Empirical relationship between specific leaf area and thermal dissipation of Phragmites australis in salt marshes of Qinwangchuan
LIQun, ZHAOCheng-Zhang*,, ZHAOLian-Chun, WANGJian-Liang, ZHANGWei-Tao, YAOWen-Xiu College of Geography and Environmental Science, Northwest Normal University, Research Center of Wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China 通讯作者:* 通信作者Author for correspondence (E-mail: zhaocz601@163.com) 收稿日期:2017-01-7 修回日期:2017-07-9 网络出版日期:2017-09-30 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41461013和91125014)
展开
摘要 植物比叶面积(SLA)与叶片热耗散的关联性分析, 对于探究植物光合产物积累与耗散及其适应特殊生境有重要意义。该研究以秦王川国家湿地公园芦苇(Phragmites australis)种群为研究对象, 根据距离水域的远近分别依次设置3个试验样地: I (离水域25-40 m, 土壤含水量(SM) 28.94%)、II (离水域10-25 m, SM 45.97%)、III (离水域0-10 m, SM 76.81%), 研究了秦王川盐沼湿地芦苇SLA与叶片热耗散的关系。结果表明: 随着土壤含水量的增加, 湿地植被群落的高度和地上生物量呈现逐渐增加的趋势, 土壤电导率和光合有效辐射呈显著减小的趋势; 叶面积、净光合速率和蒸腾速率均呈增加的趋势, 叶厚度呈减少的趋势, SLA呈现先增加后减小的趋势, 叶干质量呈现先减少后增加的趋势; 从样地I到样地III, 调节性能量耗散的量子产额(Y(NPQ))、非光化学淬灭系数呈减少的趋势, 实际光合效率、光化学淬灭系数呈增加的趋势, 非调节性能量耗散的量子产额呈先增加后减少的趋势; 在样地I和III, 芦苇SLA与Y(NPQ)呈极显著负相关关系(p < 0.01), 在样地II, SLA与Y(NPQ)显著负相关(p < 0.05)。随土壤含水量的梯度性变化, 芦苇种群通过调整叶片构建模式从而适时调整SLA的大小以改变叶片热耗散, 实现对植物吸收光能的有效利用, 反映了植物为适应特殊生境通过改变表型调控叶片热耗散实现自我保护的机制。
关键词:比叶面积;热耗散;调节性能量耗散的量子产额;芦苇;秦王川;盐沼湿地 Abstract Aims The correlation between specific leaf area (SLA) and thermal dissipation reflects not only the accumulation and dissipation of plant photosynthesis, but also plants’ adaptation to their habitats and changing environment. The objective of this study is to examine the correlation between SLA and thermal dissipation of reed (Phragmites australis) under different soil moisture conditions and salt contents.Methods Our study site was located in the National Wetland Park in Qinwangchuan, Gansu Province, China. Our sampling site extends from the edge to the central of a salt marsh where the reed was the single dominant species. The study site was divided into three zones based on the distance from the water. Within each zone, six 2 m × 2 m sampling plots were randomly located to select six reed individuals in each plot (total = 18). Vegetation height, aboveground biomass, soil moisture, and soil electrical conductivity (EC) were measured, with the six reed individuals taken to the laboratory to measure leaf thickness. Leaf net photosynthetic rate (Pn), transpiration rate (Tr), and other parameters of the reeds were also measured in each plot prior to harvesting. Quantitative measures of chlorophyll fluorescence were taken after 30-min dark adaptation. Quadrat survey method was used to model the empirical relationship between the transpiration rate and leaf characteristics.Important findings Vegetation height and aboveground biomass increased with soil moisture content, but EC and photosynthetically active radiation decreased. Leaf area, Tr and Pn increased along the gradient, leaf thickness showed decreasing, but the increasing trend of SLA switched to a decreasing trend, while leaf dry mass presented an opposite trend. From plot I to III, the quantum yield of regulated energy dissipation (Y(NPQ)) and non-photochemical quenching decreased, the actual photochemical efficiency of PSII and photochemical quenching increased, and quantum yield of non-regulated energy dissipation increased before decreasing. There appeared a highly significant negative correlation (p < 0.01) between SLA and Y(NPQ) at plot I and III, and a less significant negative correlation (p < 0.05) at plot II. Along the soil moisture gradient, reed seemed using light effectively by changing leaf thermal dissipation through adjusting their leaf size and SLA—A potential self-protection mechanism in light of adapting the habitat.
Keywords:specific leaf area;thermal dissipation;the quantum yield of regulated energy dissipation;Phragmites australis; Qinwangchuan;salt marshes -->0 PDF (1069KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 李群, 赵成章, 赵连春, 王建良, 张伟涛, 姚文秀. 秦王川盐沼湿地芦苇比叶面积与叶片热耗散的关联性分析. 植物生态学报, 2017, 41(9): 985-994 https://doi.org/10.17521/cjpe.2017.0005 LIQun, ZHAOCheng-Zhang, ZHAOLian-Chun, WANGJian-Liang, ZHANGWei-Tao, YAOWen-Xiu. Empirical relationship between specific leaf area and thermal dissipation of Phragmites australis in salt marshes of Qinwangchuan. Chinese Journal of Plant Ecology, 2017, 41(9): 985-994 https://doi.org/10.17521/cjpe.2017.0005 植物光合作用是驱动CO2在大气与生物圈之间交换的一个重要过程(Amoros-Lopez et al., 2008), 叶片的叶绿素荧光与光合作用密切相关, 在植物的光合作用、逆境生理和组织衰老等生理生态学研究方面具有重要作用(朱启红等, 2013)。光合作用通过光系统I (PSI)和光系统II (PSII)将光能转化为化学能, 其中PSII被认为对盐分胁迫尤其敏感, 在植物响应水盐胁迫的过程中发挥着重要作用(Baker, 1991), PSII调节性能量耗散的量子产额Y(NPQ)是PSII反应中心吸收的光量子利用方式之一(Kramer et al., 2004; 向芬等, 2014), 非光化学淬灭系数(NPQ)是反映植物耗散过剩光能转换为热能的指标, 可衡量植物的光破坏防御能力(张超等, 2014); 叶片作为植物体暴露在环境中表面积最大的器官, 对各种胁迫因子具有较强的感知能力, 而且能够通过叶面积、叶厚度等功能性状在形态和结构上的可塑性(Westoby & Wright, 2003; 施宇等, 2012), 差异化地调控叶片的比叶面积(SLA)、净光合速率、蒸腾作用等生理生态功能, 间接影响植物的光合能力和营养物质的吸收水平(Deng et al., 2008); SLA与叶片热耗散的关系十分密切, SLA较低的叶片将干物质多用于构建保卫结构, 叶肉细胞密度和叶片厚度高, 形成小而厚的叶片, 能够容纳更多的叶绿体, 一般具有高的Y(NPQ), 叶片的可调控生理代谢能力强, 如以热耗散等形式耗散过剩的光能(胡文海等, 2015), 因而叶片热交换和自我保护能力比较强, 有助于防止叶片过度失水和提高叶片的生境适应能力(胡耀升等, 2015), 这表明植物通常会通过调整叶片SLA来提高自身适应环境的能力。土壤水分是决定植物群落结构的一个主要环境因子(Rozema, 1996), 通过影响群落内部环境使种群内每个植株可获得性资源数量发生变化, 从而迫使植株进行表型调整, 以提高生境适合度(黎磊等, 2011); 在土壤水分差异条件下, 植物需要适时调整叶面积、SLA等叶片构型特征, 提高植物对环境空间异质性的适应能力, 借助叶片热耗散实现植物对叶片吸收的光能的积累与耗散的有效控制, 这体现了SLA与PSII热耗散机制存在着密切的关联性。因此, 研究SLA与PSII的热耗散机制, 对于探索特殊生境下的湿地植物表型和自我保护机制有着重要意义。 芦苇(Phragmites australis)属于多年生根茎型禾本科植物, 具有极强的环境适应能力, 广泛分布于内陆草甸区和近海河口湿地区, 也是西北荒漠区沼泽湿地生态系统的优势植物。目前, 学术界围绕植物代谢效率和群落密度的权衡(Deng et al., 2008)、植物叶片光合参数与环境因子的关系(Knight & Ackerly, 2003)、强光下叶绿素荧光的淬灭机理(Komura et al., 2010)、光呼吸和PSII热耗散对土壤水分的响应(张超等, 2014), 以及湿地植物叶片功能性状等问题展开了广泛研究(党晶晶等, 2015), 有关芦苇的生境胁迫适应机制(庄瑶等, 2010)、光合作用对土壤水盐因子的响应(戚志伟等, 2016)、多时空尺度蒸散模拟研究(于文颖等, 2014)等也受到了重视, 但是有关盐沼湿地植物比叶面积与叶片热耗散关联性尚不明晰。鉴于此, 本研究以秦王川国家湿地公园盐沼湿地的芦苇为研究对象, 通过分析盐沼湿地芦苇SLA与叶片热耗散的关联性, 试图明晰: (1)盐沼湿地芦苇SLA与叶片热耗散存在何种关联性?(2)形成这种关联性的主要原因有哪些?旨在丰富对湿地植物叶性状建成的种群适应性策略的认识和理解。
1 研究地区和研究方法
1.1 研究区域和样地概况
研究区位于甘肃省兰州市的秦王川盆地, 地理坐标为36.47° N, 103.65° E, 海拔1 895-1 897 m, 属大陆性季风气候, 年平均气温6.9 ℃, 年降水量 220 mm, 主要集中在6-9月, 年日照时间2β700 h, 平均无霜期126天。由于地处乌鞘岭褶皱山岭南侧的边缘低山区, 四周山地环绕, 中心地带地势平坦开阔, 形成盆地, 盆地南部广泛分布第四系松散层孔隙潜水, 含水层为砂碎石及中细砂层, 受常年或季节性水淹的影响, 形成以淡灰钙土为主的自然土壤, 发育了水生和湿生植物群落, 以湿生和挺水植物为主。主要植物有: 芦苇、枸杞(Lycium chinense)、多枝柽柳(Tamarix ramosissima)、水烛(Typha angustifolia)、灰绿藜(Chenopodium glaucum)、紫菀(Aster tataricus)、假苇拂子茅(Calamagrostis pseudophragmites)、碱蓬(Suaeda salsa)、苦苣菜(Sonchus oleraceus)、盐角草(Salicornia europaea)、早熟禾(Poa annua)等。
不同样地湿地群落的生物学特征和土壤理化性质的变化如表1所示。土壤含水量、光合有效辐射(PAR)和土壤电导率在3个观测样地中存在显著差异(p < 0.05, 表1), 从样地I到样地III土壤含水量增加了1.65倍, PAR减少了52.52%, 土壤电导率减少了73.36%; 随着土壤含水量的逐步增加和土壤电导率的逐渐减小, 芦苇高度和地上生物量随之增加, 从样地I到样地III分别增加了1.32倍和1.94倍, 且地上生物量和高度增加十分显著。表明样地I的土壤盐碱化程度最高, 群落内的光照强烈, 样地II的光照环境、土壤盐分较高、土壤含水量居中, 环境胁迫有所减弱, 而在样地III土壤盐分含量最低, 土壤含水量充足, 不存在水盐双重胁迫。表明在3个样地中, 样地III的生境条件更适宜芦苇种群的生长。 Table 1 表1 表1不同样地的湿地群落生物学特征和土壤特性(平均值±标准误差) Table 1Physiological and soil characteristics of wetland community in three sampling plots (mean ± SE)
样地 Plot
土壤含水量 Soil moisture content (%)
土壤电导率 Soil electrical conductivity (ms·cm-1)
PAR (μmol·m-2·s-1)
高度 Height (cm)
地上生物量 Aboveground biomass (g·m-2)
I
28.94 ± 1.45c
2.44 ± 0.12a
1 236.3 ± 3.78c
141.80 ± 7.09c
1 088.12 ± 54.40c
II
45.97 ± 2.30b
1.85 ± 0.09b
866.0 ± 4.43b
191.60 ± 9.58b
1 759.36 ± 87.97b
III
76.81 ± 3.84a
0.65 ± 0.03c
587.0 ± 4.99a
328.80 ± 16.64a
3 195.32 ± 159.77a
PAR, photosynthetically active radiation. Different lowercase letters in the same column indicate significant differences among plots (p < 0.05).PAR, 光合有效辐射。同列不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.2 芦苇叶性状特征和光合特性分析
如表2所示, 随着土壤水分含量的增加, 芦苇的叶面积(LA)、净光合速率(Pn)和蒸腾速率(Tr)均呈增加趋势, 从样地I到样地III分别增加了2.73、1.17和0.17倍; 芦苇的叶厚度呈减小趋势, 从样地I到样地III分别减少了25%; 比叶面积(SLA)呈先增加后减少的趋势, 总体增加了1.02倍; 叶干质量呈现先减小后增加的趋势, 总体增加了0.85倍。表明芦苇叶片形状和光合特性在3个样地变化较显著, 且叶性状具有较高的表型可塑性, 可以较好地适应特殊生境条件。 Table 2 表2 表2不同样地芦苇叶性状与光合生理参数特征(平均值±标准误差) Table 2Leaf characteristics and physiological measures of Phragmites australis in different plots (mean ± SE)
样地 Plot
I
II
III
叶面积 Leaf area (cm2)
7.71 ± 0.36c
14.19 ± 0.71b
28.75 ± 1.44a
叶厚度 Leaf thickness (mm)
0.36 ± 0.02a
0.32 ± 0.02b
0.27 ± 0.01c
叶干质量 Leaf dry mass (g)
0.39 ± 0.02b
0.22 ± 0.01c
0.72 ± 0.04a
比叶面积 Specific leaf area (cm2·g-1)
19.77 ± 0.99c
64.50 ± 3.23a
39.93 ± 2.00b
Pn (μmol CO2·m-2 ·s-1)
3.39 ± 0.17c
4.41 ± 0.22b
7.34 ± 0.37a
Tr (mmol H2O·m-2·s-1)
1.00 ± 0.05b
1.04 ± 0.05b
1.17 ± 0.06a
Pn, net photosynthetic rate; Tr, transpiration rate. Lowercase letters in the same line indicate significant differences among plots (p < 0.05).Pn, 净光合速率; Tr, 蒸腾速率。同行不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.3 不同土壤水分条件下芦苇叶片叶绿素荧光参数分析
如表3所示, 随着土壤水分含量的增加, 实际光合效率(Y(II))和光化学淬灭系数(QP)均呈增加趋势, 从样地I到样地III分别增加了0.71和0.55倍, 非光化学淬灭系数(NPQ)和调节性能量耗散的量子产额(Y(NPQ))均呈减少趋势, 从样地I到样地III分别减少了24%和28%, 非调节性能量耗散的量子产额(Y(NO))呈先增加后减少的趋势, 总体减少了3.6%。表明芦苇为适应特殊生境, 适时调整PSII天线色素吸收光能的分配, 以保护自身正常生长和繁殖。 Table 3 表3 表3不同样地芦苇叶片叶绿素荧光特征(平均值±标准误差) Table 3Leaf chlorophyll fluorescences of Phragmites australis in different plots (mean ± SE)
样地 Plot
Y(II)
QP
NPQ
Y(NO)
Y(NPQ)
I
0.21 ± 0.01c
0.42 ± 0.02b
0.45 ± 0.02a
0.29 ± 0.01a
0.50 ± 0.03a
II
0.25 ± 0.01b
0.60 ± 0.03a
0.43 ± 0.02a
0.30 ± 0.02a
0.45 ± 0.02b
III
0.36 ± 0.02a
0.65 ± 0.03a
0.34 ± 0.02b
0.28 ± 0.01a
0.36 ± 0.02c
Y(II), the actual photochemical efficiency of PSII; QP, photochemical quenching; NPQ, the quantum yield of regulated energy dissipation; Y(NO), the quantum yield of non-regulated energy dissipation; Y(NPQ), the quantum yield of regulated energy dissipation. Lowercase letters in the same column indicate significant differences among plots (p < 0.05).Y(II), 实际光合效率; QP, 光化学淬灭系数; NPQ, 非光化学猝灭系数; Y(NO), 非调节性能量耗散的量子产量; Y(NPQ), 调节性能量耗散的量子产额。同列不同小写字母表示样地间差异显著(p < 0.05)。 新窗口打开
2.4 芦苇SLA和叶片Y(NPQ)的关系分析
不同水分条件下芦苇比叶面积(SLA)与Y(NPQ)的关系存在显著差异(p < 0.05, 图1)。Pearson相关性分析表明, 芦苇叶片SLA与Y(NPQ)在样地I和III呈极显著负相关关系(yI = -1.949x + 0.427, R² = 0.813, p < 0.01, yIII = -2.967x - 0.032, R² = 0.834, p < 0.01, 图1), 在样地II, 二者呈显著负相关关系(yII = -1.232x + 1.536, R² = 0.679, p < 0.05, 图1)。表明随着土壤水分逐渐增加, 芦苇的SLA逐渐增加, Y(NPQ)逐渐减少, 二者在样地I和III呈极显著负相关关系(p < 0.01), 在样地II呈显著的负相关关系(p < 0.05), 样地II生境是芦苇植株的SLA增加和Y(NPQ)减小的过渡区域。 显示原图|下载原图ZIP|生成PPT 图1不同样地芦苇比叶面积(SLA)与调节性能量耗散的量子产额(Y(NPQ))之间关系。 -->Fig. 1Relationship between specific leaf area (SLA) and quantum yield of regulated energy dissipation (Y(NPQ)) of Phragmites australis at three sampling plots. -->
Amoros-LopezJ, Gomez-ChovaL, Vila-FrancesJ, AlonsoL, CalpeJ, MorenoJ, del Valle-TasconS (2008). Evaluation of remote sensing of vegetation fluorescence by the analysis of diurnal cycles. International Journal of Remote Sensing, 29, 5423-5436. [本文引用: 1]
[2]
BakerNR (1991). A possible role for photosystem II in environmental perturbations of photosynthesis. Physiologia Plantarum, 81, 563-570. [本文引用: 1]
[3]
DangJJ, ZhaoCZ, LiJ, HouZJ, DongXG (2015). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China. Chinese Journal of Plant Ecology, 39, 23-31. (in Chinese with English abstract)[党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚 (2015). 祁连山高寒草地甘肃臭草叶性状与坡向间的关系 . 植物生态学报, 39, 23-31.] [本文引用: 1]
[4]
Demmig-AdamsB, Adams WWIII (2006). Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytologist, 172, 11-21. [本文引用: 1]
[5]
DengJM, LiT, WangGX, LiuJ, YuZL, ZhaoCM, JiMF, ZhangQ (2008). Trade-offs between the metabolic rate and population density of plants. PLOS ONE, 3, e1799. doi: 10.1371/journal.pone.0001799. [本文引用: 2]
[6]
GivnishTJ (1978). Ecological aspects of plant morphology: Leaf form in relation to environment. Acta Biotheoretica, 27, 83-142. [本文引用: 1]
[7]
GongJR, HuangYM, GeZW, DuanQW, YouX, AnR, ZhangXS (2009). Ecological responses to soil water content in four hybrid Populous clones. Chinese Journal of Plant Ecology, 33, 387-396. (in Chinese with English abstract)[龚吉蕊, 黄永梅, 葛之葳, 段庆伟, 尤鑫, 安然, 张新时 (2009). 4种杂交杨对土壤水分变化的生态学响应 . 植物生态学报, 33, 387-396.] [本文引用: 1]
[8]
GongR, GaoQ (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions. Chinese Journal of Plant Ecology, 39, 300-308. (in Chinese with English abstract)[龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展 . 植物生态学报, 39, 300-308.] [本文引用: 1]
[9]
HanL, ZhaoCZ, XuT, FengW, DuanBB, ZhengHL (2016). Trade-off between leaf size and vein density of Achnatherum splendens in Zhangye wetland. Chinese Journal of Plant Ecology, 40, 788-797. (in Chinese with English abstract)[韩玲, 赵成章, 徐婷, 冯威, 段贝贝, 郑慧玲 (2016). 张掖湿地芨芨草叶大小和叶脉密度的权衡关系 . 植物生态学报, 40, 788-797.] [本文引用: 1]
[10]
HuWH, ZhangSS, XiaoYA, YanXH (2015). Physiological responses and photo-protective mechanisms of two Rhododendron plants to natural sunlight after long term shading. Chinese Journal of Plant Ecology, 39, 1093-1100. (in Chinese with English abstract)[胡文海, 张斯斯, 肖宜安, 闫小红 (2015). 两种杜鹃花属植物对长期遮阴后全光照环境的生理响应及其光保护机制 . 植物生态学报, 39, 1093-1100.] [本文引用: 1]
[11]
HuYS, MoXY, LiuYH (2015). Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains. Acta Ecologica Sinica, 35, 1480-1487. (in Chinese with English abstract)[胡耀升, 么旭阳, 刘艳红 (2015). 长白山森林不同演替阶段比叶面积及其影响因子 . 生态学报, 35, 1480-1487.] [本文引用: 2]
[12]
HultineKR, MarshallJD (2000). Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia, 123, 32-40. [本文引用: 1]
[13]
KangSZ, ZhangJH (2004). Controlled alternate partial root- zone irrigation: Its physiological consequences and impact on water use efficiency. Journal of Experimental Botany, 55, 2437-2446. [本文引用: 1]
[14]
KnightCA, AckerlyDD (2003). Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: Congeneric species from desert and coastal environments. New Phytologist, 160, 337-347. [本文引用: 1]
[15]
KomuraM, YamagishiA, ShibataY, IwasakiI, ItohS (2010). Mechanism of strong quenching of photosystem II chlorophyll fluorescence under drought stress in a lichen,Physciella melanchra, studied by subpicosecond fluorescence spectroscopy. Biochim Biophys Acta, 1797, 331-338. [本文引用: 2]
[16]
KramerDM, JohnsonG, KiiratsO, EdwardsGE (2004). New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynthesis Research, 79, 209-218. [本文引用: 1]
[17]
LiL, ZhouDW, ShengLX (2011). Density dependence-determined plant biomass allocation pattern. Chinese Journal of Ecology, 30, 1579-1589. (in Chinese with English abstract)[黎磊, 周道玮, 盛连喜 (2011). 密度制约决定的植物生物量分配格局 . 生态学杂志, 30, 1579-1589.] [本文引用: 1]
[18]
LiR, JiangZM, ZhangSX, CaiJ (2015). A review of new research progress on the vulnerability of xylem embolism of woody plants. Chinese Journal of Plant Ecology, 39, 838-848. (in Chinese with English abstract)[李荣, 姜在民, 张硕新, 蔡靖 (2015). 木本植物木质部栓塞脆弱性研究新进展 . 植物生态学报, 39, 838-848.] [本文引用: 1]
[19]
LiYL, CuiJY, SuYZ (2005). Specific leaf area and leaf dry matter content of some plants in different dune habitats. Acta Ecologica Sinica, 25, 304-311. (in Chinese with English abstract)[李玉霖, 崔建垣, 苏永中 (2005). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较 . 生态学报, 25, 304-311.]
[20]
LiuMH, XinZM, XuJ, SunF, DouLJ, liYH (2013). Influence of leaf size of plant on leaf transpiration and temperature in arid regions. Chinese Journal of Plant Ecology, 37, 436-442. (in Chinese with English abstract)[刘明虎, 辛智鸣, 徐军, 孙非, 窦立军, 李永华 (2013). 干旱区植物叶片大小对叶表面蒸腾及叶温的影响 . 植物生态学报, 37, 436-442.] [本文引用: 1]
[21]
LuoXG (2003). The discussion and analysis of Qinwangchuan irrigated area drainage and salinization prevention question. The Agriculture of Gansu, (12), 89. (in Chinese)[罗星光 (2003). 秦王川灌区排水和盐渍化防治问题的探讨与分析 . 甘肃农业, (12), 89.] [本文引用: 1]
[22]
OgburnRM, EdwardsEJ (2012). Quantifying succulence: A rapid, physiologically meaningful metric of plant water storage. Plant, Cell & Environment, 35, 1533-1542. [本文引用: 2]
[23]
PoorterL, BongersF (2006). Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology, 87, 1733-1743. [本文引用: 1]
[24]
QiZW, JiangN, GaoYN, LiY, WangKY, ZhangC (2016). Responses of photosynthesis of Phragmites australis to water-salt indicators of soil in Dongtan wetlands, Chongming Island. Wetland Island, 14, 538-545. (in Chinese with English abstract)[戚志伟, 姜楠, 高艳娜, 李艳, 王开运, 张超 (2016). 崇明岛东滩湿地芦苇光合作用对土壤水盐因子的响应 . 湿地科学, 14, 538-545.] [本文引用: 1]
[25]
RenQJ, LiHL, PuHY (2015). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chinese Journal of Plant Ecology, 39, 593-603. (in Chinese with English abstract)[任青吉, 李宏林, 卜海燕 (2015). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较 . 植物生态学报, 39, 593-603.] [本文引用: 1]
[26]
RozemaJ (1996). Biology of Halophytes: Halophytes and Biosaline Agriculture. Marcel Dekker , New York.17-30. [本文引用: 1]
[27]
SchreiberU (2004). Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. In: Papageorgiou GC, Govindjee eds. Chlorophyll a Fluorescence. Springer, Heidelberg, Germany. 279-319.
[28]
SchreiberU, BilgerW, NeubauerC (1994). Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze ED, Caldwell MM eds. Ecophysiology of Photosynthesis. Springer-Verlag, Berlin. 49-70. [本文引用: 1]
[29]
ScoffoniC, RawlsM, McKownA, CochardH, SackL (2011). Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture. Plant Physiology, 156, 832-843. [本文引用: 1]
[30]
ShiY, WenZM, GongSH, SongG, ZhenY, DingM (2012). Trait variations along a climatic gradient in hilly area of Loess Plateau. Research of Soil and Water Conservation, 19(1), 107-111, 116. (in Chinese with English abstract)[施宇, 温仲明, 龚时慧, 宋光, 郑颖, 丁曼 (2012). 黄土丘陵区植物功能性状沿气候梯度的变化规律 . 水土保持研究, 19(1), 107-111, 116.] [本文引用: 1]
[31]
SongLL, FanJW, WuSH, ZhongHP, WangN (2012). Response characteristics of leaf traits of common species along an altitudinal gradient in Hongchiba grassland, Chongqing. Acta Ecologica Sinica, 32, 2759-2767. (in Chinese with English abstract)[宋璐璐, 樊江文, 吴绍洪, 钟华平, 王宁 (2012). 红池坝草地常见物种叶片性状沿海拔梯度的响应特征 . 生态学报, 32, 2759-2767.] [本文引用: 1]
[32]
WestobyM, WrightIJ (2003). The leaf size-twig size spectrum and its relationship to other important spectra of variation among species. Oecologia, 135, 621-628. [本文引用: 1]
WrightIJ, WestobyM, ReichPB (2002). Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. Journal of Ecology, 90, 534-543. [本文引用: 1]
[35]
XiangF, ZhouQ, TianXR, ChenGX, XiaoY (2014). Leaf morphology and PSII chlorophyll fluorescence parameters in leaves of Sinosenecio jishouensis in different habitats. Acta Ecologica Sinica, 34, 337-344. (in Chinese with English abstract)[向芬, 周强, 田向荣, 陈功锡, 肖艳 (2014). 不同生境吉首蒲儿根叶片形态和叶绿素荧光特征的比较 . 生态学报, 34, 337-344.] [本文引用: 1]
[36]
YuHY, ChenYT, XuZZ, ZhouGS (2014). Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol. Chinese Journal of Plant Ecology, 38, 1029-1040. (in Chinese with English abstract)[于鸿莹, 陈莹婷, 许振柱, 周广胜 (2014). 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析 . 植物生态学报, 38, 1029-1040.] [本文引用: 1]
[37]
YuWY, JiRP, FengRui, WuJW, ZhaoXL, ZhangYS (2014). Advances in simulation of reed wetland evapotranspiration based on multiple time and space scales. Chinese Journal of Ecology, 33, 1388-1394. (in Chinese with English abstract)[于文颖, 纪瑞鹏, 冯锐, 武晋雯, 赵先丽, 张玉书 (2014). 芦苇湿地多时空尺度蒸散模拟研究进展 . 生态学杂志, 33, 1388-1394.] [本文引用: 1]
[38]
ZhangC, ZhanDX, ZhangPP, ZhangYL, LuoHH, ZhangWF (2014). Responses of photorespiration and thermal dissipation in PSII to soil water in cotton bracts. Chinese Journal of Plant Ecology, 38, 387-395. (in Chinese with English abstract)[张超, 占东霞, 张鹏鹏, 张亚黎, 罗宏海, 张旺锋 (2014). 棉花苞叶光呼吸和PSII热耗散对土壤水分的响应, 植物生态学报, 38, 387-395.] [本文引用: 2]
[39]
ZhangYQ, LiangCZ, WangW, WangLX, PengJT, YanJC, JiaJZ (2010). Soil salinity and Achnatherum splendens distribution. Chinese Journal of Ecology, 29, 2438-2443. (in Chinese with English abstract)[张雅琼, 梁存柱, 王炜, 王立新, 彭江涛, 闫建成, 贾成朕 (2010). 芨芨草群落土壤盐分特征 . 生态学杂志, 29, 2438-2443.] [本文引用: 1]
[40]
ZhuQH, XiaHX, XieHH, LiQ, DingWQ (2013). Response of Acorus tatarinowii in chlorophyll fluorescence to flooding stress. Oceanologia et Limnologia Sinica, 44, 1494-1499. (in Chinese with English abstract)[朱启红, 夏红霞, 谢海航, 李强, 丁武泉 (2013). 石菖蒲叶绿素荧光特性对淹水胁迫的响应 . 海洋与湖沼, 44, 1494-1499.] [本文引用: 1]
[41]
ZhuangY, SunYX, WangZS, YangLL, DengZF, YaoZG, AnSQ (2010). Research advances in ecotypes of Phragmites australis. Acta Ecologica Sinica, 30, 2173-2181. (in Chinese with English abstract)[庄瑶, 孙一香, 王中生, 杨琳璐, 邓自发, 姚志刚, 安树青 (2010). 芦苇生态型研究进展 . 生态学报, 30, 2173-2181.] [本文引用: 2]
Evaluation of remote sensing of vegetation fluorescence by the analysis of diurnal cycles. 1 2008
Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: Congeneric species from desert and coastal environments. 1 2003
Mechanism of strong quenching of photosystem II chlorophyll fluorescence under drought stress in a lichen,Physciella melanchra, studied by subpicosecond fluorescence spectroscopy. 2 2010
 , 赵连春, 王建良, 张伟涛, 姚文秀
, 赵连春, 王建良, 张伟涛, 姚文秀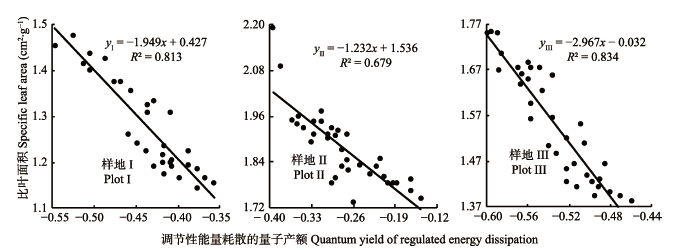 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT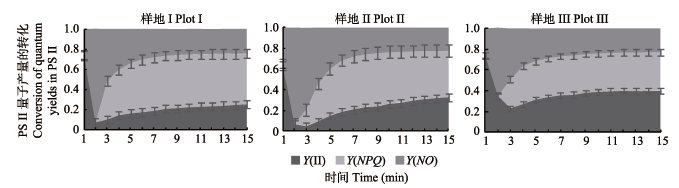 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}