter from four dominant plant species (two graminoids, two shrubs) at early and late snowmelt locations using a two-year litter-bag experiment. A seasonal experiment was also performed to estimate the relative importance of winter and summer decomposition. We found that growth form (graminoids vs. shrubs) are the main determinants of decomposition rate. Direct effect of snow cover exerted only a secondary influence. Whatever the species, early snowmelt locations showed consistently reduced decomposition rates and delayed final stages of N mineralization. This lower decomposition rate was associated with freezing soil temperatures during winter. The results suggest that a reduced snow cover may have a weak and immediate direct effect on litter decomposition rates and N availability in alpine tundra. A much larger impact on nutrient cycling is likely to be mediated by longer term changes in the relative abundance of lignin-rich dwarf shrubs.
[本文引用: 3]
DOI:10.3390/f8050176URL [本文引用: 3]
Woody debris, which is supplied by branch litter, is an important component of forest ecosystems as it contains large quantities of organic matter and nutrients. We evaluated changes in branch wood dry weight and nutrient content of six common species (Fraxinus rhynchophylla, Pinus densiflora, Prunus sargentii, Quercus mongolica, Acer pseudosieboldianum, and Symplocos chinensis for. pilosa) in a deciduous temperate forest in Korea for 40 months. Branch wood disk samples 1.4 1.6 cm thick were cut, and mass loss was measured over time using the litterbag method. No significant differences in mass loss were recorded among the six tree species. Further, mass loss was negatively correlated with initial lignin concentration and positively correlated with both initial cellulose concentration and wood density for each species. Species with high wood cellulose content had high wood density while the lignin content in wood was relatively low. Accordingly, cellulose contributed to wood density, creating a relatively lower lignin content, and the decreased lignin concentration increased the wood decomposition rate.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2486.2009.02115.xURL [本文引用: 2]
Forests in northeastern North America are influenced by varying climatic and biotic factors; however, there is concern that rapid changes in these factors may lead to important changes in ecosystem processes such as decomposition. Climate change (especially warming) is predicted to increase rates of decomposition in northern latitudes. Warming in winter may result in complex effects including decreased levels of snow cover and an increased incidence of soil freezing that will effect decomposition. Along with these changes in climate, moose densities have also been increasing in this region, likely affecting nutrient dynamics. We measured decomposition and N release from 15N-labeled sugar maple leaf litter and moose feces over 20 months in reference and snow removal treatment (to induce soil freezing) plots in two separate experiments at the Hubbard Brook Experimental Forest in New Hampshire, USA. Snow removal/soil freezing decreased decomposition of maple litter, but stimulated N transfer to soil and microbial biomass. Feces decomposed more rapidly than maple litter, and feces N moved into the mineral soil more than N derived from litter, likely due to the lower C : N ratio of feces. Feces decomposition was not affected by the snow removal treatment. Total microbial biomass (measured as microbial N and C) was not significantly affected by the treatments in either the litter or feces plots. These results suggest that increases in soil freezing and/or large herbivore populations, increase the transfer rate of N from plant detritus or digested plants into the mineral soil. Such changes suggest that altering the spatial and temporal patterns of soil freezing and moose density have important implications for ecosystem N cycling.
DOI:10.1046/j.1365-2745.1998.00295.xURL [本文引用: 1]
Abstract 1 We examined the effects of variation in gap size on above- and below-ground light and nutrient processes in a tropical wet forest in Costa Rica. 2 Trees were felled to create canopy openings ranging in size from 65 to 611m 2 . Following treefall, we measured initial litter mass in the crown zone of six gaps. During the subsequent year, we measured litter decomposition rate and light levels as well as NH 4 -N, NO 3 -N and PO 4 -P levels in surface soils, soil moisture and fine root mass. We also measured growth rates of fertilized and non-fertilized plants of four species of Miconia (Melastomataceae) to assess nutrient limitation to plant growth in large gaps. 3 Light levels in the centres of gaps were significantly related to size of the canopy opening. After 1 year, light levels near the ground in larger openings (35 40% full sunlight immediately after treefall) declined to levels similar to those in smaller gaps (10 20%). 4 Although canopy opening had only slight effects on soil NH 4 -N, NO 3 -N pools were significantly greater in gaps than in understorey at both sites. The effect was positively correlated with gap size. Extractable PO 4 -P was also greater in gaps than in adjacent understorey, although the difference did not vary as a function of gap size. In three of six gaps, fine root biomass was less in gaps than in adjacent understorey. 5 Of four species of Miconia , only M. affinis , a small tree common in early secondary forests, grew significantly faster in fertilized than in non-fertilized treatments in three large gaps. 6 Our data suggest that higher nutrient pools in surface soils of treefall gaps may result from decomposition and mineralization of the large mass of fresh litter from the fallen tree. While growth rates of shade-tolerant rain forest trees and shrubs are not much affected by the increased nutrient availability even at high light levels, growth rates of pioneer or high-light demanding species may be enhanced by increased above- and below-ground resources.
URL [本文引用: 2]
ABSTRACT Population study of collembola for one calendar year was made in relation with moisture, organic carbon and nitrogen contents of a sub-tropical forest floor upto 0-10 cm. soil layer. Soil moisture content played important role and developed a significant correlation (p>0.01) with the monthly population density of collembola. With increase carbon and nitrogen contents also observe increase population density indicating collembola's active participation in the release of these two nutrients from litter through decomposition and its incorporation in soil fertility.
DOI:10.3390/f8090314URL [本文引用: 1]
Litterfall is the primary source of carbon and nutrients that determine soil fertility in forest ecosystems. Most current studies have focused on foliar litter, but the seasonal dynamics and allometric scaling relationships among different litter components (e.g., foliar litter, woody litter, reproductive litter, and epiphytic litter) are poorly understood. Here, we investigated the litter production of various litter components in a sub-alpine spruce-fir forest on the eastern Tibetan Plateau based on one year of observations (from August 2015 to July 2016). Our results showed that total litter production (LT) was 2380 kg·ha611·year611 (3% of the aboveground forest biomass), of which 73.6% was foliar litter (LF), 15.6% was woody litter (LW), 3.0% was reproductive litter (LR), 1.3% was epiphytic litter (LE), and 6.5% was miscellaneous material (LM). The total litterfall was bimodal (with peaks occurring in April and October) and was dominated by tree species (85.4% of LT, whereas shrubs accounted for 6.8% of LT). The litter production of evergreen species (68.4% of LT) was higher than that of deciduous species (23.8% of LT). Isometric relationships were observed between litter components and the total litter (i.e., LF∝LT0.99≈1 and LR∝LT0.98≈1), and allometric relationships were also found (i.e., LW∝LT1.40>1 and LM∝LT0.82<1). However, because some components did not exhibit obvious seasonal dynamics (i.e., LE), some relationships could not be expressed using allometric equations (i.e., LE versus LT, LF versus LE, LW versus LE, and LE versus LM). Thus, the different litter components showed different seasonal dynamics, and the total litter dynamics were primarily determined by the variation in foliar litter. In addition, the allometric relationships of the forest litterfall varied with the litter components, functional types (evergreen versus deciduous) and vertical structures (tree versus shrub). This study provides basic data and a new insight for future plant litter studies.
DOI:10.1007/s11104-010-0477-0URL [本文引用: 2]
Climatic changes resulting from anthropogenic activities over the passed century are repeatedly reported to alter the functioning of pristine ecosystems worldwide, and especially those in cold biomes. Available literature on the process of plant leaf litter decomposition in the temperate Alpine zone is reviewed here, with emphasis on both direct and indirect effects of climate change phenomena on rates of litter decay. Weighing the impact of biotic and abiotic processes governing litter mass loss, it appears that an immediate intensification of decomposition rates due to temperature rise can be retarded by decreased soil moisture, insufficient snow cover insulation, and shrub expansion in the Alpine zone. This tentative conclusion, remains speculative unless empirically tested, but it has profound implications for understanding the biogeochemical cycling in the Alpine vegetation belt, and its potential role as a buffering mechanism to climate change.
DOI:10.2307/1942193URL [本文引用: 1]
Rates of weight loss and nutrient release (N, P, S, K, Mn, Ca, Zn, Fe, Mn, Cu, Na) were measured in decomposing leaf and branch tissue form yellow birch, sugar maple, and beech, and in branch tissue from red spruce and balsam fir. Neither leaf nor branch decomposition differed significantly over an elevational range of 220 m. Decomposition rates for leaves varied with yellow birch > sugar maple > beech. The decomposition rate for hardwood branches was greater than that for conifer branches, but differences between hardwoods were not significant. Maximum decomposition rates occurred during the summer for both branch and leaf tissue. The rate of nutrient release from decomposing branch and leaf litter appears to be correlated with nutrient concentration in current litter fall, precipitation, and leaf wash. The concentration and absolute weight of N. S. and P in the leaf litter of all species increased with time. The amount of the increase as well as the initiation of nutrient release was influenced by C: element ratios in the leaf tissue. These studies also indicate that P levels can influence the mineralization or immobilization of other important nutrients. Carbon-to-element ratios in decomposing litter varied between species and elevation at different times of the year, but element: P ratios were much more uniform. In branch tissue the physical loss of N- and P-rich bark and buds offset any increase in concentration that would have occurred through decomposition. Potassium and magnesium were rapidly released from the litter by leaching. Similar minimum concentrations in leaf tissue indicate that critical C: element ratios also exist for these elements. Calcium release was similar to dry weight loss, indicating that it is a structural component primarily released by decomposition. Maximum nutrient release from current litter occurred in the autumn and summer. It was not correlated with the nutrient output from the ecosystem which occurred primarily during the spring. The net output of Ca, Mg, and K from the watershed was very small compared to quantities released from current litter. Factors which contribute to the complex nature of decomposition are: seasonal heterotroph activity, heterotroph nutrient demand, environmental conditions regulations heterotroph activity, species tissue palatability, species composition of litter, tissue composition of litter, nutrient content of litter, nutrient mobility, and nutrient input (i.e., leafwash, litter fall).
DOI:10.11707/j.1001-7488.20160411URLMagsci [本文引用: 2]
林窗是森林生态系统中的一种中小尺度干扰,是促进森林更新、养分循环、功能提高的重要推动力。本文阐述林窗生态学的研究进展与展望,以期为今后的林窗理论研究和森林经营实践提供参考。林窗生态学研究集中于林窗的形成、基本特征以及林窗对森林小气候和植物群落特征等地上结构与过程的影响;近年来,林窗对细根与枯落物分解、土壤碳氮动态及酶活性以及对森林动物和土壤微生物的生理生态学特征影响研究逐渐增多,但研究的时空尺度较小且不够全面、深入。今后应着重研究林窗如何调控林分结构和森林生态服务过程与功能,重点阐明林窗对土壤碳氮分配、循环和固持,细根分解及根际效应等地下生态过程,以及对动植物与微生物生理生态学的影响与影响机制;同时,应进一步拓展研究的时空尺度,并加强地上与地下生态系统、生物与非生物因子、宏观与微观尺度等的整合研究。
DOI:10.11707/j.1001-7488.20160411URLMagsci [本文引用: 2]
林窗是森林生态系统中的一种中小尺度干扰,是促进森林更新、养分循环、功能提高的重要推动力。本文阐述林窗生态学的研究进展与展望,以期为今后的林窗理论研究和森林经营实践提供参考。林窗生态学研究集中于林窗的形成、基本特征以及林窗对森林小气候和植物群落特征等地上结构与过程的影响;近年来,林窗对细根与枯落物分解、土壤碳氮动态及酶活性以及对森林动物和土壤微生物的生理生态学特征影响研究逐渐增多,但研究的时空尺度较小且不够全面、深入。今后应着重研究林窗如何调控林分结构和森林生态服务过程与功能,重点阐明林窗对土壤碳氮分配、循环和固持,细根分解及根际效应等地下生态过程,以及对动植物与微生物生理生态学的影响与影响机制;同时,应进一步拓展研究的时空尺度,并加强地上与地下生态系统、生物与非生物因子、宏观与微观尺度等的整合研究。
DOI:10.1007/s10021-015-9921-6URL [本文引用: 3]
To understand the effects of forest gaps on lignin degradation during shrub foliar litter decomposition, a field litterbag experiment was conducted in an alpine fir (Abies faxoniana) forest of the eastern Tibet Plateau. Dwarf bamboo (Fargesia nitida) and willow (Salix paraplesia) foliar litterbags were placed on the forest floor from the gap center to the closed canopy. The litterbags were sampled during snow formation, snow coverage, snow melting and the growing season from October 2010 to October 2012. The lignin concentrations and loss in the litter were measured. Over 2 years, lignin loss was lower in the bamboo litter (34.64 43.89%) than in the willow litter (38.91 55.10%). In the bamboo litter, lignin loss mainly occurred during the first decomposition year, whereas it occurred during the second decomposition year in the willow litter. Both bamboo and willow litter lignin loss decreased from the gap center to the closed canopy during the first year and over the entire 2-year decomposition period. Compared with the closed canopy, the gap center showed higher lignin loss for both bamboo and willow litter during the two winters, but lower lignin loss during the early growing period. Additionally, the dynamics of microbial biomass carbon during litter decomposition followed the same trend as litter lignin loss during the two winters and growing period. These results indicated that alpine forest gaps had significant effects on shrub litter lignin loss and that reduced snow cover during winter warming would inhibit shrub lignin degradation in this alpine forest.
DOI:10.3724/SP.J.1258.2013.00030URL [本文引用: 2]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)$1]康定柳(Satix paraplesia)凋落物在沿林窗一林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.351%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOI:10.3724/SP.J.1258.2013.00030URL [本文引用: 2]
高山/亚高山森林灌木层植物凋落物的分解对于系统物质循环等过程具有重要意义,并可能受到冬季不同厚度雪被斑块下冻融格局的影响。该文采用凋落物分解袋法,研究了高山森林典型灌层植物华西箭竹(Fargesia nitida)$1]康定柳(Satix paraplesia)凋落物在沿林窗一林下形成的冬季雪被厚度梯度(厚型雪被斑块、较厚型雪被斑块、中型雪被斑块、薄型雪被斑块、无雪被斑块)上在第一年不同关键时期(冻结初期、冻结期、融化期、生长季节初期和生长季节后期)的质量损失特征。在整个冻融季节,华西箭竹和康定柳凋落叶的平均质量损失分别占全年的(48.78±2.351%和(46.60±5.02)%。冻融季节雪被覆盖斑块下凋落叶的失重率表现出厚型雪被斑块大于薄型雪被斑块的趋势,而生长季节无雪被斑块的失重率明显较高。尽管如此,华西箭竹凋落物第一年分解表现出随冬季雪被厚度增加而增加的趋势,但康定柳凋落物第一年失重率以薄型雪被斑块最高,而无雪被斑块最低。同时,相关分析表明冻融季节凋落叶的失重率与平均温度和负积温呈极显著正相关,生长季节凋落叶的失重率与所调查的温度因子并无显著相关关系,但全年凋落物失重率与平均温度和正/负积温均显著相关。这些结果清晰地表明,未来冬季变暖情境下高山森林冬季雪被格局的改变将显著影响灌层植物凋落物分解,影响趋势随着物种的差异具有明显差异。
DOI:10.1007/s11104-015-2479-4URL [本文引用: 2]
Forest gap manipulates the hydrothermal dynamics of the forest floor, creating heterogeneous microenvironments and controlling understory ecosystem processes. However, how the heterogeneity in environ
DOI:10.2307/2963492URL [本文引用: 1]
I compared effects of increased temperature and litter from different Alaskan tundra plant species on cycling of carbon and nitrogen through Litter and soil in microcosms. Warming between 4 degrees and 10 degrees C significantly increased rates of soil and litter respiration, litter decomposition, litter nitrogen release, and soil net nitrogen mineralization. Thus, future warming will directly increase rates of carbon and nitrogen cycling through litter and soil in tundra. In addition, differences among species' litter in rates of decomposition, N release, and effects on soil net nitrogen mineralization were sometimes larger than differences between the two temperature treatments within a species. Thus, changes in plant community structure and composition associated with future warming will have important consequences for how elements cycle through litter and soil in tundra. In general, species within a growth form (graminoids, evergreen shrubs, deciduous shrubs, and mosses) were more similar in their effects on decomposition than were species belonging to different growth forms, with graminoid litter having the fastest rate and litter of deciduous shrubs and mosses having the slowest rates. Differences in rates of litter decomposition were more related to carbon quality than to nitrogen concentration. Increased abundance of deciduous shrubs with future climate warming will promote carbon storage, because of their relatively large allocation to woody stems that decompose slowly. Changes in moss abundance will also have important consequences for future carbon and nitrogen cycling, since moss litter is extremely recalcitrant and has a low potential to immobilize nitrogen.
DOI:10.1007/BF02179958URL
Mass and nitrogen (N) dynamics of leaf litter measured in Alaskan tussock tundra differed greatly from measurements of these processes made in temperate ecosystems. Nearly all litter mass and N loss occurred during the winter when soils were mostly frozen. Litter lost mass during the first summer, but during the subsequent two summers when biological activity was presumably higher than it is during winter, litter mass remained constant and litter immobilized N. By contrast, litter lost significant mass and N over both winters of measurement. Mass loss and N dynamics were unaffected by microsite variation in soil temperature and moisture. Whether wintertime mass and N loss resulted from biological activity during winter or from physical processes (e.g., fragmentation or leaching) associated with freeze-thaw is unknown, but has implications for how future climate warming will alter carbon (C) and N cycling in tundra. We hypothesize that spring runoff over permafrost as soils melt results in significant losses of C and N from litter, consistent with the observed influx of terrestrial organic matter to tundra lakes and streams after snow melt and the strong N limitation of terrestrial primary production.
DOI:10.1016/j.apsoil.2007.01.003URL [本文引用: 1]
The effect of temperature changes on soil communities is an important aspect when estimating the effects of a predicted climate change. The aim of this investigation was to increase knowledge on how freeze-thaw cycles alter the soil microarthropod community in the sub-arctic. The abundance of springtails and mites was investigated after three seasons of prolonged periods of freeze-thaw cycles in the field, and the presence or absence of migration barriers, at two different field sites. Dome shaped transparent plastic greenhouses were successfully used as a novel method to increase freeze-thaw cycle frequencies in the soil. At a fellfield site, freeze-thaw treatment did not lead to significant differences in the five main soil faunal groups, but increased abundance were seen in a number of separate taxa. There was no freeze-thaw treatment effect on soil microbial biomass or soil nutrients, although treatments interacted as inorganic N increased in the separate freeze-thaw and migration barrier treatments. By contrast, at a glade site responses were strong due to more pronounced increases in the number of freeze-thaw cycles. The highest numbers of Collembola after 2 years of treatment were found in the freeze-thaw plots, in combination with migration barriers. The freeze-thaw treatment here also resulted in more Oribatida, microbial biomass C and dissolved organic C. A common hypothesis is that an increased number of freeze-thaw cycles would result in elevated winter mortality in microarthropods due to increased risk of inoculative freezing. However, we observed no increased mortality due to freeze-thaw events. Rather, there was a stimulation of soil microarthropods and microbial biomass, perhaps due to a prolonged period of microbial and faunal activity when the soil is repeatedly frozen and thawed compared to a constantly frozen soil.
DOI:10.3321/j.issn:1006-687X.2002.05.002URL [本文引用: 2]
分析了云南西北部碧塔海自然保护区的亚高山针叶林林窗大小对林窗植被和更新的影响 .调查区域出现I级林窗 (面积 5 0m2 )占 36 % ,II级 (5 0~ 75m2 ) 2 1% ,III级 (75~ 10 0m2 ) 19% ,IV级 (10 0~ 12 5m2 ) 13%和V级 (12 5m2 ) 11% .结果表明 ,亚高山针叶林因林窗大小不同而导致小环境条件的差异 ,使林窗植被和更新与林窗干扰密切相关 .林窗大小对林窗植被的物种多样性、结构、密度分布以及云冷杉更新苗的空间分布都有不同程度的影响 .小林窗(I级 )的物种多样性比大林窗 (IV级和V级 )、中林窗 (II级和III级 )高 .云冷杉更新幼苗以中、小林窗较多 ,且在小林窗中呈随机分布 ,主要分布在林窗中心和过渡区域 (88% ) ;在大、中林窗中则呈集群分布 ,主要分布在过渡区域 ,占4 4 %~ 5 0 % .同时 ,研究表明该区亚高山针叶林有效更新的林窗阈值面积大约为 75m2 .图 4表 3参 2 2
DOI:10.3321/j.issn:1006-687X.2002.05.002URL [本文引用: 2]
分析了云南西北部碧塔海自然保护区的亚高山针叶林林窗大小对林窗植被和更新的影响 .调查区域出现I级林窗 (面积 5 0m2 )占 36 % ,II级 (5 0~ 75m2 ) 2 1% ,III级 (75~ 10 0m2 ) 19% ,IV级 (10 0~ 12 5m2 ) 13%和V级 (12 5m2 ) 11% .结果表明 ,亚高山针叶林因林窗大小不同而导致小环境条件的差异 ,使林窗植被和更新与林窗干扰密切相关 .林窗大小对林窗植被的物种多样性、结构、密度分布以及云冷杉更新苗的空间分布都有不同程度的影响 .小林窗(I级 )的物种多样性比大林窗 (IV级和V级 )、中林窗 (II级和III级 )高 .云冷杉更新幼苗以中、小林窗较多 ,且在小林窗中呈随机分布 ,主要分布在林窗中心和过渡区域 (88% ) ;在大、中林窗中则呈集群分布 ,主要分布在过渡区域 ,占4 4 %~ 5 0 % .同时 ,研究表明该区亚高山针叶林有效更新的林窗阈值面积大约为 75m2 .图 4表 3参 2 2
DOI:10.1139/er-9-1-1URL [本文引用: 2]
Small-scale gap disturbance in forests is reviewed. Caused by the death of individual or multiple trees with subsequent fall from the canopy, gaps have been extensively studied in temperate deciduous and tropical forests for the past 20 years. This review considers much of this research with a view to assessing the importance of gap disturbance in boreal forests. Because of the ubiquity of large-scale, stand-initiating disturbances such as landscape-level fires, epidemic insect outbreaks, and periodic extensive windthrow events, gap processes in boreal forests have received little attention. Research in the Scandinavian and Russian boreal forest, as well as in high-altitude boreal "outliers" found in Japan and the United States, is showing that gap disturbance determines forest structure and processes to a greater extent than previously assumed. Boreal forests dominated by the shade-tolerant fir (–spruce () complex are particularly well-adapted to the development of long-term, old-growth continuity in the absence of large-scale disturbance. : gap dynamics, disturbance, boreal forests, temperate forests, tropical forests, silviculture.L'auteur présente une revue sur les ouvertures par perturbations à petite échelle en milieu forestier. Occasionn ées par la mort d'un ou plusieurs arbres suivie de leur chute de la canopée, ces ouvertures ont été largement étudiées en foréts décidues tempérées et tropicales au cours des 20 dernières années. Cette revue prend en compte une bonne partie de cette recherche pour évaluer l'importance de l'ouverture par perturbation en for éts boréales. 08 cause de l'ubiquité des perturbations à grande échelle, initiatrices de peuplements, telles que les incendies au niveau du paysage, les épidémies d'insectes et les évènements périodiques de grands chablis, les processus d'ouverture en forét boréale n'ont re04u que peu d'attention. Des recherches conduites dans les foréts boréales de Scandinavie et de Russie, ainsi que dans des stations subalpines au Japon et aux 07tats-Unis, montrent que l'ouverture par perturbation détermine la structure de la forét et de ses processus beaucoup plus qu'on l'avait jusqu'ici assumé. Les foréts boréales dominées par le complexe sapin tolérant ()–épinette () sont particulièrement bien adaptées au développement de foréts surannées en continuité et à long terme, en absence de perturbation à grande échelle. Mots clés : dynamique des ouvertures, perturbation, for éts boréales, foréts tempérées, foréts tropicale, sylviculture. [Traduit par la Rédaction]
DOI:10.2307/1932179URL [本文引用: 2]
See full-text article at JSTOR
DOI:10.1126/science.1134853URL [本文引用: 1]
DOI:10.1139/cjfr-30-11-1751URL [本文引用: 1]
The rate of mass loss of three standard litter substrates was measured in litter bag studies in forests and adjacent clearcuts at 21 sites throughout British Columbia, Canada, to test the hypotheses that (i) rates of mass loss are greater in clearcuts than in forests, and (ii) clear felling would stimulate decomposition most in colder zones. The standard substrates used were lodgepole pine (Pin...
DOI:10.1139/x03-152URL [本文引用: 1]
DOI:10.1016/j.electacta.2009.11.058URL [本文引用: 2]
Effects of gap-size classes on litter decomposition rates were investigated in a high-elevation forest for 4 years by placing leaf litter of beech (Fagus orientalis Lipsky.), oak (Quercus robur L.), and chestnut (Castanea sativa Mill.) in (1) a closed canopy, (2) small gaps with a diameter of less than 15 m, (3) intermediate gaps with a diameter of 15-30 m, and (4) large gaps with a diameter of more than 30 m. The leaf litter placed under the closed canopy or within the small gaps decayed more rapidly than those in the intermediate or the large gaps for all three deciduous species. Among the microclimatic and soil factors, soil temperature was most strongly positively correlated with litter decomposition rates, and among the three species, initial lignin concentration was the best predictor of litter decomposition rates. Differences in litter decomposition rates among the four gap-size classes were generally significant, but varied among the three species. Litter with low lignin concentrations tended to be more responsive to canopy openings. Large forest gaps significantly reduced litter decomposition rates in this study by changing environmental conditions, especially by decreasing soil temperature and soil pH which reduced soil respiration rates. These changes may result in reduced nutrient cycling, carbon cycling, and organic matter turnover rates in these forest ecosystems.
DOI:10.1016/j.foreco.2011.01.011URL [本文引用: 1]
As silvicultural objectives have changed over the last several decades, managers are increasingly designing cutting regimes that mimic natural disturbance with the hopes that such systems will restore forests to a more natural condition while optimizing harvest yield. Treefall gaps, canopy openings caused by the death of one or more trees, are the dominant form of disturbance in many forest systems worldwide. These gaps play an important role in forest ecology by helping to maintain bio- and pedo-diversity, influencing nutrient cycling, and preserving the uneven-age nature of late-successional forests. In gap literature, there are inconsistencies with regard to gap terminology, methods for identifying and studying gaps, and modeling gap disturbances. From the papers reviewed, the size of treefall gaps ranges widely from 10 to >5000 m 2; we suggest that the maximum gap size should be set at 1000 m 2. Larger openings tend to have microclimates and return intervals significantly different than smaller treefall gaps. Two main definitions of treefall gaps exist: canopy gap: a ole in the forest through all levels down to an average height of 2 m above ground and extended gap: canopy gap plus the area that extends to the bases of surrounding canopy trees. Although researchers have assumed a variety of gap shapes to simplify measuring gap size, gaps are often irregularly shaped and so we recommend that gap areas and shapes be determined from detailed field measurements. Gap age may be determined from tree ring analysis of released trees in or near the gap edge, the spacing of whorls on released saplings, or from decomposition of gap-making trees. Windthrow is the main cause of canopy gaps in a variety of ecosystems; other causes include insects, diseases, acidic deposition, drought, and climate change. Treefall-gap models have been developed to predict the following processes during gap making or infilling: (i) gap abundance, (ii) forest structure, (iii) spatial and temporal variations in light levels, (iv) canopy dynamics, and (v) soil nutrient and water regimes. We recommend a protocol for gap studies and identify future research topics.
DOI:10.1007/s10342-015-0913-zURL [本文引用: 3]
An increasing number of studies provide evidence that mixed-species stands can overyield monocultures. But it is still hardly understood, how the overyielding at the stand level emerges from the...
DOI:10.1016/j.jhydrol.2010.08.009URL [本文引用: 2]
The past century has seen significant research comparing snow accumulation and ablation in forested and open sites. In this review we compile and standardize the results of previous empirical studies to generate statistical relations between changes in forest cover and the associated changes in snow accumulation and ablation rate. The analysis drew upon 33 articles documenting these relationships at 65 individual sites in North America and Europe from the 1930s to present. Changes in forest cover explained 57% and 72% of the variance of relative changes in snow accumulation and ablation, respectively. The incorporation of geographic and average historic climatic information did not significantly improve the ability to predict changes in snow processes, mainly because most of the studies did not provide enough information on site characteristics such as slope and aspect or meteorological conditions taking place during the experiments. Two simple linear models using forest cover as the sole predictor of changes in snow accumulation and ablation are provided, as well as a review of the main sources of variation that prevent the elaboration of more accurate multiple regression models. Further studies should provide detailed information regarding the main sources of variation influencing snow processes including the effect of year-to-year changes in weather variables during the monitoring period.
DOI:10.1016/j.actao.2009.11.002URL [本文引用: 5]
Mass loss and nutrient release of forest litter during the freeze–thaw season could play an essential role in C and nutrient cycling in cold regions, but few studies in some key ecosystems have been available. In order to characterize litter decomposition during the freeze–thaw season in a subalpine forest region of western China, a field experiment using the litterbag method was conducted on the decomposition of foliar litter of two dominant species, fir ( Abies faxoniana) and birch ( Betula platyphylla) under their respective forests. Over the freeze–thaw season following leaf-fall, about 18% and 20% of mass, 13% and 14% of lignin, 30% and 26% of cellulose, 14% and 21% of C, 30% and 27% of N, 17% and 15% of P, and 17% and 13% of K were lost from fir and birch litters, respectively. The lost mass and components accounted for more than 64% and 65% of mass, 72% and 69% of lignin, 75% and 60% of cellulose, 49% and 59% of C, 56% and 71% of N, 62% and 37% of P, and 38% and 37% of K in 1 year net loss rate of fir and birch litter, respectively. In addition, the loss of mass, lignin, cellulose and component bio-elements during the freeze–thaw season correlated closely with the initial substrate type and the levels of the individual bio-elements. The results demonstrated that litter decomposition during the freeze–thaw season contributes significantly to the first year decomposition in these subalpine forests.
[本文引用: 7]
[本文引用: 7]
DOI:10.3724/SP.J.1145.2013.00922URL [本文引用: 4]
了解高山森林林隙结构与形成机制对认识气候变化情景下的森林群落更新具有重要意义,但一直缺乏必要关注.通过典型样地法研究了川西高山岷江冷杉林隙结构特征和干扰状况.结果表明,林隙大小以中小型为主,密度为14.67个/hm-2,63.64%以折干形成.扩展林隙(EG)和林冠林隙(CG)分别占森林景观面积的12.60%和23.05%,干扰频率分别为115.25 m2 hm-2 a-1和63.02 m2 hm-2 a-1,林隙周转率为260.30 a.形成木径级集中于40-60 cm,树高主要在25-30 m,由单株形成木形成的林隙占50.09%;每个林隙约1.52株形成木,每株形成EG和CG面积分别为103.20 m2和56.43 m2.边界木平均胸径为50.16 cm,胸径结构分布曲线尖峰左偏,平均胸径和高度与EG和CG面积呈幂函数相关.综上所述,相对于亚高山针叶林,高山森林近20 a形成的林隙比例更高,形成木和边界木的高度更低但胸径更大,折干比例也更高,恶劣的环境条件(如风雪)可能是高山森林群落更新的主要原因.图5表4参33
DOI:10.3724/SP.J.1145.2013.00922URL [本文引用: 4]
了解高山森林林隙结构与形成机制对认识气候变化情景下的森林群落更新具有重要意义,但一直缺乏必要关注.通过典型样地法研究了川西高山岷江冷杉林隙结构特征和干扰状况.结果表明,林隙大小以中小型为主,密度为14.67个/hm-2,63.64%以折干形成.扩展林隙(EG)和林冠林隙(CG)分别占森林景观面积的12.60%和23.05%,干扰频率分别为115.25 m2 hm-2 a-1和63.02 m2 hm-2 a-1,林隙周转率为260.30 a.形成木径级集中于40-60 cm,树高主要在25-30 m,由单株形成木形成的林隙占50.09%;每个林隙约1.52株形成木,每株形成EG和CG面积分别为103.20 m2和56.43 m2.边界木平均胸径为50.16 cm,胸径结构分布曲线尖峰左偏,平均胸径和高度与EG和CG面积呈幂函数相关.综上所述,相对于亚高山针叶林,高山森林近20 a形成的林隙比例更高,形成木和边界木的高度更低但胸径更大,折干比例也更高,恶劣的环境条件(如风雪)可能是高山森林群落更新的主要原因.图5表4参33
DOI:10.1371/journal.pone.0097112URLPMID:4018275 [本文引用: 2]
Abstract There is increasing attention on the effects of seasonal snowpack on wintertime litter decomposition, as well as the processes following it, in cold biomes. However, little information is available on how litter nitrogen (N) dynamics vary with snowpack variations created by tree crown canopies in alpine forests. Therefore, to understand the effects of seasonal snowpack on litter N dynamics during different critical stages, litterbags with fir (Abies faxoniana), birch (Betula albo-sinensis), larch (Larix mastersiana) and cypress (Sabina saltuaria) foliar litter were placed on the forest floor beneath snowpack created by forest gaps in the eastern Tibet Plateau. The litterbags were sampled at the onset of freezing, deep freezing, thawing and growing stages from October 2010 to October 2012. Mass loss and N concentrations in litter were measured. Over two years of decomposition, N release occurred mainly during the first year, especially during the first winter. Litter N release rates (both in the first year and during the entire two-year decomposition study period) were higher in the center of canopy gaps than under closed canopy, regardless of species. Litter N release rates in winter were also highest in the center of canopy gaps and lowest under closed canopy, regardless of species, however the reverse was found during the growing season. Compared with broadleaf litter, needle litter N release comparisons of gap center to closed canopy showed much stronger responses to the changes in snow cover in winter and availability of sunshine during the growing season. As the decomposition proceeded, decomposing litter quality, microbial biomass and environmental temperature were important factors related to litter N release rate. This suggests that if winter warm with climate change, reduced snow cover in winter might slow down litter N release in alpine forest.
DOI:10.3321/j.issn:1000-0933.1999.02.002URL [本文引用: 1]
通过对海南岛霸王岭自然保护区原始热带山地雨林中林隙和非林隙林 分的调查,分析了海南岛热带山地雨林中主要树种在林隙内外的数量特征、树种对林隙的更新反应和树种多样性的变化规律.结果表明:(1)不同树种在林隙内和 非林隙林分中出现的频度、密度和显著度不同,从而表现出其重要值的不同.根据不同树种在林隙内外重要值位序的差异大小,将海南热带山地雨林中树种划分为7 种类型:即①只出现于林隙中,②只出现于非林隙林分中,③对林隙更新反应不显著,④对林隙有强烈正更新反应,⑤对林隙有强烈负更新反应,⑥对林隙有中等正 更新反应和⑦对林隙有中等负更新反应的树种.(2)海南热带山地雨林中主要树种的更新密度随林隙年龄阶段的变化曲线有单峰型和双峰型两大类,而单峰型中又 包括了前期高峰型、中期高峰型和后期高峰型3种亚类型.(3)林隙的存在增加了海南热带山地雨林的物种丰富度,提高了其多样性.(4)随林隙不同年龄阶段 的变化,树种多样性的变化趋势基本上为单峰型,在10~30a期间的多样性最大.
DOI:10.3321/j.issn:1000-0933.1999.02.002URL [本文引用: 1]
通过对海南岛霸王岭自然保护区原始热带山地雨林中林隙和非林隙林 分的调查,分析了海南岛热带山地雨林中主要树种在林隙内外的数量特征、树种对林隙的更新反应和树种多样性的变化规律.结果表明:(1)不同树种在林隙内和 非林隙林分中出现的频度、密度和显著度不同,从而表现出其重要值的不同.根据不同树种在林隙内外重要值位序的差异大小,将海南热带山地雨林中树种划分为7 种类型:即①只出现于林隙中,②只出现于非林隙林分中,③对林隙更新反应不显著,④对林隙有强烈正更新反应,⑤对林隙有强烈负更新反应,⑥对林隙有中等正 更新反应和⑦对林隙有中等负更新反应的树种.(2)海南热带山地雨林中主要树种的更新密度随林隙年龄阶段的变化曲线有单峰型和双峰型两大类,而单峰型中又 包括了前期高峰型、中期高峰型和后期高峰型3种亚类型.(3)林隙的存在增加了海南热带山地雨林的物种丰富度,提高了其多样性.(4)随林隙不同年龄阶段 的变化,树种多样性的变化趋势基本上为单峰型,在10~30a期间的多样性最大.
DOI:10.2307/1941693URL [本文引用: 4]
We examined the effects of gap size on rates of mass loss of leaf, branch and bark litter, and microbial activity in a Castanopsis kawakamii forest over a 360-d period. Fiver forest gap size classes that compromise a gap size gradient were chosen: (1) closed canopy of pure Castanopsis kawakamii vegetation; (2) small gaps with a diameter of < 5 m; (3) small-to-intermediate gaps with a diameter of 5-15 m; (4) intermediate-to-large gaps with a diameter of 15-30 m; (5) large gaps with a diameter of > 30 m. After 360 d, plant litter decomposing under closed canopy or within the small gaps lost mass more rapidly than those in large gaps. Among the litter quality parameters measured, initial N concentrations were most strongly positively correlated with the annual decay rates, and among the microclimatic factors, soil moisture content was the best predictor. Total substrate-induced respiration (SIR) was highest under closed canopy, intermediate in small gaps, and lowest in large gap size classes. Within gap size classes, total microbial respiration was highest for leaves, intermediate for bark, and lowest for branches. Across all sample dates, soil moisture content was the best predictor of microbial activity among the measured microclimatic factors. Plant litter C/N ratios were best predictors of microbial activity among the measured litter quality indices. Decomposition rate constants were linearly correlated with overall mean values of microbial activities for all three types of decomposing litter within five different gap size classes (R^2 = 0.984). The SIR rates of decomposing plant litter as a measure of potential active microbial biomass reflected the microclimate and litter quality. Our results suggest that large gaps significantly reduce microbial activity and decomposition rates by changing environmental conditions that consequently should reduce nutrient cycling rates in this system.
DOI:10.1016/j.agrformet.2015.06.015URL [本文引用: 2]
Gap size is critically important to ecological processes that drive forest dynamics within the gap, yet its threshold has never been explicitly defined. Consequently, gap sizes reported in the literature ranged from 4m2 to 2ha, which makes comparisons among and synthesis of the published gap studies difficult. We suggested that the lower size limit be defined by the mean shadow length (SL) of canopy trees surrounding the gap (CTSG) at local 12:00 during growing season (GS), while the upper size limit be defined by considering the farthest impact of CTSG on growth of shade intolerant tree species, which was determined by the mean of SL at the initial and the final times when 30-min photosynthetic active radiation (PAR) is more than the light saturation point for shade intolerant tree species each day during GS. The lower and upper limits of expanded gaps (the canopy gap plus the area extending to the bases of the canopy trees surrounding the gap) represented by gap diameter: CTSG height (RD/H) were 0.49 and 3.49, respectively, for temperate forest areas. The lower limit of gap size is determined only by the location and the height of CTSG, which should be applicable worldwide. We also tried to provide a universal method for determining the upper limit of gap size without applying the observed PAR data, and using only sunshine duration, an easily obtained variable from meteorological stations worldwide. We suggest that expanded gaps may be classified as: small gap, 0.49<RD/H 1.0, medium gap, 1.0<RD/H 2.0; large gap, 2.0<RD/H<3.5 in temperate forests.
Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates
1
2008
... id="C4">植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(
Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystem: A triangular relationship
3
1997
... id="C4">植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
The freezer defrosting: Global warming and litter decomposition rates in cold biomes
4
2006
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
... ), 且枝条冬季总质量损失比例明显高于生长季节.这主要是由于亚高山和高山森林季节性雪被和冻融循环特征及耐寒生物种群活动明显促进了寒冷季节的凋落物分解(
Influence of gap size and soil properties on microbial biomass in a subtropical humid forest of north-east India
2000
Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient
4
2010
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C24">质量损失率和质量损失速率(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... ); (2)雪被的绝热保温作用维持耐寒生物种群活动和生物活性同样能提高冬季凋落叶和枝条降解率(
Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. 3rd edn
3
2014
... id="C4">植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(
... ).已有的研究表明, 凋落叶约占森林生态系统凋落物年归还总量的70%, 凋落枝约占26% (
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Branch wood decomposition of tree species in a deciduous temperate forest in Korea
3
2017
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... ), 且显著高于温带落叶森林(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
大兴安岭低质林生态改造后枯落物水文效应变化
1
2015
... id="C17">将采集的样品分为两部分.一部分(4袋)用于枝条持水能力测定, 具体方法为: 将烘干至恒质量的凋落物称量, 然后装入凋落袋中, 浸入清水中24 h, 水面高度以浸过凋落袋, 凋落袋不露出水面为度, 将内装凋落物的凋落袋从清水中取出并悬挂在空中静置约5 min, 当无水滴滴下时立刻称量, 最后将袋中的凋落物烘干(65 ℃)称量, 计算最大持水率(
大兴安岭低质林生态改造后枯落物水文效应变化
1
2015
... id="C17">将采集的样品分为两部分.一部分(4袋)用于枝条持水能力测定, 具体方法为: 将烘干至恒质量的凋落物称量, 然后装入凋落袋中, 浸入清水中24 h, 水面高度以浸过凋落袋, 凋落袋不露出水面为度, 将内装凋落物的凋落袋从清水中取出并悬挂在空中静置约5 min, 当无水滴滴下时立刻称量, 最后将袋中的凋落物烘干(65 ℃)称量, 计算最大持水率(
Winter climate implications for decomposition in northeastern forests, comparisons change of sugar maple litter with herbivore fecal
2
2010
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
Tree fall gap size effects on above- and below-ground processes in a tropical wet forest
1
1998
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
Monthly changes of collembolan population under the gradients of moisture, organic carbon and nitrogen contents in a sub-tropical forest soil
2
2012
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C43">凋落物的分解是一个整体而连续的过程, 林窗对凋落物分解的影响常常随林窗面积大小和方位而发生变化, 这导致迄今为止的研究结果仍具有很大的不确定性(
Seasonal dynamics of litter fall in a sub-alpine spruce-fir forest on the eastern Tibetan Plateau: Allometric scaling relationships based on one year of observations
1
2017
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
Dynamics of alpine plant litter decomposition in a changing climate
2
2010
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Nutrient release from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire
1
1973
... id="C4">植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(
林窗生态学研究进展
2
2016
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
林窗生态学研究进展
2
2016
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest
3
2016
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
2
2013
... id="C27">凋落物质量损失4年时间各阶段所占比例(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
雪被斑块对高山森林两种灌木凋落叶质量损失的影响
2
2013
... id="C27">凋落物质量损失4年时间各阶段所占比例(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
The effects of forest gaps on cellulose degradation in the foliar litter of two shrub species in an alpine fir forest
2
2015
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
Temperature and plant species control over litter decomposition in Alaskan tundra
1
1996
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Winter regulation of tundra litter carbon and nitrogen dynamics
1996
Responses of springtail and mite populations to prolonged periods of soil freeze-thaw cycles in a sub-Arctic ecosystem
1
2007
... id="C18">在温度监测的任何时段, 将凋落袋下方地表温度低于0 ℃持续至少3 h, 随后回升到0 ℃至少持续3 h (或高于0 ℃持续至少3 h, 随后跌至0 ℃以下持续至少3 h)的温度动态过程记作一次冻融循环(
滇西北亚高山针叶林林窗大小与更新的初步分析
2
2002
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... ), 森林群落的自然更新以林窗更新为主(
滇西北亚高山针叶林林窗大小与更新的初步分析
2
2002
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... ), 森林群落的自然更新以林窗更新为主(
Gap dynamics of forest trees: A review with particular attention to boreal forests
2
2001
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... ), 可能通过两方面的机制影响高山森林凋落物的分解过程: (1)形成不同于林下的微环境条件及土壤生物群落结构(
Energy storage and the balance of producers and decomposers in ecological systems
2
1963
... id="C20">凋落物分解指数模型为: y = ae-kt
式中y为凋落物的月残留率(%), a为拟合参数, e为自然对数底, t为分解时间(月), k为凋落物分解系数(g·g-1) (
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Global-scale similarities in nitrogen release patterns during long-term decomposition
1
2007
... id="C4">植物凋落物分解是维持森林生态系统碳和养分循环的重要生态学过程, 受到生物与非生物因素的综合影响(
Effects of clear-cutting on decomposition rates of litter and forest floor in forests of British Columbia
1
2000
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
Effect of gap size on litter decomposition and soil concentrations in a high- elevation spruce fir forest
1
2003
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
Effects of gap-size classes on long-term litter decomposition rates of beech, oak and chestnut species at high elevations in Northeast Turkey
2
2008
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
... ;
Methods for studying treefall gaps: A review
1
2011
... id="C8">在高山森林生态系统定位研究站建立的1 hm2岷江冷杉原始林样地(坡向NE 38°, 坡度 24°)进行分解实验, 海拔3 598 m.林窗设置均指扩展林窗, 其边缘以边界木树干基部为准(
The decomposition rate of non-stem components of coarse woody debris (CWD) in European boreal forests mainly depends on site moisture and tree species
3
2016
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... ;
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Forest effects on snow accumulation and ablation: An integrative review of empirical results
2
2010
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
Litter decomposition in two subalpine forests during the freeze-thaw season
5
2010
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
... ).Hobbie和Chapin (1996)研究发现, 寒冷地区凋落物分解主要发生在第1年, 第2年以后以冬季分解为主.而川西亚高山和高山区域岷江冷杉新鲜凋落叶第1年质量损失的53%-64%出现在冬季(
... ;
高山森林林窗对凋落叶分解的影响
7
2016
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C7">研究区域位于四川省阿坝藏族羌族自治州理县的四川农业大学高山森林生态系统定位研究站(102.88°-102.95° E, 31.23°-31.32° N, 海拔2 458-4 619 m).研究站坐落于米亚罗自然保护区, 地处青藏高原东缘与四川盆地的过渡带, 区域内年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1 ℃, 年降水量约850 mm, 季节性冻融期长达5-6个月(
... id="C22">50%分解时间= -ln(1-0.50)/k (
... id="C24">质量损失率和质量损失速率(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
... ;
高山森林林窗对凋落叶分解的影响
7
2016
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C7">研究区域位于四川省阿坝藏族羌族自治州理县的四川农业大学高山森林生态系统定位研究站(102.88°-102.95° E, 31.23°-31.32° N, 海拔2 458-4 619 m).研究站坐落于米亚罗自然保护区, 地处青藏高原东缘与四川盆地的过渡带, 区域内年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1 ℃, 年降水量约850 mm, 季节性冻融期长达5-6个月(
... id="C22">50%分解时间= -ln(1-0.50)/k (
... id="C24">质量损失率和质量损失速率(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
... ;
川西高山森林林隙特征及干扰状况
4
2013
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C7">研究区域位于四川省阿坝藏族羌族自治州理县的四川农业大学高山森林生态系统定位研究站(102.88°-102.95° E, 31.23°-31.32° N, 海拔2 458-4 619 m).研究站坐落于米亚罗自然保护区, 地处青藏高原东缘与四川盆地的过渡带, 区域内年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1 ℃, 年降水量约850 mm, 季节性冻融期长达5-6个月(
... id="C8">在高山森林生态系统定位研究站建立的1 hm2岷江冷杉原始林样地(坡向NE 38°, 坡度 24°)进行分解实验, 海拔3 598 m.林窗设置均指扩展林窗, 其边缘以边界木树干基部为准(
川西高山森林林隙特征及干扰状况
4
2013
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
... id="C7">研究区域位于四川省阿坝藏族羌族自治州理县的四川农业大学高山森林生态系统定位研究站(102.88°-102.95° E, 31.23°-31.32° N, 海拔2 458-4 619 m).研究站坐落于米亚罗自然保护区, 地处青藏高原东缘与四川盆地的过渡带, 区域内年平均气温2-4 ℃, 最高气温23.7 ℃, 最低气温-18.1 ℃, 年降水量约850 mm, 季节性冻融期长达5-6个月(
... id="C8">在高山森林生态系统定位研究站建立的1 hm2岷江冷杉原始林样地(坡向NE 38°, 坡度 24°)进行分解实验, 海拔3 598 m.林窗设置均指扩展林窗, 其边缘以边界木树干基部为准(
Foliar litter nitrogen dynamics as affected by forest gap in the alpine forest of Eastern, Tibet Plateau
2
2014
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C6">位于青藏高原东缘的川西亚高山森林在调节区域气候、涵养水源、保育生物多样性等方面具有十分重要的作用(
海南霸王岭热带山地雨林林隙更新规律的研究
1
1999
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
海南霸王岭热带山地雨林林隙更新规律的研究
1
1999
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
Erects of gap size on litter decomposition and microbial activity in a subtropical forest
4
1995
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C40">林窗不仅是森林生态系统长期维持和循环更新的重要驱动力(
... id="C41">目前, 已有的林窗分解实验大多基于凋落叶、细根、粗母质残体(粗枝和倒木)开展, 较少涉及凋落枝(
... id="C42">川西亚高山森林位于青藏高原东缘, 冬季具有明显的季节性雪被和土壤冻融过程(
On the size of forest gaps: Can their lower and upper limits be objectively defined?
2
2015
... id="C5">林窗干扰是高山森林生态系统物质循环和群落更新的重要形式(
... id="C43">凋落物的分解是一个整体而连续的过程, 林窗对凋落物分解的影响常常随林窗面积大小和方位而发生变化, 这导致迄今为止的研究结果仍具有很大的不确定性(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}