
Effects of water condition on photochemical efficiency and physiological characteristics in artificially cultivated moss Syntrichia caninervis
XUHong-Mei通讯作者:
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (1270KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主。藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(Eldridge & Leys, 2003; 杨凯等, 2012)。荒漠藓类植物在极端环境中经过长期进化具备了一定的环境适应策略以避免不利环境条件的伤害, 如, 当环境缺水时荒漠藓类植株个体干燥收缩, 呈休眠状态, 遇水后又能迅速恢复生理活性, 表现出较强的耐旱特性(田桂泉等, 2005b); 同时, 在长期干旱少雨的沙漠环境下, 藓类植物具有较强的无性繁殖能力, 其配子体的任何部位都能再生形成新的植株(陈圆圆等, 2008)。藓类植物的这些功能与特性决定了其在沙漠修复和植被管理中具有重要的生态意义(Stark & McLetchie, 2006)。
自然条件下藓类结皮的形成需要几年甚至数十年的时间(Brotherson et al., 1983)。过度放牧等人类活动以及恶劣的自然环境对藓类结皮造成了极大的破坏, 进一步加速了沙漠化进程(Belnap, 1995)。因此, 藓类结皮的大规模人工培养将有助于荒漠生态系统的可持续发展, 可能是实现荒漠地区沙面固定和荒漠化防治的重要途径。目前关于藓类结皮的人工培养包括室内培养箱或温室大棚中培养(田桂泉, 2005a; Zhao et al., 2014), 以及野外环境下人工促进藓类结皮恢复(Lan et al., 2015), 两种方式均获得了一定成效, 培养出了盖度和密度较高的藓类结皮, 其假根具有捆绑沙粒的作用(许书军, 2007), 但培养后期藓株较野外藓株颜色浅, 人工培养藓株主要表现为嫩黄绿色, 而野外藓株主要呈现深绿色, 且人工培养藓株在生物量及株高方面都不及野外藓株(Xiao et al., 2011)。野外藓类结皮恢复耗时较长, 同时两种条件培养下的藓类结皮后期均出现衰退现象(田桂泉, 2005a)。如何改进藓类结皮的生长条件, 增强其后期的生存能力, 从而为人工培养荒漠藓类植物最终应用于野外的土壤生态恢复工作提供理论依据。干旱区水分是限制维管植物生长的最为关键的因子(Xu et al., 2007), 对于藓类结皮的生长恢复也起着至关重要的作用。野外人工藓类结皮层面积和盖度与水分条件显著相关, 会随着降雨量增加而拓展, 反之减小(贾艳等, 2012)。在野外藓结皮恢复研究中发现, 与营养基质和光照条件相比, 水分条件是首要的限制因子, 良好的水分条件或较高的空气湿度能够有效地促进藓类植物的定植生长, 产生更多原丝体(Duckett et al., 2004; Kidron et al., 2009)。苔藓植物本身对水分变化也十分敏感(Wu et al., 2015), 遇到干旱时, 在形态结构方面, 荒漠藓类可通过增加细胞壁厚度、调整叶片形态及疣密度、叶片芒尖长度等来减少水分散失(郑云普等, 2009; 陶冶和张元明, 2012), 在生理上能通过积累脯氨酸、可溶性糖等物质来提高细胞渗透压, 稳定细胞内大分子物质功能(张显强等, 2004), 以及合成大量的保护酶, 如超氧化物歧化酶(SOD)、过氧化物酶(POD)等, 防御过多的活性氧和其他过氧化物自由基对细胞膜系统的伤害(Wood et al., 2000; 张静和张元明, 2014), 从而保护植物细胞的活性, 因此我们推测水分变化会显著影响人工培育苔藓的光合生理活性, 从而影响植物的生长过程。
本研究选取古尔班通古特沙漠生物结皮优势藓类齿肋赤藓(Syntrichia caninervis)为实验对象, 研究3种不同的水分条件处理对室内人工培养齿肋赤藓光合活性、渗透调节物质(游离脯氨酸、可溶性蛋白、可溶性糖)含量和抗氧化酶活性等方面的影响, 研究人工培养藓类结皮后期不同水分条件下的生理特征, 确定人工培养齿肋赤藓培养后期管理的最佳水分处理条件, 为人工培养藓类结皮在野外成功定植提供一定的理论基础。
1 材料和方法
1.1 样品及培养基质的采集
2014年11月末, 选择古尔班通古特沙漠南缘典型丘间低地(44.67° N, 88.17° E), 采集长势良好的齿肋赤藓植株。采集前用装有蒸馏水的喷壶将结皮表面喷湿, 以确保采集样品的完整性。利用自制的圆形PVC管(内径6 cm, 高3.5 cm)随机采集远离灌丛的齿肋赤藓斑块10份, 并及时运回实验室自然风干, 同时将藓斑块中的植物残渣、土块、石子等剔除。在丘间低地无结皮发养区, 采集2-10 cm土壤层沙土, 带回实验室自然风干, 研磨过筛(2 mm)用作齿肋赤藓的沙质培养基质。1.2 试验设计
1.2.1 齿肋赤藓的人工培养实验采用茎叶碎片撒播法接种, 将齿肋赤藓植株地上茎叶部分统一用植物式样粉碎机(型号: FZ102)粉碎备用。称取60 g沙土, 均匀撒在培养皿内。将准备好的接种样品均匀撒于沙土表面(每皿干质量0.2 g), 在碎片表面再均匀覆一层2 mm厚的沙土; 用小型喷壶缓慢均匀地将少量蒸馏水(13 mL)喷洒在培养皿沙土表面, 用封口膜将培养皿密封后放入人工气候箱(PQX-400, 上海海向仪器设备厂)中培养。培养条件统一设置为: 温度15℃/10 ℃ (白天/夜晚), 光照强度12 000 lx, 光照周期14 h/10 h (光/暗)。
1.2.2 水分处理
沙漠水分来源主要依靠降水, 而降水的分配具有明显的季节差异, 如古尔班通古特沙漠降雨多集中在早春夏末, 早春融雪及夏季降雨后土壤水分短暂时间内处于一个过饱和的状态且面临一定的水分胁迫, 其他季节干旱。研究区降水主要以0-5 mm的小雨量为主, 占该沙漠总降雨频次的89.8%, 平均降雨间隔在6天左右(王亚婷和唐立松, 2009)。为研究不同水分处理对室内人工培养的齿肋赤藓光合生理及生化特性的影响, 本试验通过设置不同的水分施加频次来模拟水分胁迫的程度。实验中选取15个培养48天且其长势较一致的齿肋赤藓培养皿, 每天施加一定水量(模拟研究区自然条件春季融雪或降雨频繁季节时, 水分条件充足, 苔藓往往处于水分过剩的状态, 即为完全湿润处理)、每3天加水(中度湿润处理, 野外藓结皮培养3-4天加水藓结皮发育良好(Zhao et al., 2014))、每6天加水, 藓株遭遇干旱缺水, 因此本文以每6天加水一次作为干旱处理) 3种水分梯度处理, 加水时间统一为10:00, 每次水量为2 mm降雨(相当于13 mL), 降雨量大小设置参照我们前期研究, 在古尔班通古特沙漠, 小于2 mm的降雨为无效降雨, 当降雨量≥2 mm时苔藓植株表现为碳的净吸收(Wu et al., 2015)。同时, 考虑到人工苔藓的生长需要较长时间, 整个处理时间为48天。
模拟降雨量的计算公式为:
V蒸馏水=1/4× πd2 × h降雨量
式中: V蒸馏水为所添加的蒸馏水的体积(mL); d为培养皿的内径(cm); h降雨量为模拟降雨量(cm)。
1.2.3 培养齿肋赤藓光化学效率的测定
处理48天后, 利用便携式调制叶绿素荧光仪(PAM 2500, Walz, Heinz, Germany)进行人工培养齿肋赤藓叶绿素荧光特性的测定。测定前先对样品进行30 min充分暗适应后, 每个培养皿内随机选取3株藓株于波长650 nm、强度为0.1 μmol·m-2·s-1的红光下测得初始荧光(F0), 最大荧光(Fm)于强度为 10β000 μmol·m-2·s-1的饱和脉冲光下测得, 脉冲时间为0.8 s。在光照强度为250 μmol·m-2·s-1的光照下测定稳态荧光(Ft), 待Ft稳定后, 打开一次饱和脉冲光测得光适应后的最大荧光(Fm°), 样品重复数为5个。根据以上参数可以计算光系统II (PSII)的最大光化学效率(Fv/Fm) = (Fm - F0)/Fm和光适应下PSII的实际光化学效率(ΦPSII) = Fm° - Ft/Fm° (张静和张元明, 2011)。
1.2.4 取样
待藓株的光化学效率测定完毕, 先用镊子从每个培养皿中随机分离出一定数量的齿肋赤藓单株, 并用刀片小心地切除其假根, 并用自来水缓慢地冲洗干净, 避免夹杂沙粒的影响。每一样品称取0.25 g (鲜质量)用于植物组织含水量、光合色素含量、可溶性糖、游离脯氨酸和丙二醛含量的测定, 称取0.15 g样品(鲜质量)用于抗氧化酶含量的测定。
1.2.5 植物组织含水量测定
将称量过鲜质量的齿肋赤藓样品置于烘箱内, 105 ℃下杀青, 然后将烘箱温度降至80 ℃, 烘至恒质量后称取其干质量。
组织含水量(占干质量%) = (mF - mD)/mF ×100%
式中: mF为样品鲜质量(g); mD为样品干质量(g)。
1.2.6 光合色素含量的测定
光合色素含量的测定参照包维楷和冷俐(2005)的方法, 95%的乙醇提取比色测定法, 以95%的乙醇为空白对照, 在665、649、470 nm下测定样品的吸光度。各色素含量的计算公式如下:
叶绿素a含量: ρchl-a = 13.95A665 nm - 6.88A649 nm
叶绿素b含量: ρchl-b = 24.96A649 nm - 7.32A649 nm
类胡萝卜素含量: ρcaro = (1000 A470 nm - 2.05ρchl-a - 114.8ρchl-b)/245
色素含量= ρ × V/1000/m
式中: ρ为色素浓度(mg·L-1); V为提取液体积(L); m为样品干质量(g)。
1.2.7 渗透调节物质和抗氧化酶活性的测定
可溶性糖、可溶性蛋白和脯氨酸含量分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定(Gonzalez & Pignata, 1994; 尹本丰和张元明, 2015); 过氧化氢酶(CAT)、POD、 SOD活性和丙二醛(MDA)含量分别采用紫外分光光度法、愈创木酚法、羟胺法和硫代巴比妥酸(TBA)法进行测定(Sun et al., 2009; 尹本丰和张元明, 2015)。
1.3 数据处理
实验中每个水分条件处理设置5个重复, 实验结果均测定5次, 取平均值。各生理生化指标含量测定均以齿肋赤藓干质量表示。使用SPSS 19.0软件, 对所获得的不同水分条件处理下的人工培养齿肋赤藓植株的各项指标进行数据正态性检验(K-S检验)。对同一测试指标各水分条件处理下的数据进行单因素方差分析和多重比较(LSD), Origin 8.0软件作图。2 结果和分析
2.1 不同水分条件处理对人工培养齿肋赤藓植株含水量的影响
人工培养的齿肋赤藓植株含水量随加水频次的增加呈显著上升的趋势(图1)。每6天的加水处理下, 人工培养的齿肋赤藓植株平均含水量占藓株干质量的50%左右, 藓株处于干旱缺水状态; 每3天的加水处理下藓株平均含水量占藓株干质量的80%左右, 处于中等含水量状态; 而每天的加水处理下, 藓株平均含水量占藓株干质量的110%左右, 处于饱和含水量状态。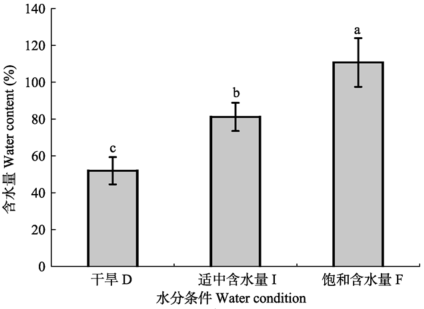 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1人工培养齿肋赤藓植株含水量对不同水分处理的响应(平均值±标准偏差)。不同字母表示差异显著(p < 0.05)。
-->Fig. 1Responses of water content in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD). D, drought treatment; I, intermediate water supply; F, fully watered. Different lowercase letters indicate significant differences (p < 0.05).
-->
2.2 不同水分条件处理对人工培养的齿肋赤藓光合色素含量的影响
在不同的水分条件处理下, 人工培养的齿肋赤藓的叶绿素a、叶绿素b、总叶绿素、类胡萝卜素含量以及叶绿素a/b的变化具有一定差异(表1)。随着加水频次的增加, 即水分含量的增加, 叶绿素a、叶绿素b、总叶绿素含量及叶绿素a/b呈现出先升高后降低的变化趋势, 在中度湿润处理下, 叶绿素a、叶绿素b、总叶绿素含量最高, 在完全湿润处理下, 三者含量较低, 但叶绿素a含量及叶绿素a/b仍高于干旱处理, 且叶绿素a/b在湿润处理下显著高于干旱处理(p < 0.05); 随着水分含量的增加, 类胡萝卜素含量逐渐降低。Table 1
表1
表1人工培养齿肋赤藓光合色素含量对不同水分处理的响应(平均值±标准偏差)
Table 1Responses of photosynthetic pigment content in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD)
| 处理 Treatment | 叶绿素a Chlorophyll a (mg·g-1 fresh mass) | 叶绿素b Chlorophyll b (mg·g-1 fresh mass) | 总叶绿素 Total chlorophyll (mg·g-1 fresh mass) | 类胡萝卜素 Carotenoid (mg·g-1 fresh mass) | 叶绿素a/b Chlorophyll a/b |
|---|---|---|---|---|---|
| 干旱处理 Drought treatment | 0.19 ± 0.02b | 0.37 ± 0.02a | 0.56 ± 0.03b | 0.03 ± 0.01a | 0.51 ± 0.18b |
| 中度湿润处理 Intermediate water supply treatment | 0.49 ± 0.03a | 0.42 ± 0.45a | 0.91 ± 0.03a | 0.02 ± 0.00b | 1.28 ± 0.25a |
| 完全湿润处理 Fully watered treatment | 0.21 ± 0.03b | 0.20 ± 0.05b | 0.41 ± 0.06c | 0.01 ± 0.00c | 1.25 ± 0.28a |
新窗口打开
2.3 不同水分条件处理对人工培养的齿肋赤藓PSII光化学效率的影响
由图2可知, 人工培养的齿肋赤藓PSII的光化学效率在不同的水分条件处理下发生了显著变化。在中度湿润处理时, Fv/Fm值最高(图2A), 显著高于干旱处理及完全湿润处理(p < 0.05), 随着水分含量的增加, Fv/Fm呈现出先升高后降低的趋势, 在完全湿润处理时, Fv/Fm值最低; ΦPSII的变化趋势和Fv/Fm一致(图2B)。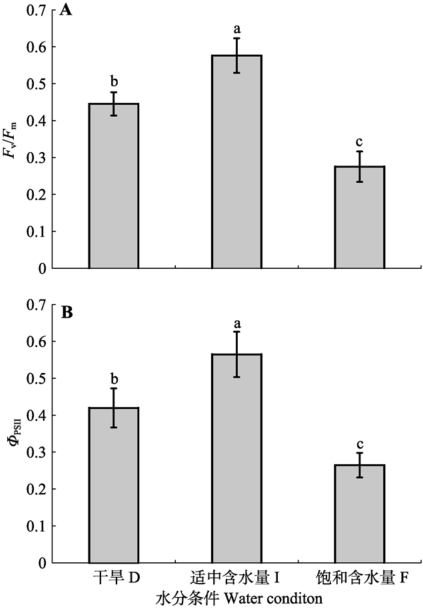 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2人工培养齿肋赤藓光系统II (PSII)的最大光化学效率(Fv/Fm)和实际光化学效率(ΦPSII)对不同水分处理的响应(平均值±标准偏差)。不同字母表示差异显著(p < 0.05)。
-->Fig. 2Responses of maximal photochemical efficiency of PSII (Fv/Fm) and actual photochemical efficiency (ΦPSII) in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD). D, drought treatment; I, intermediate water supply; F, fully watered. Different lowercase letters indicate significant differences (p < 0.05).
-->
2.4 不同水分条件处理对人工培养齿肋赤藓渗透调节物质含量的影响
由图3可知, 在中度湿润处理时, 人工培养齿肋赤藓的可溶性糖及脯氨酸含量最低, 随着水分含量的增加, 两者含量呈现先降低后缓慢升高的趋势, 干旱处理下, 两者含量最高, 显著高于中度湿润处理和完全湿润处理(p < 0.05)(图3A、3B); 相反, 人工培养齿肋赤藓的可溶性蛋白含量在中度湿润处理时含量最高, 随着水分含量的增加, 其含量呈现先升高后迅速降低的趋势, 完全湿润处理时, 其含量最低(图3C)。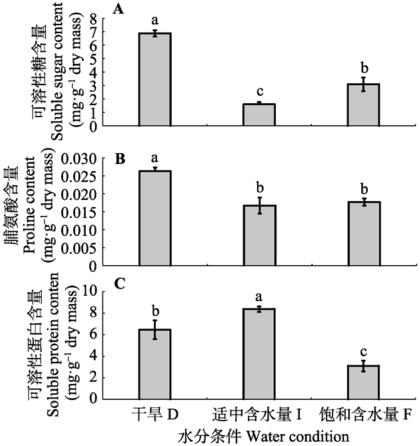 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3人工培养齿肋赤藓渗透调节物质含量对不同水分处理的响应(平均值±标准偏差)。不同字母表示差异显著(p < 0.05)。
-->Fig. 3Reponses of osmolyte content in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD). D, drought treatment; I, intermediate water supply; F, fully watered. Different lowercase letters indicate significant differences (p < 0.05).
-->
2.5 不同水分条件处理对人工培养齿肋赤藓抗氧化酶活性的影响
植物在受到逆境胁迫时, 抗氧化酶类(CAT、POD、SOD)能有效地清除活性氧, 保证细胞正常的生理功能, 维持其对逆境胁迫的抗性。由图4可知, 人工培养齿肋赤藓的3种抗氧化酶(CAT、POD、SOD) 活性在3种不同的水分条件处理下表现出一定的差异。随着水分含量的增加, 人工培养齿肋赤藓的抗氧化酶(CAT、POD、SOD)活性呈现先降低后升高的变化趋势, 在中度湿润处理时, 3种抗氧化酶活性均最低, 干旱处理下, 3种酶活性均最高; CAT及SOD活性在完全湿润的处理下虽高于中度湿润处理, 但差异不显著(p > 0.05)(图4A、4C); POD活性在干旱处理及完全湿润处理下无显著差异(p > 0.05), 但均显著高于中度湿润处理(p < 0.05)(图4B)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4人工培养齿肋赤藓抗氧化酶活性对不同水分处理的响应(平均值±标准偏差)。CAT, 过氧化氢酶; POD, 过氧化物酶; SOD, 超氧化物歧化酶。不同字母表示差异显著(p < 0.05)。
-->Fig. 4Reponses of antioxidant enzyme activities in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD). D, drought treatment; I, intermediate water supply; F, fully watered. CAT, catalase; POD, peroxidase; SOD, superoxide dismutase. Different lowercase letters indicate significant differences (p < 0.05).
-->
2.6 不同水分条件处理对人工培养齿肋赤藓MDA含量的影响
不同水分处理下人工培养齿肋赤藓MDA含量变化显著(图5)。干旱处理下, 其含量最高, 随着水分含量的增加, 其含量呈现出先降低后缓慢升高的趋势, 干旱处理时, 其含量最高, 显著高于中度湿润处理及完全湿润处理(p < 0.05)。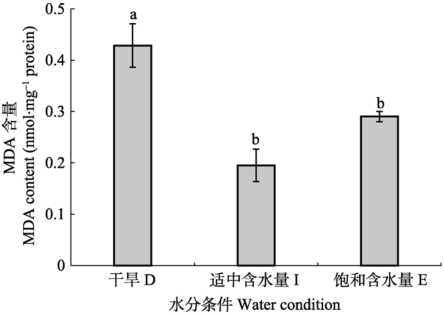 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5人工培养齿肋赤藓的丙二醛(MDA)含量对不同水分处理的响应(平均值±标准偏差)。不同字母表示差异显著(p < 0.05)。
-->Fig. 5Response of malondialdehyde (MDA) content in artificially cultivated Syntrichia caninervis to different water treatments (mean ± SD). D, drought treatment; I, intermediate water supply; F, fully watered. Different lowercase letters indicate significant differences (p < 0.05).
-->
3 讨论
3.1 人工培养齿肋赤藓植株含水量对不同水分条件处理的响应
水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(Brown & Ernest, 2002; 郑云普等, 2009; Benassi et al., 2011)。研究表明野外原生齿肋赤藓在不同水分条件(如降雨、积雪融化等)下, 植株含水量会发生显著变化, 即随着降雨量的增加或降雪后藓株含水量会显著升高(张静和张元明, 2014; 尹本丰和张元明, 2015)。而野外取材经室内培养后所得人工培养齿肋赤藓也如原生齿肋赤藓一样对水分具有较高的利用效率, 能够快速有效地吸收水分, 但干旱和完全湿润处理的植株叶片表现出一定的紧皱发黄现象, 说明这两种加水处理不利于藓株的生长。3.2 人工培养齿肋赤藓光合色素含量对不同水分条件处理的响应
光合色素反应植物对环境中光资源的利用情况, 其中叶绿素在光合作用中起核心作用, 其含量与光合速率紧密相关, 既可反映植物叶片光合作用功能的强弱, 也可用以表征逆境胁迫下植物组织器官的受损状况(魏美丽和张元明, 2009)。本研究中干旱处理(每3天加水)下叶绿素a及叶绿素b含量均下降, 叶绿素a含量下降显著, 这与Bu等(2015)在研究影响毛乌素沙地藓结皮野外定植发育的关键因素得出每隔2天加水处理高于每天加水及每6天加水处理藓结皮的叶绿素a含量结果一致。叶绿素a更多地结合在光系统反应中心上, 其功能主要是将汇聚的光能转变为化学能进行光化学反应, 而叶绿素b是光能捕获者, 主要负责收集光能(Rodriguez & Redman, 2005)。干旱处理主要影响了叶绿素a的含量, 这可能是因为活性氧降低了叶绿素合成酶的活性, 阻碍了叶绿素的合成, 另一方面加速叶绿素的分解, 造成光合色素含量下降, 从而降低了人工培养齿肋赤藓的光合效率。相关研究表明, 频繁的小于1.25 mm的降雨量会导致野外藓结皮碳收支失衡, 光合作用效率降低, 藓株色素含量随之降低(Reed et al., 2012)。完全湿润处理下(模拟设置的降雨量为2 mm, 大于1.25 mm)每天加水, 频次过高, 水分过剩, 阻碍了藓株光合作用的碳交换, 由此造成叶绿素含量显著下降。叶绿素a/b值的变化可以反映叶片光合活性的变化, 本试验结果得出干旱及完全湿润处理下人工培养齿肋赤藓的叶绿素a/b值的显著降低, 说明两种水分条件处理均在一定程度上抑制了人工培养齿肋赤藓的光能转化。类胡萝卜素不仅是光能的捕获者, 也具有防护叶绿素伤害的功能(邓馨等, 2000), 干旱及完全湿润处理下人工培养齿肋赤藓的类胡萝卜素含量均有所上升, 从而在一定程度上缓解了2种水分条件处理对其叶片光合作用的抑制。3.3 人工培养齿肋赤藓光化学效率对不同水分条件处理的响应
光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(Maxwell & Johnson, 2000)。作为PSII的光化学活性的度量, Fv/Fm指的是PSII的最大光化学效率, 即植物叶片经过充分暗适应后PSII最大的或潜在的量子效率指标, 可以反映PSII反应中心的活性, ΦPSII表示PSII反应中心受到环境胁迫时, 存在部分反应中心关闭情况下的实际光化学效率。在非胁迫条件下Fv/Fm数值通常在0.76-0.83 (Proctor, 2003), 本研究结果显示, 随着加水频次的增加, 人工培养齿肋赤藓的Fv/Fm及ΦPSII均呈现先升高后降低的趋势, 干旱处理下人工培养齿肋赤藓的Fv/Fm降至0.45, 说明其叶绿体光合活性受到了抑制, 叶片光系统电子和质子缓冲库容量变小, 直接影响了光合作用的电子传递, 最终导致PSII的实际光化学效率ΦPSII也显著下降, 光合效率降低(吴金芝等, 2015), 这与衣艳君和刘家尧(2007)、Beckett等(2000)的研究结果一致。他们在对毛尖紫萼藓(Grimmia pilifera)以及波叶仙鹤藓(Atrichum undulatum)等进行脱水干旱胁迫时, 发现其Fv/Fm和ΦPSII均下降。完全湿润处理下, 人工培养齿肋赤藓的Fv/Fm及ΦPSII最低, 这与相关原生齿肋赤藓在最适水分条件下其光化学效率最高, 随着水分含量的增加, 光化学效率逐渐降低的研究结果(张静和张元明, 2011)一致。此外, 针对维管植物的相关研究也得到相同的结果, 如高水分含量条件下, 秋华柳(Salix variegata)植株叶片PSII的Fv/Fm也表现出一定程度的降低(罗芳丽等, 2007)。由此说明高水分含量条件下所导致的低光及低氧环境会影响植株叶片的气孔开度及CO2的扩散速率, 从而使植株叶片的光化学活性降低(Mielke et al., 2003), 但降低程度还与植株生境及所设置的水分含量高低有关(刘泽彬等, 2014)。3.4 不同水分条件处理对人工培养齿肋赤藓渗透调节物质的影响
渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行。因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(Pandey et al., 2010)。脯氨酸作为植物蛋白质组分之一, 以游离状态广泛存在于植物体内, 是植物重要的渗透调节物质(Singh et al., 1972)。研究表明, 逆境胁迫下会引起藓株游离脯氨酸含量的增加, 以此来保护细胞减轻损害(吴楠等, 2009)。本文研究发现, 干旱处理下人工培养齿肋赤藓的游离脯氨酸含量最高, 说明干旱处理能够诱导人工培养齿肋赤藓植株体内游离脯氨酸的积累, 从而免于藓株细胞因过度缺水失活, 也表明人工培养齿肋赤藓对干旱具有一定的适应能力。完全湿润处理下, 人工培养齿肋赤藓的游离脯氨酸含量较中度湿润处理有所上升, 推测其原因可能是完全湿润处理下人工培养齿肋赤藓的蛋白质合成速率有所下降或受阻, 由此导致脯氨酸含量上升。脯氨酸含量的增加可以维持细胞渗透势, 保持细胞结构, 说明脯氨酸含量增加也是人工培养齿肋赤藓适应高水分含量条件的一种表现。糖类的代谢是植物的基础代谢之一, 干旱胁迫下可溶性糖对植物的保护作用已被诸多研究证实, 叶片中可溶性糖含量随干旱胁迫的加剧显著增加, 且耐旱性越强, 增加幅度越大(张显强等, 2004)。本研究结果表明, 干旱处理下, 人工培养齿肋赤藓植株体内的可溶性糖含量最高, 可溶性糖含量的增加不仅可为植株的生长提供必要的能量, 还能调节体内代谢, 并且稳定细胞中酶分子活性的构象, 提高植物的抗逆适应性(Albrecht et al., 2004), 这一结论在本试验中得到证实。完全湿润处理下人工培养齿肋赤藓的可溶性糖含量较中度湿润处理有所上升, 但仍低于干旱处理下人工培养齿肋赤藓的可溶性糖含量, 可能源于藓株在高水分含量条件下光合作用效率降低带来的糖类的消耗大于产出所致。此外, 植物体内蛋白质含量的变化也可反映植物受逆境胁迫的程度(康俊梅等, 2005)。研究表明不同藓类植物的耐旱程度不同, 相比土生对齿藓(Didymodon vinealis), 真藓(Bryum argenteum)的耐旱能力较强, 在重度干旱胁迫处理下其可溶性蛋白含量仅有小幅下降, 说明与土生对赤藓相比, 真藓能够忍受更极端的干旱胁迫(石勇等, 2012)。本研究中, 干旱处理及完全湿润处理下人工培养齿肋赤藓的可溶性蛋白含量较中度湿润处理均有显著下降, 说明水分过少或过多都不利于人工培养齿肋赤藓的生长, 阻碍其蛋白质的合成或使原有蛋白质受到破坏从而使得植株体内的蛋白质含量下降。但同时, 当植物受到逆境胁迫时, 体内的保护酶也会被激活而发挥作用, 被激活的保护酶种类越多, 活性越强, 蛋白质遭受的损害越少, 含量下降幅度越小(魏美丽, 2009), 干旱处理及完全湿润处理下人工培养齿肋赤藓的可溶性蛋白含量均有下降, 但干旱处理仍高于湿润处理下人工培养齿肋赤藓的可溶性蛋白含量, 这一方面源于干旱处理下所激活的保护酶含量较多, 另一方面也说明人工培养齿肋赤藓具有一定的耐旱性。3.5 不同水分条件处理对人工培养齿肋赤藓抗氧化酶活性及MDA含量的影响
当植物受到逆境胁迫时, 植物细胞内活性氧自由基产生和清除的平衡遭到破坏, 使活性氧大量富集, 它们对植物细胞具有损伤作用, 而植物细胞的抗氧化酶系统(CAT、POD、SOD)在消除或降低氧自由基带来的损伤中发挥着重要作用。CAT主要清除过氧化氢自由基, POD主要清除过氧化物自由基, SOD主要清除超氧自由基(Cho & Seo, 2005; Meloni et al., 2003), 诸多****针对逆境胁迫条件下苔藓植物抗氧化酶系统方面已开展了多项研究, 但是有关干旱胁迫下苔藓植物体内抗氧化酶类活性的变化并不一致, 有结果表明, 随着干旱胁迫的加剧, 土生对齿藓CAT活性无显著变化, POD活性则呈先上升后下降的趋势, SOD活性也无显著变化; 杨武(2008)研究发现, 多蒴灰藓(Hypnum fertile)抗氧化酶活性随含水量的下降均呈持续上升的变化趋势。本研究结果表明, 人工培养齿肋赤藓当水分含量较低, 即处于干旱处理时其CAT、POD、SOD活性显著较高, 随着水分含量的增加, 人工培养齿肋赤藓的抗氧化酶(CAT、POD、SOD)活性均呈现先降低后缓慢升高的趋势, 湿润处理条件下人工培养齿肋赤藓的抗氧化酶(CAT、POD、SOD)活性较高, 但仍低于干旱处理下其抗氧化酶活性。这一结果与杨武(2008)的研究结果较为一致。这可能说明面对干旱胁迫, 不同的藓类植物中清除其体内活性氧自由基中各抗氧化酶所起的作用不同, 土生对赤藓的抗氧化酶中主要是POD发挥主要作用, CAT和SOD起辅助作用, 而多蒴灰藓和本研究中的人工培养齿肋赤藓的3种抗氧化酶均发挥有效作用以缓解逆境胁迫对藓株细胞膜系统造成的氧化损伤。针对野外生境中齿肋赤藓的研究也表明, 当降雨量增加时, 其抗氧化酶活性会随之递减(张静和张元明, 2014), 且降雨量过大时, 藓株体内产生的大量活性氧自由基更会加剧齿肋赤藓的膜脂过氧化程度, 对藓株造成一定的损害, 这可能是因为高水分条件下所导致的低氧及低光照环境引起植物细胞活性氧自由基的大量积累而对植物产生毒害作用, 植物自身的抗氧化酶类由此发挥作用以减轻活性氧积累对植物的伤害(Islam & Macdonald, 2004)。MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(Oliver et al., 2000) , 是胁迫重要的生理指标。该研究表明, 干旱处理下人工培养齿肋赤藓的MDA含量最高, 随着水分含量的增加, 人工培养齿肋赤藓植株的MDA含量逐渐降低, 干旱处理下, 水分含量短缺, 无法满足人工培养齿肋赤藓正常生长过程中对水分的需求, 致使细胞膜处于渗透胁迫, 引发细胞质膜过氧化反应, 使得人工培养齿肋赤藓植株体内MDA含量显著高于中度湿润处理及完全湿润处理。杜晓等(2012)对金发藓(Polytrichum commune)和湿地匐灯藓(Plagiomnium acutum)的脱水胁迫实验也证实, 随着脱水时间的增加两种藓植株的MDA含量显著升高。研究认为, MDA质量摩尔浓度与植物抗旱性密切相关, 但MDA大量增加时, 体内细胞受到较严重的破坏(郭郁频等, 2014)。本试验干旱处理对人工培养齿肋赤藓造成了重度干旱胁迫, 致使其体内活性氧自由基大量积累, 尽管其抗氧化酶活性显著升高, 可能本试验胁迫处理直接, 没有设置过渡梯度, 致使抗氧化酶一时无法及时有效地发挥作用, 引发膜脂过氧化反应, 膜脂遭到活性氧攻击后严重受损, 膜透性增大, 造成MDA的积累。完全湿润处理下, 人工培养齿肋赤藓的MDA含量显著高于中度湿润处理, 这可能是由于高水分含量条件下的缺氧环境确实对细胞膜正常代谢活动造成一定威胁, 但由此激发藓株自身抗氧化酶很快发挥作用, 有效地减少了膜脂过氧化作用造成的MDA的大量积累。综上所述, 由藓类植物和土壤相互作用形成的藓结皮与生境、光照、生物组成及水分等多种因素具有复杂的因果关系(曹同等, 1997; 赵允格和许明祥, 2010; Reynolds & McLetchie, 2011), 而相关室内人工培养和野外恢复藓类植物促进藓结皮层形成试验研究都表明水分条件是最关键的限制因素, 对水分的需求远远大于对光照以及营养基质等的需求(Kidron et al., 2010; Stark et al., 2011), 在藓类植物人工培养的初期都表现出较快的生长和繁殖速度, 这不是光合作用的结果, 而是消耗繁殖体内储存的营养物质的结果, 达到一定的生长限度后将进入稳定或衰退状态, 此时给予良好的水分条件将有助于植株有效地进行光合作用, 进一步维持生长, 这在本研究中也得到了证实。在本研究中, 干旱处理及完全湿润下人工培养齿肋赤藓的总叶绿素含量及光化学效率均显著较低, 同时导致人工培养齿肋赤藓体内膜脂过氧化程度升高, 同时引发其体内脯氨酸及可溶性糖大量积累, 以及抗氧化酶活性的显著上升, 这些都有效地保护了其细胞免遭干旱胁迫的过度损害。与干旱处理相比, 完全湿润处理下人工培养齿肋赤藓的细胞膜趋于稳定, 但其叶绿素含量及光化学效率都显著较低, 不利于人工培养齿肋赤藓的生长。而中度湿润处理下, 人工培养齿肋赤藓表现出较优的光合生理活性, 说明此水分条件人工培养齿肋赤藓能够较好地生长。我们认为, 人工培养齿肋赤藓对于不同的水分条件处理虽然表现出了一定的适应性, 但水分胁迫也对其生长有一定的抑制作用, 中度湿润处理下, 人工培养齿肋赤藓能保持较好的生长态势, 因此可以考虑在人工培养藓植物后期采用每3天的加水处理方式, 在有效地利用水资源的基础上, 缓解人工培养藓株后期的衰退现象, 为人工培养藓类植物野外定植的节约型水分管理模式提供理论依据, 从而加速人工培养荒漠藓类应用到野外发挥防风固沙生态功能的进程。
致谢 由衷地感谢中国科学院新疆生态与地理研究所张静博士、吴林博士、尹本丰硕士以及新疆师范大学本科生田曼在样品采集和数据分析过程中给予的帮助。
The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | In terms of gene expression and carbohydrate metabolism, the response of wheat seedlings to hypoxia is dramatically different from the anoxic response. Total carbohydrate content of roots increased 4-fold during 6 days of hypoxia, with a 17-fold increase in fructans. In contrast, anoxically treated roots depleted all soluble carbohydrates and died within 72 h. Gas exchange measurements (CO 2 release vs. O 2 uptake) demonstrate that hypoxia establishes a new balance between fermentation and aerobic respiration in the roots without altering the flux of carbon through glycolysis. Furthermore, the respiratory component of this new balance is 55% higher in roots that have been hypoxically pretreated compared to non-hypoxically pretreated roots. The establishment of this new homeostasis under hypoxia involves the induction of glycolytic (aldolase and enolase) and fermentative enzymes (pyruvate decarboxylase, alcohol dehydrogenase, and lactate dehydrogenase). Enzyme induction is generally complete within 24 h with mRNA induction occurring primarily during Period I (0–6 h of hypoxia), and maximal enzymes activities attained during Period II (6–24 h of hypoxia). Accumulation rates of Suc, hexoses, and fructans also change during Periods I and II. By the start of Period III (24–144 h of hypoxia), the metabolic adjustments are complete and fructans are the major carbohydrate accumulated. In anoxia, the pattern of enzyme induction was dramatically different: aldolase was not induced and declined throughout the treatment. Alcohol dehydrogenase, pyruvate decarboxylase, and lactate dehydrogenase were induced as in hypoxia, but rapidly declined within 72 h of anoxia. Only enolase exhibited a similar expression pattern in both anoxia and hypoxia. |
| [2] | |
| [3] | Pulse amplitude modulation fluorescence was used to investigate whether abscisic acid (ABA) pretreatment increases the desiccation tolerance of photosynthesis in the moss Atrichum undulatum . In unstressed plants, ABA pretreatment decreased the F V / F m ratio, largely as a result of an increase in F o . This indicated a reduction in energy transfer between LHCII and PSII, possibly hardening the moss to subsequent stress by reducing the production of the reactive oxygen species near PSII. During desiccation, F 0 , F m , F v / F m , 桅PSII, and NPQ and F 0 quenching declined in ABA-treated and nontreated mosses. However, during rehydration, F 0 , F m , F v / F m , and 桅PSII recovered faster in ABA-treated plants, suggesting that ABA improved the tolerance of photosystem聽II to desiccation. NPQ increased upon rehydration in mosses from both treatments, but much more rapidly in ABA-treated plants; during the first hour of rehydration, NPQ was two-fold greater in plants treated with ABA. F 0 quenching followed a similar pattern, indicating that ABA treatment stimulated zeaxanthin-based quenching. The implications of these results for the mechanisms of ABA-induced desiccation tolerance in A.聽undulatum are discussed. |
| [4] | Abstract Maintaining soil stability and normal water and nutrient cycles in desert systems is critical to avoiding desertification. These particular ecosystem processes are threatened by trampling of livestock and people, and by off-road vehicle use. Soil compaction and disruption of cryptobiotic soil surfaces (composed of cyanobacteria, lichens, and mosses) can result in decreased water availability to vascular plants through decreased water infiltration and increased albedo with possible decreased precipitation. Surface disturbance may also cause accelerated soil loss through wind and water erosion and decreased diversity and abundance of soil biota. In addition, nutrient cycles can be altered through lowered nitrogen and carbon inputs and slowed decomposition of soil organic matter, resulting in lower nutrient levels in associated vascular plants. Some cold desert systems may be especially susceptible to these disruptions due to the paucity of surface-rooting vascular plants for soil stabilization, fewer nitrogen-fixing higher plants, and lower soil temperatures, which slow nutrient cycles. Desert soils may recover slowly from surface disturbances, resulting in increased vulnerability to desertification. Recovery from compaction and decreased soil stability is estimated to take several hundred years. Re-establishment rates for soil bacterial and fungal populations are not known. The nitrogen fixation capability of soil requires at least 50 years to recover. Recovery of crusts can be hampered by large amounts of moving sediment, and re-establishment can be extremely difficult in some areas. Given the sensitivity of these resources and slow recovery times, desertification threatens million of hectares of semiarid lands in the United States. |
| [5] | Three prominent life history patterns attributed to dioecious species of mosses (low sex expression, male rarity, and low frequencies of sexual reproduction) were investigated in the desert moss Syntrichia caninervis along an elevation gradient in the Spring Mountains of the Mojave Desert (U.S.A.). Low elevation patches exhibited significantly lower frequencies of sex expression, individuals with fewer inflorescences, and fewer male individuals. Sexual reproduction was infrequent at all elevations, with only seven of 85 patches showing evidence of recent sporophyte production and only nine of 708 perichaetia being fertilized. After factoring out abortive sporophytes, the percentage of patches maturing sporophytes was 5.9% and the estimated fraction of fertilized perichaetia was 0.7%. In addition, as elevation increased from low to middle/high elevation, individuals of Syntrichia caninervis exhibited significant increases in biomass, total stem length, number of ramets, and ramet length. The decreased availability of water at lower elevations may stunt individual plant size, inhibiting sex expression, and promoting growth of female-only individuals, thereby depressing sexual reproduction. Our data are consistent with the hypothesis that male rarity may be caused in part by differential desiccation tolerance between the sexes, with males less able to tolerate repeated cycles of hydration and desiccation due to their higher energetic requirements for sex expression. |
| [6] | The effects of long-term grazing (40 years) on cryptogamic crusts of Navajo National Monument were investigated. Both vascular and nonvascular communities were heavily impacted with the cryptogamic community showing the greatest reduction in cover. Lichens and mosses were the most damaged, while the algae were much more tolerant. Individual cryptogam species were affected in similar patterns with all identifiable species showing reduced cover. Vascular plant species were also affected with grasses showing the greatest reduction under grazing pressure. |
| [7] | Water is the lifeblood of the desert. It comes in rains that are typically scant and sporadic, but can be so intense as to cause flooding. Because water is the resource in shortest supply, the amount and timing of precipitation directly limits plantgrowth and primary production. Seasons of exceptionally heavy and frequent rains produce the spectacular desert blooms shown in nature films and magazines. Seasons of exceptionally high rainfall are also thought to cause increases in rodent populationsand outbreaks of rodentborne diseases such as hantavirus and plague. El Nino is supposed to cause exceptionally heavy winter rainfall in the deserts of southwestern North America, leading in turn to plant growth, abundant seeds and insects, high populations of small mammals, density-dependent increases in parasites and diseases, and increased contact between rodents, their pathogens, and humans, resulting in disease epidemics. Thus, the outbreak of the Sin Nombre strain of hantavirus that killed 27 people in the Four Corners region of the southwestern United States in the summer of 1993 was attributed to the rains, plant production, and rodent increases triggered by the El Nino events of 1991-1992 and 1992-1993 (Harper and Meyer 1999). |
| [8] | Biological soil crusts (BSCs) are a widespread photosynthetic ground cover in arid and semiarid areas. They have many positive ecological functions, such as increasing soil stability, and reducing water and wind erosion. Using artificial technology to achieve the rapid development of BSCs is expected to become a low-cost and highly beneficial ecological restoration measure. In the present study, typical moss-dominated crusts in a region characterized by mobile dunes (Mu Us Sandland, China) were collected, and a 40-day cultivation experiment was performed to investigate key factors, including watering frequency, light intensity and a nutrient addition, which affect the rapid development of moss crusts and their optimal combination. The results demonstrated that watering frequency and illumination had a significant positive effect (P=0.049, three-factor ANOVA) and a highly significant, complicated effect (P=0.000, three-factor ANOVA), respectively, on the plant density of bryophytes, and a highly significant positive effect on the chlorophyll a and exopolysaccharide contents (P=0.000, P=0.000; P=0.000, P=0.000; one-way ANOVA). Knop nutrient solution did not have a significant positive but rather negative effect on the promotion of moss-dominated crust development (P=0.270, three-factor ANOVA). Moss-dominated crusts treated with the combination of moderate-intensity light (6,000 lx) + high watering frequency (1 watering/2 days) - Knop had the highest moss plant densities, while the treatment with high-intensity light (12,000 lx) + high watering frequency (1 watering/2 days) + Knop nutrient solution had higher chlorophyll a contents than that under other treatments. It is entirely feasible to achieve the rapid development of moss crusts under laboratory conditions by regulating key factors and creating the right environment. Future applications may seek to use cultured bryophytes to control erosion in vulnerable areas with urgent needs. |
| [9] | , 16, |
| [10] | , 6, |
| [11] | We have developed a Cd-resistant type (RT) from a Cd-sensitive wild-type (WT) of Arabidopsis thalaiana, and seedlings were grown on MS medium containing up to 500 渭M of CdCl 2 to study the relationship between Cd-induced phytoxicity and oxidative stress. Compared to WT, RT showed a higher level of survival, and lower hydrogen peroxide and lipid peroxidation levels expressed as thiobarbituric acid reacting substances (TBARS) production. These results indicate that RT seedlings experienced lower oxidative stress from Cd exposure. Furthermore, compared to WT, RT had significantly higher activities of superoxide dismutase (SOD and enzymes related to hydrogen peroxide removal such as guaiacol peroxidase (GPX), ascorbate peroxidase (APX) and glutathione reductase (GR). The differential between the responses of WT and RT seedlings to Cd suggests that Cd-induced phytotoxicity can be induced by hydrogen peroxide accumulation and subsequent oxidative stress. The results also imply that a lower hydrogen peroxide accumulation confers Cd-tolerance in seedlings. |
| [12] | 复 苏植物是研究植物耐脱水机制的良好材料 ,因为它们能忍耐极度干旱 ,以一种类似休眠的方式度过严酷的干旱期 ,在水分适宜时重新恢复生活力[1- 3] 。正是因为在完全脱水后还可复苏 ,所以它们在脱水复水过程中所独具的生理生化特点必然具有某种适应意义。深入研究这些特点不但 |
| [13] | 以来自不同水分生境的金发藓和湿地匐灯藓为材料,对二者在脱水与 复水胁迫条件下的活性氧代谢、脂质过氧化损伤程度及其抗氧化系统应答的差异进行比较研究.结果显示:在脱水与复水过程中,(1)硅胶快速脱水更接近阳光直 射条件下藓类植物的水分丧失.(2)随着含水量的变化,湿地匐灯藓虽然能够在复水后迅速修复细胞的完整性,但变化剧烈;金发藓则能够始终维持较低的膜透 性.(3)2种藓类植物的丙二醛(MDA)含量变化均呈先升后降趋势,但金发藓的MDA含量明显低于湿地匐灯藓.(4)2种藓类植物的超氧阴离子自由基 (O2-·)产生速率和过氧化氢含量(H2O2)的变化均与MDA含量变化相似,且金发藓活性氧水平明显高于湿地匐灯藓.(5)2种藓类植物的超氧化物歧 化酶(SOD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)活性受活性氧诱导亦呈先升后降的趋势,但金发藓抗氧化酶对活性氧迸发的应答更快,活 性更强.(6)2种藓类植物的抗坏血酸(AsA)含量呈先降后升态势,但金发藓的含量低于湿地匐灯藓.研究表明,来自不同生境的2种藓类植物对脱水胁迫所 致的氧化胁迫均具有很强的适应能力,尤其是复水过程中的修复能力,但不同藓类可能通过不同途径和机制来适应脱水所致的氧化胁迫;来自易发生水分亏缺生境的 金发藓可能因具有更强抗氧化能力,从而获得比来自水分充沛生境的湿地匐灯藓更高的脱水耐性. |
| [14] | Abstract In vitro cultivation is not only essential for the use of bryophytes in cellular, developmental and molecular research but is also vital to the elucidation of the roles of juvenile stages in reproductive biology. Other important uses include the discovery of new characters for systematics, the conservation of rare taxa and understanding the functional significance of fungal and cyanobacterial association in hepatics and hornworts. This article, based on the culturing of over 300 mosses and nearly 50 hepatics sets out the practicalities of in vitro cultivation and reviews the potential rewards. Cultures, initiated from surface-sterilized spores, gemmae or vegetative fragments, are most easily handled in Petri dishes on media containing inorganic salts. Solidification of the medium with Phytogel or Gelrite is preferred to traditional agar because of toxic impurities in the latter. Dilution of the nutrients can be beneficial to the growth of some taxa. Prior knowledge of the seasonality of sporophyt... |
| [15] | A portable wind tunnel was used to test the contribution of biological and physical elements to overall soil aggregation on a soil dominated by biological soil crusts in south-eastern Australia. After moderate disturbance and simulated wind erosion, 90% of surface aggregates on the loamy soil and 76% on the sandy soil were dominated by biological elements (cryptogams). Lower levels of biological bonding were observed on the severely disturbed treatment. Linear regression indicated a significant positive relationship ( r 2 =0·72) between biological soil crust cover and dry aggregation levels greater than 0·85mm. To maintain sediment transport below an erosion control target of 5gm 611 s 611 for a 65kmh 611 wind at 10m height, a crust cover of approximately 20% is required. When a multiple regression model which sequentially fitted biological crust cover and dry aggregation greater than 0·85mm was applied to the data, dry aggregation accounted for more of the variation in sediment transport rate than biological crust cover. These data were used to develop a conceptual model which integrates crust cover and dry aggregation, and provides a useful framework within which to predict the likely impacts of changes in soil crust cover and aggregation. |
| [16] | Abstract The lichen, Punclelia subrudecta (Ny1.) Krog., was transplanted to 10 biomonitoring sites during the period December 1990-March 1991. the total amounts of metals (Mn, Cu, Pb, Zn, Fe and A1) detected in the lichen thalli after the period of exposure, were compared with the chlorophyll degradation, amounts of sulphur, MDA concentrations and soluble protein concentrations in the same material. the MDA content was directly related to the amount of sulphur in the lichen transplanted material. A contamination index (C.I.) was calculated from the amounts of sulphur, chlorophyll-伪, phaeophytin-伪, and MDA. |
| [17] | 为鉴定评价早熟禾幼苗抗旱性,将14个早熟禾品种的种子经消毒后播种在塑料花盆中,出苗后间苗,2~3个真叶定株,每盆留生长一致、分布均匀的幼苗15株,定期浇水,以保证幼苗的正常生长。待生长到3~4个真叶时进行干旱处理,依据Michel公式计算不同水势所需的PEG-6000的量,然后加入PEG-6000溶液进行干旱处理,处理的水势梯度为0MPa(即为对照),-0.3,-0.6,-0.9和-1.2MPa。处理后每2d进行称重补水以维持溶液浓度。设干旱处理与对照(正常浇水)两组,对14个早熟禾品种幼苗进行模拟干旱胁迫,4次重复。测定叶绿素(CHL)含量、丙二醛(MDA)含量、脯氨酸(Pro)含量、过氧化物酶(POD)以及超氧化物歧化酶(SOD)活性等生理生化指标。结果表明,随着干旱胁迫的加剧,早熟禾幼苗叶片CHL含量呈下降趋势;Pro含量、MDA含量、SOD活性总体呈上升趋势;POD活性总体呈先降再升后降的趋势;利用隶属函数分析得出14个早熟禾品种的抗旱性强弱顺序依次为:3号恩托佩草地早熟禾10号细叶早熟禾1号优异草地早熟禾2号斗士草地早熟禾=9号蓝钻草地早熟禾7号草地早熟禾11号草地早熟禾4号草地早熟禾5号瓦巴斯草地早熟禾8号早熟禾=13号肯塔基草地早熟禾14号草地早熟禾6号莫诺波利草地早熟禾=12号菲尔津草地早熟禾。 |
| [18] | Black spruce [ Picea mariana (Mill.) B.S.P.] and tamarack [ Larix laricina (Du Roi) K. Koch] are the predominant tree species in boreal peatlands. The effects of 34 days of flooding on morphological and physiological responses were investigated in the greenhouse for black spruce and tamarack seedlings in their second growing season (18 months old). Flooding resulted in reduced root hydraulic conductance, net assimilation rate and stomatal conductance and increased needle electrolyte leakage in both species. Flooded tamarack seedlings maintained a higher net assimilation rate and stomatal conductance compared to flooded black spruce. Flooded tamarack seedlings were also able to maintain higher root hydraulic conductance compared to flooded black spruce seedlings at a comparable time period of flooding. Root respiration declined in both species under flooding. Sugar concentration increased in shoots while decreasing in roots in both species under flooding. Needles of flooded black spruce appeared necrotic and electrolyte leakage increased over time with flooding and remained significantly higher than in flooded tamarack seedlings. No visible damage symptoms were observed in flooded tamarack seedlings. Flooded tamarack seedlings developed adventitious roots beginning 16聽days after the start of flooding treatment. Adventitious roots exhibited significantly higher root hydraulic conductivity than similarly sized flooded tamarack roots. Flooded black spruce lacked any such morphological adaptation. These results suggest that tamarack is better able to adjust both morphologically and physiologically to prolonged soil flooding than black spruce seedlings. |
| [19] | |
| [20] | 利用SDS-PAGE电泳分析法探讨干旱对供试苜蓿品种叶片可溶性蛋白的变化及其与干旱强度的关系。结果表明:可溶性蛋白的变化与干旱强度有直接关系,随着干旱胁迫强度的增加某些可溶性蛋白的变化表现为先增强后减弱;干旱胁迫诱导蛋白量的变化各异,抗旱性强的品种能诱导基因更强的表达以适应干旱胁迫,而抗旱性较弱的品种可能减弱或丧失其自我调节能力,产生的干旱诱导蛋白量比前者少;在干旱15 d时,抗旱性强的品种均被诱导产生新的蛋白带,只有两个抗旱性弱的品种出现该谱带;供试苜蓿品种的抗旱性存在明显的遗传多样性,其中抗旱性强的品种是苜蓿抗旱性选育的重要种质资源。 |
| [21] | Microbiotic crusts play an important role in arid and semi-arid regions. Yet, very little information exists regarding the factors that impact their development. In an attempt to assess the main factors that may determine their growth, measurements of the amount of fines (silt and clay), rain, moisture content, wetness duration and wind erosion and deposition were carried out along a 12 station transect within a partially crusted dune field in the western Negev Desert and compared to the crust cover and chlorophyll content. Surface stability was the only variable that exhibited significant relationship with crust cover while daylight wetness duration exhibited strong positive relationship ( r 2 = 0路92-0路99) with the crust's chlorophyll content. The data point out that microbiotic crusts may serve as a useful biomarker for surface stability. While wetness duration and wind will control crust cover and the crust chlorophyll content in semi-stable habitats (with absolute annual change in sand level of 2-3 mm), stable habitats (absolute change <1 mm) will be controlled primarily by moisture, while habitats with low surface stability (absolute change of tens and hundreds of millimeters) will be primarily controlled by wind. Furthermore, owing to the strong positive relationship between daylight wetness duration and the crust's chlorophyll content, the crust may serve as a useful biomarker for the quantification of surface wetness duration. Copyright 漏 2009 John Wiley & Sons, Ltd. |
| [22] | When a cluster analysis was performed, five types of microbiotic crusts were defined, four of which were cyanobacterial (A鈥揇) and one moss-dominated crust (E). The crusts differed in their physical and biological properties. They showed an increase in chlorophyll content, protein, carbohydrates and organic matter from A to E, with concomitant increase in species diversity, thickness, roughness and strength, but with some variables (crack density and infiltrability) showing a reversed trend at the moss-dominated crust. The increase in the biomass components of the crust and the gradual change of the physical properties are explained by the improved physical conditions (primarily wetness duration), which facilitates longer hours of photosynthetic activity and consequently the introduction of additional, more mesic species such as green algae, lichens and mosses. Extended wetness duration was found to shift the crust type from cyanobacterial to moss-dominated crust. The spatial distribution of the crusts, as verified by crust mapping, coincided with the daylight surface duration, which in turn was controlled by topography (aspect, angle and slope position). It implies that whereas initial physical conditions dictates species composition and thus crust type, the crust type in turn is responsible for characterizing the physical properties of the surface, which may largely affect ecological and geomorphological processes. |
| [23] | 61There is heterogeneity in crust development and succession in the different regions.61K, Na, silt contents and prior biomass accumulation mainly affect lichen emergence.61Early crusts and water holding content provide the guarantee for moss germination.61A positive feedback mechanism forms between crust development and soil environments.61A negative feedback mechanism forms between free living algae and crust succession. |
| [24] | 通过设置对照(CK)、持续性水淹(CF)、间歇性水淹(PF)和全淹(FF)4个水淹处理,模拟三峡库区秋冬季消落带土壤水淹变化,研究2年生中华蚊母树的生理生态适应机制。结果表明: 不同水淹形式并没有显著影响中华蚊母树的株高和基径,而光合色素含量、光合及叶绿素荧光特性在不同水淹形式下却有所差异,CF和PF植株光合色素含量、净光合速率(<em>P</em><sub>n</sub>)、气孔导度(<em>G</em><sub>s</sub>)、蒸腾速率(<em>Tr</em>)、最大光化学效率(<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>)、光化学猝灭(<em>q</em><sub>P</sub>)以及电子传递速率(ETR)均出现了显著下降,但PF植株下降的幅度要小于CF植株,除光合色素含量、<em>G</em><sub>s</sub>和<em>Tr</em>外,其他指标均差异显著,而FF植株光合色素含量、<em>P</em><sub>n</sub>、<em>G</em><sub>s</sub>、<em>Tr</em>、<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>、<em>q</em><sub>P</sub>以及ETR均未受到水淹的影响,与CK植株差异不显著; CF和PF植株叶片非光化学淬灭(<em>q</em><sub>N</sub>)未受到水淹的影响,但FF植株叶片<em>q</em><sub>N</sub>与CK相比却显著下降。虽然CF植株较其他处理植株叶绿素含量、<em>P</em><sub>n</sub>、<em>G</em><sub>s</sub>、<em>Tr、F</em><sub>v</sub>/<em>F</em><sub>m</sub><em>q</em><sub>P</sub>以及ETR均下降明显,但水淹处理植株能够通过形成不定根、特化的皮孔以及减少叶片数量来增加耐淹能力。因此,中华蚊母树具有较强的耐淹能力,在不同水淹形式下均表现出一定的适应性和可塑性。 |
| [25] | |
| [26] | Chlorophyll fluorescence analysis has become one of the most powerful and widely used techniques available to plant physiologists and ecophysiologists. This review aims to provide an introduction for the novice into the methodology and applications of chlorophyll fluorescence. After a brief introduction into the theoretical background of the technique, the methodology and some of the technical pitfalls that can be encountered are explained. A selection of examples is then used to illustrate the types of information that fluorescence can provide. |
| [27] | The effects of NaCl stress on the activity of antioxidant enzyme such as superoxide dismutase (SOD: EC 1.15.1.1), peroxidase (POD: EC 1.11.1.7), glutathione reductase (GR: EC 1.6.4.2), rate of lipid peroxidation, gas-exchange, chlorophyll content and chlorophyll fluorescence were investigated in two cotton cultivars, Guazuncho and Pora (hybrids between Gossypium hirsutum 脳 G. arboretum 脳 G. raimondii ) grown in nutrient solution. Plants were treated with three salt concentrations (50, 100 and 200 mol m 鈭3 NaCl) for 21 days. The SOD activity in Pora increases with the increase in the intensity of NaCl stress, but salt treatment had no significant effect on this enzyme activity in Guazuncho. The POD and GR activities showed similar trends under salt stress, in both cotton cultivars. In Pora, there was an average increase in GR activity of about 53%, but there was no further increase at higher NaCl concentrations. In Guazuncho, no change in GR activity was observed. Net photosynthesis and stomatal conductance decreased in response to salt stress, but Pora showed a smaller reduction in photosynthesis than Guazuncho. The results indicated that stomatal aperture limited leaf photosynthetic capacity in the NaCl-treated plants of both cultivars. However, significant reduction in the leaf chlorophyll contents due to NaCl stress was observed only on Guazuncho. In both cotton cultivars, the photochemical efficiency of PSII was not affected by salt stress. These results suggest that salt-tolerant cotton varieties may have a better protection against reactive oxygen species (ROS) by increasing the activity of antioxidant enzymes under salt stress. |
| [28] | Effects of soil flooding on photosynthesis and growth of Genipa americana L. seedlings, a neotropical fruit-tree species used in gallery forest restoration programs, were studied under glasshouse conditions. Despite the high survival rate and wide distribution in flood-prone habitats of the neotropics, previous studies demonstrated that growth of G. americana is reduced under soil flooding. Using leaf gas exchange and chlorophyll fluorescence measurements, we tested the hypothesis that stomatal limitation of photosynthesis is the main factor that reduces carbon uptake and growth rates of G. americana seedlings. Throughout a 63-day flooding period, the survival rates were 100%. The maximum values of the net photosynthetic rate ( A ) and stomatal conductance to water vapor ( g s ) of control seedlings were 9.86 μmol CO 2 m 612 s 611 and 0.525 mol H 2 O m 612 s 611 , respectively. The earliest effects of flooding were significant decreases in g s and A , development of hypertrophied lenticels and decrease in the dry weight of roots. A strong effect of the leaf-to-air vapor pressure deficit (LAVPD) on g s and A were observed that was enhanced under flooded conditions. Between 14 and 63 days after flooding, significant reductions in g s (31.7% of control) and A (52.9% of control) were observed followed by significant increments in non-photochemical quenching ( q N ) (187.5% of control). During the same period, there were no differences among treatments for the ratio between variable to initial fluorescence ( F v / F 0 ), the maximum quantum efficiency of the photosystem II ( F v / F m ) and photochemical quenching ( q P ), indicating that there was no damage to the photosynthetic apparatus. Based on the results, we conclude that decreases in stomatal opening and stomatal limitation of photosynthesis, followed by decrease in individual leaf area are the main causes of reductions in carbon uptake and whole plant biomass of flooded seedlings. |
| [29] | The development of a complete understanding of how plants interact with the environment at the cellular level is a crucial step in advancing our ability to unravel the complexities of plant ecology particularly with regard to the role that many of the less complex plants (i.e., algae, lichens, and bryophytes) play in plant communities and in establishing areas for colonization by their more complex brothers. One of the main barriers to the advancement of this area of plant biology has been the paucity of simple and appropriate experimental models that would enable the researcher to biochemically and genetically dissect the response of less complex plants to environmental stress. A number of bryophytes model systems have been developed and they have been powerful experimental tools for the elucidation of complex biological processes in plants. Recently there has been a resurgent interest in bryophytes as models systems due to the discovery and development of homologous recombination technologies in the moss Physcomitrella patens (Hedw.) Brach & Schimp. In this report we introduce the desiccation-tolerant moss Tortula ruralis (Hedw.) Gaert., Meyer, and Scherb, as a model for stress tolerance mechanisms that offers a great deal of promise for advancing our efforts to understand how plants respond to and survive the severest of stressful environments. T. ruralis, a species native to Northern and Western North America, has been the most intensely studied of all bryophytes with respect to its physiological, biochemical, and cellular responses, to the severest of water stresses, desiccation. It is our hope that the research conducted using this bryophyte will lay the foundationfor not only the ecology of bryophytes and other less complex plants but also for the role of desiccation-tolerance in the evolution of land plants and the determination of mechanisms by which plant cells can withstand environmental insults. We will focus the discussion on the research we and others have conducted in an effort to understand the ability of T. ruralis to withstand the complete loss of free water from the protoplasm of its cells. |
| [30] | Abstract Selaginella bryopteris is a lycophyte resurrection plant, which incurves during desiccation and recovers on availability of moisture. The aim of the study was to test and understand the various physiological and biochemical changes the fronds undergo during desiccation and rehydration, to get an insight as to how this plant adapts and survives through the dry phase. Upon desiccation, S. bryopteris fronds showed drastic inhibition in net photosynthesis (A) and maximal photochemical efficiency of PSII (F(v)/F(m)) however, chlorophyll content did not show much variation. Dark respiration (R(d)) continued even at 10% relative water content (RWC), and showed a burst after rehydration, which is proposed to be crucial to establish protection mechanisms. Desiccation caused an enhanced production of reactive oxygen species (ROS) and increased lipid peroxidation. Proline accumulation increased substantially by 11-fold. Sucrose and starch contents decreased upon desiccation as compared to control. The antioxidative enzymes viz. superoxide dismutase (SOD), ascorbate peroxidase (APX) and catalase (CAT) along with soluble acid invertase increased during desiccation. S. bryopteris shows mechanical as well as physiological mechanisms for tolerance to extreme levels of desiccation stress. The rapid and almost complete recovery of F(v)/F(m) after rehydration clearly indicates the absence of marked photoinhibitory or thermal injury to PSII during desiccation. This along with the homoiochlorophyllous characteristics enables S. bryopteris to recover its A. The antioxidant metabolism further plays an important role in the desiccation tolerance of S. bryopteris. Copyright 漏 2010 Elsevier GmbH. All rights reserved. |
| [31] | AbstractChlorophyll fluorescence provides a non-invasive and non-destructive method to follow various aspects of the photosynthetic function of bryophytes under relatively natural conditions, which is easy to use and can be applied to small amounts of material. Some of its potentialities (and potential pitfalls) for bryophyte desiccation physiology are outlined. Data are presented on the responses of eleven desiccation-tolerant bryophytes to drying at –41, –114, –218 and –412 MPa for periods up to ~240 days. Recovery was assessed from FV/FM after 20 min and 24 h re-wetting, and from the mean FM value after 24 h. For the more desiccation-tolerant species, Grimmia pulvinata, Syntrichia ruralis, Andreaea rothii, Racomitrium lanuginosum, R. aquaticum, Leucodon sciuroides, Pleurochaete squarrosa and Ulota crispa, long-term survival (>30–120 d) was generally best at ~–100 to –200 MPa (20–45% r.h.). The moderately desiccation-tolerant Anomodon viticulosus, Porella platyphylla and P. obtusata survived best at the... |
| [32] | Arid and semi-arid ecosystems cover ~40% of Earth鈥檚 terrestrial surface1, but we know little about how climate change will affect these widespread landscapes. Like many drylands, the Colorado Plateau in southwestern United States is predicted to experience elevated temperatures and alterations to the timing and amount of annual precipitation2, 3, 4. We used a factorial warming and supplemental rainfall experiment on the Colorado Plateau to show that altered precipitation resulted in pronounced mortality of the widespread moss Syntrichia caninervis. Increased frequency of 1.2鈥塵m summer rainfall events reduced moss cover from ~25% of total surface cover to <2% after only one growing season, whereas increased temperature had no effect. Laboratory measurements identified a physiological mechanism behind the mortality: small precipitation events caused a negative moss carbon balance, whereas larger events maintained net carbon uptake. Multiple metrics of nitrogen cycling were notably different with moss mortality and had significant implications for soil fertility. Mosses are important members in many dryland ecosystems and the community changes observed here reveal how subtle modifications to climate can affect ecosystem structure and function on unexpectedly short timescales. Moreover, mortality resulted from increased precipitation through smaller, more frequent events, underscoring the importance of precipitation event size and timing, and highlighting our inadequate understanding of relationships between climate and ecosystem function in drylands. |
| [33] | |
| [34] | Fossil records suggest that bacteria developed the ability to photosynthesize ≈3,500 million years ago (mya), initiating a very slow accumulation of atmospheric oxygen (1). Recent geochemical models suggest that atmospheric oxygen did not accumulate to levels conducive for aerobic life until 500–1,000 mya (2,023). The oxygenation of Earth's atmosphere resulted in the emergence of aerobic organisms followed by a great diversification of biological species and the eventual evolution of humans. |
| [35] | |
| [36] | |
| [37] | Climate change in the Mojave Desert (USA) may result in a greater intensity of summer (monsoon) rain events and greater atmospheric N deposition. Patches of the dominant biological crust moss Research highlights? An added precipitation treatment resulted in lower shoot biomass of Syntrichia caninervis. ? A low N treatment negatively affected shoot regeneration in S. caninervis. ? A high N treatment adversely affected the resumption of apical growth in S. caninervis. ? Desiccation stressed shoots of S. caninervis regenerated more slowly, produced lower protonemal cover, and produced fewer shoots than unstressed controls. |
| [38] | Abstract Top of page Abstract Introduction Materials and methods Results Discussion References Female plants of the moss Syntrichia caninervis are more common than male plants and are found in more thermally stressful habitats than male plants. We hypothesized that this distribution pattern is due to a more favorable regeneration response of females to thermal stress compared to that of males. Hydrated leaves from four genotypes of both sexes of S. caninervis were exposed for 60min under lighted conditions to temperatures of 20, 30, 35, 40, and 45掳C. Leaves were allowed to regenerate on native sand under recovery conditions (12-h photoperiod, 20掳C lighted, and 8掳C dark) for 56days, and over this period, protonemal emergence, growth rate, and shoot production were assessed. Leaves exposed to higher heat shocks produced protonemata significantly later, exhibited significantly reduced growth rates over the course of the experiment, and produced fewer shoots but did not differ in the probability of producing a shoot. Males tended to produce protonemata earlier than females at the highest thermal stress, whereas females tended to produce protonemata earlier under control conditions. Female leaves regenerated at twice the rate of male leaves, producing twice the area of protonemal cover; this gender difference was lessened at the highest thermal stress. Female leaves regenerated significantly more shoots than males, with each sex exhibiting different peaks of shoot production depending on the thermal stress. No interaction effect was detectable between gender and stress treatment. While females had a more favorable regeneration response relative to males, thermal stress diminished this difference, thus suggesting that our hypothesis was not supported. |
| [39] | The short-term responses and mechanisms of antioxidants in moss Hypnum plumaeforme subjected to single or combined Pb and/or Ni stress has been revealed in this study, in order to clarify (1) the relationship between the stress intensity and antioxidant fluctuation, (2) the difference between single and combined stress, and (3) the possibility of biomonitoring by the application of antioxidant fluctuation under stress. The results showed that the stress induced dose dependent formation of reactive oxygen species (ROS) and subsequent lipid peroxidation. Total chlorophyll (Chl) content and superoxide dismutase (SOD) activity were initiated under lower stress but were inhibited under higher stress. Both single and combined stress decreased catalase (CAT) activity but increased peroxidase (POD) activity, indicating POD in the moss played an important role in resisting the oxidative stress induced by Pb and Ni. The accumulation of \( \cdot O_2^ - \) and H 2 O 2 in H. plumaeforme was respectively related to the low activity of SOD and the decreased activity of CAT. The study indicated that Pb and Ni had synergistic effect in inducing the oxidative stress in moss H. plumaeforme , especially under the combination of high concentration of Ni (0.1, 1.0聽mM) and Pb. POD and CAT activity, as well as H 2 O 2 and MDA content, which increased or decreased regularly with a dose dependent under Pb and Ni stress, could be used as an effective indicator in moss biomonitoring, especially in the case of light pollution caused by heavy metals without the changes in the appearance of mosses. |
| [40] | 凝结水(隐匿降水)和降水是荒漠地区两种不同形式的水分来源,对 荒漠生态系统极为重要.叶片毛尖( leaf hair points,LHPs)是很多荒漠藓类的重要外部形态结构特征,但它在藓类植物利用凝结水和降水中的作用尚不清楚.齿肋赤藓(Syntrichia caninervis)是古尔班通古特沙漠藓类结皮的优势种,其叶片顶端具有较长的白色毛尖.采用自制微渗计(h=3.5 cm,d=5.7cm)研究了正常有毛尖和人工去除毛尖的齿肋赤藓结皮的凝结水特征及其在3种模拟降水梯度(1、3、6 mm)下的短期蒸发特征,结果表明:(1)在凝结阶段,有毛尖结皮每时段的凝结水量均大于无毛尖结皮,但日出后有毛尖结皮凝结水下降速度稍快;有毛尖结皮 的日凝结水量均大于无毛尖结皮,且前者的总凝结水量比后者多10.26%,即毛尖能增加10.26%的凝结水量.(2)3种模拟降水梯度下,有毛尖结皮在 各时段的累积蒸发率始终小于无毛尖结皮,以蒸发前期阶段最为明显,表明毛尖能显著减少和延缓结皮内的水分蒸发、延长植株水合时间,而且降水量越大减缓效果 越明显.毛尖对这2种不同形式水分利用的差异是因为利用方式不同:对凝结水的利用主要归因于毛尖增大了结皮表面粗糙度,而减少对降水的蒸发主要是毛尖能反 射阳光、形成内部毛细管及减小植株间隙的缘故.因此,毛尖的存在有利于齿肋赤藓结皮对凝结水和降水的利用,增强了藓类结皮对干旱环境的适应能力. |
| [41] | 通过藓类结皮层的自然恢复和人 工培养藓类植物促进结皮层形成试验,研究了腾格里沙漠固定沙丘生物 结皮层形成过程中优势成分真藓(Bryum argenteum)的繁殖生物学特性,结果表明:藓类结皮层人工去除后在3-4 年内70%的样方基本恢复,在此过程中真藓主要靠茎叶碎片传播和繁殖;通过分株法、撒茎叶法培养的真藓在1个 月后长满整个样地,主要通过如下方式繁殖--茎的碎片连续分枝可产生小植株,茎、叶均可产生原丝体,由原丝 体发育产生小植株,小植株又可再生原丝体,如此反复产生新植物体,这一过程是野外人工促进生物结皮层形成过 程中真藓主要的繁殖途径。与室内培养中真藓的繁殖特性相比较,野外培养的真藓在繁殖过程中产生的原丝体较 粗壮,分枝多,但在两种条件下的繁殖特性相同,能够揭示该地区自然条件下藓类植物萌发和定居的繁殖机理。该 研究为人工促进生物结皮层形成及治理受损结皮层提供了实验依据。 |
| [42] | |
| [43] | 对古尔班通古特沙漠南缘降雨模式和不同生活型植物对小雨量降水的生理响应进行了研究.通过分析 1998-2007年的降雨数据,发现本区的降水模式为小雨量,高频率,0~5 mm的小雨量降雨占总降雨频次的89.8%,平均降雨间隔在6 d左右.选择了本区具有代表性的16种植物,利用叶片相对含水量作为指标,研究了不同生活型植物对小雨量降雨(1、2、3、4和5 mm)的响应.结果表明,所有草本植物(11种)对2~5 mm的降雨都有不同程度的响应,只有1种灌木(驼绒藜Ceratoides latens),对4 mm降雨有响应.这12种植物在降雨后的第1天,叶片相对含水量显著增加,水分状况得到显著改善,表明这些植物对小雨量(≤5 mm)降雨有显著响应,而半灌木(沙漠绢蒿Seriphidium santolinum)、灌木(梭梭Haloxylon ammodendron、柽柳Tamarix ramosissima)、小乔木(白梭梭Haloxylon persicum)对5 mm以下的降雨均无显著响应,小雨量降雨可能主要是被1年生草本植物所利用.通过统计各降雨梯度上响应的植物种数发现,在降雨量达到3 mm时开始响应的植物种类最多.综合考虑降雨模式与不同生活型植物对小雨量降雨的响应特点,高频率的小雨量降雨,可能是维持本区较高植被覆盖度的重要原 因. |
| [44] | , . 应用光学显微镜和透射电子显微镜,对新疆古尔班通古特沙漠生物结 皮中齿肋赤藓(Syntrichia caninervis Mitt.)在脱水条件下叶片细胞的亚显微结构特征以及生理生化指标的变化进行了观察。此研究将为耐旱苔藓的适应性提供证据,也为干旱区生物结皮固沙作用 的发挥提供理论指导。 1.在湿润状态下,齿肋赤藓叶片背卷、中肋粗壮,多数叶片中上部具两层细胞,基部为单层细胞;透射电镜下细胞壁较厚,具疣状突起,且细胞壁通道较多,叶绿 体、细胞核、线粒体、微体、液泡等细胞器明显。其中,叶绿体中基粒片层结构和类囊体清晰,纵向排列,细胞膜和叶绿体双层膜系统... |
| [45] | 利用光学显微镜和透射电镜,对新疆古尔班通古特沙漠生物结皮中齿肋赤藓叶片细胞的结构特征进 行了观察研究。结果表明,齿肋赤藓叶片边缘背卷、中肋粗壮,多数叶片中上部具两层细胞,基部为单层细胞;透射电镜下细胞壁较厚,具疣状突起,且细胞壁通道 较多,细胞中叶绿体、细胞核、线粒体、微体、液泡等细胞器明显。其中,叶绿体中基粒片层结构和类囊体清晰,纵向排列,细胞膜和叶绿体双层膜系统保持完好。 表明生物结皮中齿肋赤藓对干旱环境具有较强的结构适应性。 |
| [46] | ABSTRACT Bryophytes have been powerful experimental tools for the elucidation of complex biological processes. Analysis of organisms from these ancient clades is an active and ongoing enterprise that will provide greater insight into the development, physiology, phylogenetics, and stress-induced cellular responses of plants. To maintain their relevance as experimental models, the analysis of mosses must expand to include modern molecular tools such as a knowledge of the genome via large-scale DNA sequencing, the ability to create transgenic individuals via transformation, and the capability to create gene knock-outs by homologous recombination. The availability of these molecular tools is limited when compared to flowering plants. However, in mosses such as Physcomitrella patens, Funaria hygrometrica, Ceratodon purpureus, and Tortula ruralis these tools are rapidly being developed for the study of molecular genetics. Efficient targeted gene disruption (i.e., homologous recombination) is a well-established tool in both yeast and murine cells that until recently was unknown in any plant model system. Recently, Schaefer and Zryd (1997) demonstrated that efficient homologous recombination occurs in P. patens. The ability to perform efficient homologous recombination in P. patens is at present unique amongst all plants and represents an extremely powerful technique for the functional analysis of plant genes. |
| [47] | 为明确不同抗旱性冬小麦品种灌浆期旗叶的叶绿素荧光特性及其与籽粒产量的关系,在防雨棚控水条件下,测定和分析了干旱胁迫后强抗旱性品种晋麦47、弱抗旱性品种偃展4110和中等抗旱性品种矮抗58旗叶荧光参数、籽粒产量和水分利用效率的变化。结果表明,随着光量子密度(PFD)的增加,冬小麦旗叶实际荧光(F)、表观电子传递速率(ETR)和非光化学淬灭系数(NPQ)值逐渐升高,PFD超过189μmol·m-2·s-2后,F值基本稳定,而光下最大荧光(F,m)值持续降低,导致其他叶绿素荧光参数改变,PFD在189~1 360μmol·m-2·s-2时F,m变化幅度较大。干旱胁迫降低了小麦旗叶F、F,m、实际量子产量(Yield)、ETR、NPQ值。不同品种荧光参数值受干旱胁迫的影响程度不同,偃展4110受影响最大,晋麦47较为稳定。干旱胁迫条件下3个品种的产量差异显著,表现为晋麦47〉矮抗58〉偃展4110。灌浆中期旗叶主要叶绿素荧光参数与产量间具有显著的相关性,可以作为高产品种筛选的生理指标;而干旱处理与适水处理的F比值、F,m比值则与两种水分条件下的产量比值呈显著正相关,可以作为品种抗旱性鉴定的生理指标。 |
| [48] | Precipitation is the major driver of ecosystem functions and processes in semiarid and arid regions. Although re-wetting pulses generate a significant portion of the total annual CO 2 exchange between atmosphere and soil, there has been little recognition of the importance of photosynthetic and respiratory activities of biological soil crusts (biocrusts) in desert soil CO 2 exchange. In this study in the Gurbantunggut Desert of northwestern China, our objective was to determine the extent to which precipitation intensity could influence soil CO 2 exchange of the desert ecosystem and the role played by moss crust in soil C balance during this process. In field experiments, net CO 2 exchange (NCE) was measured in moss crusted soil and in bareland once a month from March to November in 2013. In laboratory experiments, simulated precipitation treatments (0mm, 2mm, 5mm, 10mm and 15mm) were applied to moss crust, and NCE of moss crusted soil and its three flux components (crust photosynthesis, crust respiration, and subsoil respiration) were measured. Temporal variation of NCE varied with soil moisture and temperature. Soil moisture alone can explain 71鈥74% of variation in NCE. Soil type (moss crusted soil or bareland) also had a significant effect on NCE ( P <0.01), but this was dependent on soil moisture which is directly linked to precipitation pulse. The response of NCE to precipitation pulse in moss crust differed significantly from that of bareland. After a 2mm precipitation pulse, the crust gross photosynthetic rate (GPc) was lower than the crust respiration rate (Rc), resulting in C efflux. When precipitation intensity was equal to or greater than 5mm, GPc fully offset total respiration, resulting in an increase in C uptake. C gain was positively correlated with intensity of precipitation pulse. Regardless of different precipitation intensities, Rc was significantly higher than that of subsoil respiration. Thus, precipitation primarily drives moss crust-derived CO 2 exchange, which significantly influences the balance of soil-level CO 2 exchange in desert ecosystems. Overall, this study demonstrates that in desert ecosystems, the regulation of atmospheric-soil C balance by moss crusts depends on the intensity of precipitation. |
| [49] | 采用电导法、光镜观测、加热烘干法、茚三酮比色法、蒽酮比色法分别分析了变水过程中,刺叶赤藓(Syntrichia caninervis Mitt.)质膜透性、外部形态结构、相对含水量、游离脯氨酸、可溶性糖含量的变化,结果表明:(1)0—6,6—12,12—24和24—48h四个脱水时段之间、0—0.5,0.5—1,1—1.5和1.5—2h四个复水时段之间电导率的差异并不显著(p0.01),说明在脱水、复水过程中,刺叶赤藓叶片质膜结构保持稳定;(2)脱水时叶片皱缩,紧贴于茎,有效避免了水分散失;复水后的叶片可借助粗壮中肋的支持和导水作用迅速展开;(3)叶片相对含水量在脱水6h后迅速下降,14h后维持在5%的较低水平,有利于植株快速进入休眠状态,维持膜结构的完整性;(4)脱水过程中,游离脯氨酸、可溶性糖含量均稳步提高,二者之间呈显著的线性正相关(R=0.952,p0.01);二者均与叶片相对含水量呈显著的线性负相关(R分别达-0.946和-0.961,p均0.01).前者可能通过和磷脂的相互作用有效稳定膜结构,后者可能通过维持小分子内部或它们之间的氢键来稳定膜系统或蛋白质的结构. |
| [50] | Biological soil crusts are receiving increasing attention due to their potentially positive effects on desert ecosystems. Artificial propagation of crusts is one way to restore degraded desert soils. However, little is known about their effectiveness on overland flow or water infiltration. Consequently, a study was done on soils of the Loess Plateau of China that had crusts propagated on different slopes. The results show that: (1) it is feasible to artificially grow moss-dominated biological soil crusts (moss crusts) in laboratory; when inoculated by sprinkling crushed fragments of stems and leaves of natural moss crusts they will cover the soil surface almost completely after about 10 months; (2) the artificially propagated moss crusts significantly increase infiltration consequently decreasing overland flow and this increase in infiltration is positively correlated to the soil coverage extent; (3) overland flow results increased by slope in moss-protected and bare soils, consequently the action of moss crusts is particularly important for protecting steep slopes; (4) the start time of runoff process is delayed by moss crusts, and the soil water redistribution process is also greatly affected. These results may be useful for helping to control desertification on the Loess Plateau of China or similar regions. |
| [51] | As part of global climate change, variation in precipitation in arid ecosystems is leading to plant adaptation in water-use strategies; significant interspecific differences in response will change the plant composition of desert communities. This integrated study on the ecophysiological and individual morphological scale investigated the response, acclimation and adaptation of two desert shrubs, with different water-use strategies, to variations in water conditions. The experiments were carried out on two native dominant desert shrubs, Tamarix ramosissima and Haloxylon ammodendron, under three precipitation treatments (natural, double and no precipitation, respectively), in their original habitats on the southern periphery of Gurbantonggut Desert, Central Asia, during the growing season in 2005. Changes in photosynthesis, transpiration, leaf water potential, water-use efficiency, above-ground biomass accumulation and root distribution of the two species were examined and compared under the contrasting precipitation treatments. There were significant interspecific differences in water-use strategy and maintenance of photosynthesis under variation in precipitation. For the phreatophyte T. ramosissima, physiological activity and biomass accumulation rely on the stable groundwater, which shields it from fluctuation in the water status of the upper soil layers caused by precipitation. For the non-phreatophyte H. ammodendron, efficient morphological adjustment, combined with strong stomatal control, contributes to its acclimation to variation in precipitation. On account of its positive responses to increased precipitation, H. ammodendron is predicted to succeed in interspecific competition in a future, moister habitat. |
| [52] | , . 本文通过调查荒漠苔藓刺叶墙藓组织培养影响因素,探讨了荒漠苔藓及其生物结皮室内快速繁殖可能性,为大规模培养荒漠苔藓以及生物结皮提供基础研究数据;通过调查含水量、叶绿素荧光、电导率、色素含量、超微结构以及再生潜力等传统指标,评价刺叶墙藓对不同脱水速度、不同温度在形态学、生理学以及繁殖生物学等方面的响应,对荒漠地区苔藓生物结皮合理保护与修复具有指导意义;将扫描电镜(SEM)、显微红外(Micro-FTIR)及单次反射(ATR)技术、顺磁共振自选标记(EPR Spin Label)、气质连用(GC/MS)、液质连用(LC/MS)、离子色谱(IC)热动力测定工具(TGA和DSC)、飞行时间质谱(MALDI-TOF)等新技术运用在苔藓耐性研究方面,摸索了刺叶墙藓在表层结构与蜡质化学、细胞内化学成分与凝胶态-液晶态-玻璃态相互转变、细胞膜透性与细胞质分子流动性以及细胞膜界面结构等对不同脱水速度和温度响应,是苔藓生物学研究上新突破,更为研究相对低等且形态较小生物体开辟了一条新途径。同时,运用双向电泳技术,比较了脱水-复水过程中刺叶墙藓蛋白质表达差异,筛选与刺叶墙藓耐干相关功能蛋白,为荒漠苔藓耐干热基因分离及其功能基因在农作物改良中应用奠定基础。 首先按照组织培养方法调查了几种常见影响因素如消毒方法、培养基、激素、培养温度、外植体、湿度及光照强度与时间等对刺叶墙藓再生的影响。最终确定刺叶墙藓快繁路线为:外植体离体叶片→温度25/15℃→营养源Knop→RH 65-80%→光周期16H→光照强度85-120 PPFD→培养周期2个月,材料无需经过消毒处理;在掌握刺叶墙藓生长影响因素后,尝试构建苔藓结皮:用离体绿叶在20/10℃诱导原丝体,继而转入河砂中,得到原始苔藓结皮,表明苔藓结皮存在人工培育可能。 为了了解荒漠苔藓对沙漠常见水分和温度胁迫的抗性机制,研究了脱水和高温及联合处理下,刺叶墙藓在形态结构、生理和繁殖特征等方面的响应。形态上刺叶墙藓具有明显的生态优势:①双层细胞壁和脂肪粒含量丰富,其中老化组织比新生组织更具有耐性优势;②快速脱水处理后不同发育阶段的叶片细胞叶绿体变圆,没有出现细胞壁和细胞膜破损;③45℃热处理仅引起叶绿体结构疏松;④45℃干热处理引起叶绿体产生大量气泡且结构扭曲,部分细胞膜破损,表明45℃下脱水和温度对细胞膜存在协同效应;⑤快速脱水、45℃热处理、45℃干热处理后的叶片细胞复水后叶绿体能够恢复正常形态。生理上刺叶墙藓具有明显的抗干热响应:①几种胁迫都引起细胞溶质大量渗漏;②色素和可溶性糖含量仅对热和干热两种处理响应剧烈;③几种胁迫叶绿素荧光活性均降低,但脱水和干热处理后叶片光系统能够快速恢复至正常水平,高温处理(≥50℃)荧光活性丧失,表明极端高温对光系统有致命影响;④高温降低刺叶墙藓再生能力,但干热联合胁迫(60℃)没有对再生能力产生显著影响;⑤脱水和高温联合处理对生理影响存在两种生态效应,即温度≤45℃二者存在协同作用,温度45℃脱水对高温效应存在拮抗作用。 为了进一步阐明刺叶墙藓耐干和耐热根本原因,从结构功能和结构化学方面对其进行了探讨。刺叶墙藓外被蜡纸层,有细胞壁瘤状凸起密布在表面,可以减少水分丧失和阻碍热空气影响,而且蜡质含量高达2.64mg g-1干重,90%以上为长链脂肪酸和烷烃,表明其细胞最外层具有明显生态优势。运用显微红外特征吸收峰揭示了刺叶墙藓化学成分变化与耐热机理存在相关关系。幼嫩组织增加亲水区脂肪和蛋白质含量、提高α-螺旋结构含量,使得细胞结构能够高温下稳定;老化组织通过提高碳氢化合物和疏水区内脂肪含量、稳定蛋白质结构等来维持细胞结构稳定,类似于糖类和蛋白质在脱水过程保护作用。运用顺磁共振自选标记方法研究了细胞膜、细胞质在脱水和干热过程中结构维持特性,刺叶墙藓细胞质稳定主要依靠高含量糖类和生物大分子,在脱水短期内实现玻璃化转变,钝化生理代谢和分子反应活性;依靠具有抗氧化功能两性分子参与膜内,减少或者消除氧自由基对细胞膜损伤,从而保证将脱水时细胞结构损伤降低在可修复范围。 以上结果表明,刺叶墙藓耐干热除了形态学优势以外,细胞质玻璃化转变在脱水过程对细胞结构具有重要作用。那么再复水过程必然引起大规模细胞溶质渗漏,因此复水后启动损伤修复机制对于刺叶墙藓非常重要。先前研究过程已经证明复水后大量蛋白表达对于修复脱水以及复水造成细胞损伤至关重要。运用双向电泳技术,初步探讨了刺叶墙藓蛋白组学响应,结果发现再复水后新表达或者表达量增加蛋白有脱水素(13.1kDa)、复水蛋白(67.5 kDa)、热激蛋白(71.4 kDa和71.2 kDa)以及小分子热激蛋白(36.7 kDa),其中脱水素和复水蛋白已经在其它生物中证明与耐干性有关,热激蛋白已经证明与耐热和耐干有关,因此这些蛋白可能与刺叶墙藓修复机制相关。 |
| [53] | 采用改进的土壤水稳性团聚体数 量测定方法,研究了黄土丘陵区不同组成和生物量的生物结皮层水稳定性.结果表明:生物结皮层的水稳定性与其生物组成有关,苔藓结皮的水稳定性显著高于藻结 皮,震荡390次后,苔藓结皮的厚度和质量损失率仅分别是藻结皮损失率的47.3%和40.1%;生物结皮层水稳定性与生物结皮的生物组成有关,60%以 上苔藓覆盖度的生物结皮的稳定性最高,质量和厚度损失率分别是藻结皮(无苔藓覆盖)损失率的28.6%和22.7%;生物结皮层水平方向结构水稳定性显著 大于垂直方向,震荡390次后,苔藓结皮的面积损失率仅为厚度损失率的6.4%.试验分析证实,生物结皮层是一种水平方向稳定性极强的层状结构体,这一结 构特性增强了其抗风蚀和水蚀的能力. |
| [54] | . . 藓类植物形态结构简单,容易受环境影响,常用来做为环境变化的指示植物。研究藓类植物适应环境的形态结构和生理特性,一方面可以增加对藓类植物适应环境机制的了解,另外一方面也可以为评估环境变化对藓类植物的影响,筛选对环境敏感的生物指示植物提供基础资料。我们以不同水分条件下的的藓类植物为材料,对它们的茎、叶进行了形态解剖研究,并以多蒴灰藓(Hypnum fertile Senden.)为对象,在实验控制条件下,测定了干旱、温度、酸雨、氮沉降、紫外辐射等环境因子对其生理指标影响。主要内容、结果和结论如下: 1.使用石蜡切片法,对采自不同生境下的30种藓类植物茎的横切面进行了解剖比较。发现30种藓类植物茎的形状、细胞形状、表皮层数及厚度、皮部是否分化及细胞层数、中轴有无形状及所占比例等特征因种类不同而差异明显。分析发现,藓类植物茎的表皮及外皮部细胞层数、细胞壁厚度与生境的相关性较大。 2.使用石蜡切片法,对采自不同生境下的17种藓类植物的叶片进行解剖观察和比较,发现不同的藓类植物在中肋导水主细胞的有无、厚壁细胞是否分化、中肋细胞层数及细胞密度、叶片细胞层数、叶表附属物、叶片细胞密度等方面存在明显差异。分析发现,藓类植物在叶片细胞胞壁厚薄、叶片附属结构、中肋结构等方面表现出对水分条件的适应,而细胞密度与水分条件相关性不大。 3.以多蒴灰藓为材料,用聚乙二醇(PEG-6000)模拟干旱胁迫处理,强度为-5~50 Bar PEG溶液,胁迫处理3天。实验处理后,多朔灰藓叶绿素含量和丙二醛(MDA)含量没有明显变化,可溶性糖含量在胁迫程度大于-25Bar时与对照相比具有明显增加,在胁迫强度大于-15 Bar时,与对照相比,多蒴灰藓的脯氨酸含量明显增加,其超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性明显增强。多蒴灰藓通过增加体内渗透调节物质含量、提高抗氧化酶活性免受干旱胁迫的伤害。 4.5℃低温处理下,多朔灰藓可溶糖、脯氨酸、丙二醛含量以及POD、CAT活性都逐渐升高,SOD活性下降;40℃高温处理下,可溶性糖含量先升高后降低,脯氨酸含量逐渐升高,SOD、POD、CAT活性在高温下先升高后降低,丙二醛含量逐渐升高且增加幅度大于5℃;20℃培养下的多蒴灰藓具有相对较高的酶活性。研究表明多朔灰藓能通过增加渗透调节物质含量和提高保护酶活性抵御温度胁迫;与高温相比,多蒴灰藓更适合生长于低温环境。 5.用喷施法研究模拟酸雨对多朔灰藓影响。结果表明,随着处理液的pH降低,多蒴灰藓的叶绿素、可溶性糖含量随pH值而降低,脯氨酸、丙二醛含量和质膜透性升高,SOD、CAT活性先升高然后缓慢下降,再到迅速下降,POD活性先缓慢上升然后快速上升。用差异显著性检验分析表明,模拟酸雨对多蒴灰藓的影响阈值为pH 4.0。 6.室内模拟环境UV-B辐射(6.93 KJ m~(-2)·d~(-1))和增强的UV-B辐射(9.01KJ m~(-2)·d~(-1),高30%),研究了紫外线辐射对多朔灰藓的影响。结果表明三周内增强的UV-B辐射导致多蒴灰藓配子体叶绿素和类胡萝卜素含量渐少,而MDA含量增加,SOD、POD和CAT的活性下降。说明UV-B辐射的增强对多蒴灰藓产生氧化胁迫。 7.用喷施法,研究了不同浓度、不同pH的硝酸溶液和铵盐溶液对多朔灰藓和东亚小金发藓(Pogonatum inflexum(Lindb.)Lac.)硝酸还原酶活性的短期影响。结果表明硝酸盐诱导硝酸还原酶活性逐渐升高,多朔灰藓酶活性增加速率大于东亚小金发藓。铵盐则抑制硝酸还原酶活性;pH3的溶液能显著降低多朔灰藓硝酸还原酶活性,对东亚小金发藓没有明显影响。与东亚小金发藓相比,多朔灰藓对N沉降更敏感。 |
| [55] | 利用快速叶绿素荧光动力学技术研究了毛尖紫萼藓脱水和复水过程中叶绿素荧 光变化,结果显示在脱水过程中毛尖紫萼藓PSII的最大光化学效率(Fv/Fm)、光合机构电子传递的量子产额(ETo/ABS)、捕获的激子将电子传递 到电子传递链中超过QA的其它电子受体的概率(ETo/TRo)、单位叶面积的反应中心的数量(RC/CSo) 以及PSⅡ受体库(Area) 对叶片含水量的响应等均存在相对含水量阈值.在阈值范围内脱水,对以上荧光参数影响不大,低于阈值后,各荧光参数值迅速下降,直至PSII反应中心完全关 闭以及光化学过程结束.再复水后,毛尖紫萼藓光合机构的最大捕光效率、实际光化学效率、PSII反应中心受体侧的电子传递链以及反应中心均能得到快速而有 效的恢复.表明一定时间内脱水不会对毛尖紫萼藓的光合器官造成严重伤害,光合系统仍维持在可恢复状态. |
| [56] | |
| [57] | |
| [58] | 新疆古尔班通古特沙漠地表具有发育完好的生物土壤结皮,本文以参与组成生物土壤结皮的优势藓类———齿肋赤藓(Syntrichia caninervis)为研究对象,在实验室内对其进行模拟降雨处理,降雨量分别为0mm(对照)、2mm、6mm、10mm和15mm,研究模拟降雨影响下齿肋赤藓植株含水量、光合色素含量及可溶性糖、游离脯氨酸、可溶性蛋白、抗氧化酶(SOD、CAT和POD)、丙二醛含量的变化,以探讨其对降雨的生理生化响应。结果表明:随降雨量的增加,齿肋赤藓植株含水量显著增加;随齿肋赤藓植株含水量的增加,其光合色素含量和可溶性蛋白含量显著增加;随降雨量增加,抗氧化酶(SOD、CAT和POD)活性和可溶性糖含量呈大致递减的趋势,游离脯氨酸和丙二醛含量呈先下降后上升的变化趋势。在干燥状态下齿肋赤藓通过提高体内保护酶(SOD、CAT和POD)活性,增加可溶性糖和游离脯氨酸含量来抵御外界干旱胁迫的环境;而降雨能够增加齿肋赤藓植株含水量,激活其光合生理活性,显著提高光合色素含量,但是降雨量过大又将加剧齿肋赤藓膜脂过氧化的程度。 |
| [59] | |
| [60] | Biological soil crusts (BSCs) play an important role in the early succession of vegetation restoration in the Loess Plateau, China. To evaluate the effects of artificially cultivated BSCs on the soil surface micro-environment, we obtained natural moss crusts and moss-lichen crusts from the Loess Plateau of Shaanxi province, and subsequently inoculated and cultivated on horizontal and sloping surfaces of loess soil in a greenhouse. The chemical and biological properties of the subsoil under cultivated BSCs were determined after 10 weeks of cultivation. The results indicated that BSCs coverage was more than 65% after 10 weeks of cultivation. Moss crust coverage reached 40% after 5 weeks of cultivation. Compared with the control, soil organic matter and available nitrogen contents in moss crust with the horizontal treatments increased by 100.87% and 48.23%, respectively; increased by 67.56% and 52.17% with the sloping treatments, respectively; they also increased in moss-lichen crust with horizontal and sloping treatments, but there was no significant difference. Available phosphorus in cultivated BSCs was reduced, soil pH was lower and cationic exchange capacity was higher in cultivated BSCs than in the control. Alkaline phosphatase, urease and invertase activities were increased in artificially cultivated BSCs, and alkaline phosphatase activity in all cultivated BSCs was obviously higher than that in the control. Numbers of soil bacteria, fungi and actinomycetes were increased in the formation process of cultivated BSCs. These results indicate that BSCs could be formed rapidly in short-term cultivation and improve the micro-environment of soil surface, which provides a scientific reference for vegetation restoration and ecological reconstruction in the Loess Plateau, China. |
| [61] | 生物结皮是具有生命活性的地表覆被物,在黄土丘陵半干旱区广泛发 育.迄今有关该区不同类型生物结皮的光合、呼吸作用潜力及其影响因素还不清楚.通过野外调查采样,利用室内半开放光合测定系统,测定了黄土丘陵区典型的藻 类(3年生)、藓类(土生扭口藓)生物结皮(13年生)光合及呼吸作用过程对水分、温度及光照等环境因子的响应,在此基础上,解释了不同类型生物结皮空间 分布格局的内在原因.结果表明:(1)在适合的光温水条件下,生物结皮的光合及呼吸作用强度取决于生物组成,研究区藓结皮的光合及呼吸作用速率显著高于藻 结皮;(2)水分是生物结皮光合、呼吸作用的关键影响因子,只有当水分含量达到一定水平,才有光合及呼吸的可能.黄土丘陵区藓类及藻类生物结皮光合作用最 适水分条件分别为80%-100%和40%-80%田间持水量.(3)藓类及藻类生物结皮光合作用光补偿点PAR均低于10μmol·m-2·s-1,光 饱和点PAR分别为1000和800μmol·m-2·s-1,研究中2类生物结皮均未出现光抑制现象.(4)温度显著影响生物结皮的光合及呼吸作用,研 究区藓类及藻类生物结皮的最适光合温度分别为20-25℃和25-30℃.(5)生物结皮光合作用对含水量及温度的响应可以解释结皮的空间分布特征.藻类 植物光合作用的最适含水量较低而最适温度较高,因此在失水较快的阳坡及开阔地面上依然可以大面积发育;而藓类植物光合作用对水分条件要求较高,因此在水分 条件较好的阴坡、低洼地才易演替至藓结皮阶段. |
| [62] | 在实验室内通过光镜与扫描电镜相结合的手段观察了新疆古尔班通古特沙漠藓类结皮层的优势物种——齿肋赤藓(Syntrichia caninervis Mitt.)的形态结构特征及其原丝体发育的全过程,并分别从藓类植物生长发育所必经的配子体和原丝体两个阶段对齿肋赤藓形态结构与荒漠干旱的生态环境之间的适应性关系进行了探讨。研究结果表明,①植物体密集丛生;叶片边缘强烈背卷,两面均具马蹄形细疣;中肋粗壮并突出叶尖成白色长毛尖。②原丝体系统的细胞短粗,细胞质高度浓缩,液泡较小,细胞壁较厚;原丝体系统具有短枝群。 |
1
2004
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
苔藓植物光合色素含量测定方法——以暖地大叶藓为例.
1
2005
... 光合色素含量的测定参照包维楷
ABA treatment increases both the desiccation tolerance of photosynthesis, and nonphotochemical quenching in the moss Atrichum undulatum.
2000
Surface disturbances: Their role in accelerating desertification.
1
1995
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
Plant size, sex expression and sexual reproduction along an elevation gradient in a desert moss.
1
2011
... 水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(
Effects of long-term grazing on cryptogam crust cover in Navajo National Monument, Ariz.
1
1983
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
Rain and rodents: Complex dynamics of desert consumers although water is the primary limiting resource in desert ecosystems, the relationship between rodent population dynamics and precipitations complex and nonlinear.
1
2002
... 水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(
Key factors influencing rapid development of potentially dune-stabilizing moss- dominated crusts.
2015
苔藓植物的生物多样性及其保护.
1
1997
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
藓类植物的无性繁殖及其应用.
1
2008
... 苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主.藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(
Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation.
1
2005
... 当植物受到逆境胁迫时, 植物细胞内活性氧自由基产生和清除的平衡遭到破坏, 使活性氧大量富集, 它们对植物细胞具有损伤作用, 而植物细胞的抗氧化酶系统(CAT、POD、SOD)在消除或降低氧自由基带来的损伤中发挥着重要作用.CAT主要清除过氧化氢自由基, POD主要清除过氧化物自由基, SOD主要清除超氧自由基(
脱水和复水对复苏植物牛耳草(Boea hygrometrica)离体叶片光合作用的影响.
1
2000
... 光合色素反应植物对环境中光资源的利用情况, 其中叶绿素在光合作用中起核心作用, 其含量与光合速率紧密相关, 既可反映植物叶片光合作用功能的强弱, 也可用以表征逆境胁迫下植物组织器官的受损状况(
快速脱水与复水过程中2种藓类植物的活性氧代谢与脂质过氧化特征.
2012
In vitro cultivation of bryophytes: A review of practicalities, problems, progress and promise.
1
2004
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
Exploring some relationships between biological soil crusts, soil aggregation and wind erosion.
1
2003
... 苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主.藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(
The influence of air pollution on soluble proteins, chlorophyll degradation, MDA, sulphur and heavy metals in a transplanted lichen.
1
1994
... 可溶性糖、可溶性蛋白和脯氨酸含量分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定(
不同早熟禾品种对干旱胁迫的生理响应及抗旱性评价.
1
2014
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding.
1
2004
... 当植物受到逆境胁迫时, 植物细胞内活性氧自由基产生和清除的平衡遭到破坏, 使活性氧大量富集, 它们对植物细胞具有损伤作用, 而植物细胞的抗氧化酶系统(CAT、POD、SOD)在消除或降低氧自由基带来的损伤中发挥着重要作用.CAT主要清除过氧化氢自由基, POD主要清除过氧化物自由基, SOD主要清除超氧自由基(
藓类结皮层人工培养试验和维持机制研究.
1
2012
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
干旱对苜蓿叶片可溶性蛋白的影响.
1
2005
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Microbiotic crusts as biomarkers for surface stability and wetness duration in the Negev Desert.
1
2009
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
Properties and spatial distribution of microbiotic crusts in the Negev Desert.
1
2010
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
Analysis of environmental factors determining development and succession in biological soil crusts.
1
2015
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
模拟水淹对中华蚊母树生长及光合特性的影响.
1
2014
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
三峡库区岸生植物秋华柳对水淹的光合和生长响应. 植物生态学报,
1
2007
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
Chlorophyll fluorescence—— A practical guide.
1
2000
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress.
1
2003
... 当植物受到逆境胁迫时, 植物细胞内活性氧自由基产生和清除的平衡遭到破坏, 使活性氧大量富集, 它们对植物细胞具有损伤作用, 而植物细胞的抗氧化酶系统(CAT、POD、SOD)在消除或降低氧自由基带来的损伤中发挥着重要作用.CAT主要清除过氧化氢自由基, POD主要清除过氧化物自由基, SOD主要清除超氧自由基(
Leaf gas exchange, chlorophyll fluorescence and growth responses of Genipa americana seedlings to soil flooding.
1
2003
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
Bryophytes as experimental models for the study of environmental stress tolerance: Tortula ruralis and desiccation-tolerance in mosses.
1
2000
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
Desiccation-induced physiological and biochemical changes in resurrection plant, Selaginella bryopteris.
1
2010
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Experiments on the effect of different intensities of desiccation on bryophyte survival, using chlorophyll fluorescence as an index of recovery.
1
2003
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
Changes to dryland rainfall result in rapid moss mortality and altered soil fertility.
1
2012
... 光合色素反应植物对环境中光资源的利用情况, 其中叶绿素在光合作用中起核心作用, 其含量与光合速率紧密相关, 既可反映植物叶片光合作用功能的强弱, 也可用以表征逆境胁迫下植物组织器官的受损状况(
Short distances between extreme microhabitats do not result in ecotypes in Syntrichia caninervis.
1
2011
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
Balancing the generation and elimination of reactive oxygen species.
1
2005
... 光合色素反应植物对环境中光资源的利用情况, 其中叶绿素在光合作用中起核心作用, 其含量与光合速率紧密相关, 既可反映植物叶片光合作用功能的强弱, 也可用以表征逆境胁迫下植物组织器官的受损状况(
逐渐干旱胁迫下生物土壤结皮中真藓和土生对齿藓的活性氧清除机制.
1
2012
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Proline accumulation and varietal adaptability to drought in barley: A potential metabolic measure of drought resistance.
1
1972
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Effects of increased summer precipitation and N deposition on Mojave Desert populations of the biological crust moss Syntrichia caninervis.
1
2011
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
Gender-specific heat-shock tolerance of hydrated leaves in the desert moss Syntrichia caninervis.
1
2006
... 苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主.藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(
Response mechanisms of antioxidants in bryophyte (Hypnum plumaeforme) under the stress of single or combined Pb and/or Ni.
1
2009
... 可溶性糖、可溶性蛋白和脯氨酸含量分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定(
叶片毛尖对齿肋赤藓结皮凝结水形成及蒸发的影响.
1
2012
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
a). 腾格里沙漠固定沙丘藓类植物结皮层的自然恢复及人工培养试验研究.
2
2005
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... ).野外藓类结皮恢复耗时较长, 同时两种条件培养下的藓类结皮后期均出现衰退现象(
b). 固定沙丘生物结皮层藓类植物形态结构及其适应性研究.
1
2005
... 苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主.藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(
古尔班通古特沙漠不同生活型植物对小雨量降雨的响应.
1
2009
... 沙漠水分来源主要依靠降水, 而降水的分配具有明显的季节差异, 如古尔班通古特沙漠降雨多集中在早春夏末, 早春融雪及夏季降雨后土壤水分短暂时间内处于一个过饱和的状态且面临一定的水分胁迫, 其他季节干旱.研究区降水主要以0-5 mm的小雨量为主, 占该沙漠总降雨频次的89.8%, 平均降雨间隔在6天左右(
脱水对齿肋赤藓叶片亚显微结构及生理生化指标的影响
1
2009
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
生物结皮中齿肋赤藓叶片细胞显微与亚显微结构特征.
1
2009
... 光合色素反应植物对环境中光资源的利用情况, 其中叶绿素在光合作用中起核心作用, 其含量与光合速率紧密相关, 既可反映植物叶片光合作用功能的强弱, 也可用以表征逆境胁迫下植物组织器官的受损状况(
Bryophytes as model systems.
1
2000
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
不同冬小麦品种旗叶叶绿素荧光特性及其对干旱胁迫的响应.
3
2015
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... 沙漠水分来源主要依靠降水, 而降水的分配具有明显的季节差异, 如古尔班通古特沙漠降雨多集中在早春夏末, 早春融雪及夏季降雨后土壤水分短暂时间内处于一个过饱和的状态且面临一定的水分胁迫, 其他季节干旱.研究区降水主要以0-5 mm的小雨量为主, 占该沙漠总降雨频次的89.8%, 平均降雨间隔在6天左右(
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
Precipitation intensity is the primary driver of moss crust-derived CO2 exchange: Implications for soil C balance in a temperate desert of northwestern China.
2015
生物土壤结皮中刺叶赤藓质膜透性对脱水复水过程的响应.
1
2009
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Artificial culture of biological soil crusts and its effects on overland flow and infiltration under simulated rainfall.
1
2011
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation.
1
2007
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
典型荒漠苔藓人工繁殖特征与抗御干热环境胁迫的生理生化机制研究
1
2007
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
黄土丘陵区生物土壤结皮层水稳性.
1
2012
... 苔藓结皮是荒漠生物结皮发育的最高阶段, 多以藓类植物为主.藓类植物虽没有完善的维管束和真正的根, 但通过大量的假根将植物体与土壤颗粒紧密连接捆绑在一起形成藓类结皮, 有效地减少了土壤表面的风蚀和水蚀(
藓类植物适应环境的形态结构及生理学机制
2008
毛尖紫萼藓(Grimmia pilifera P. Beauv) PSII光化学效率对脱水和复水的响应.
2007
冻融过程对荒漠区不同微生境下齿肋赤藓渗透调节物含量和抗氧化酶活力的影响.
3
2015
... 可溶性糖、可溶性蛋白和脯氨酸含量分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定(
... ;
... 水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(
冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响.
2
2011
... 处理48天后, 利用便携式调制叶绿素荧光仪(PAM 2500, Walz, Heinz, Germany)进行人工培养齿肋赤藓叶绿素荧光特性的测定.测定前先对样品进行30 min充分暗适应后, 每个培养皿内随机选取3株藓株于波长650 nm、强度为0.1 μmol·m-2·s-1的红光下测得初始荧光(F0), 最大荧光(Fm)于强度为 10β000 μmol·m-2·s-1的饱和脉冲光下测得, 脉冲时间为0.8 s.在光照强度为250 μmol·m-2·s-1的光照下测定稳态荧光(Ft), 待Ft稳定后, 打开一次饱和脉冲光测得光适应后的最大荧光(Fm°), 样品重复数为5个.根据以上参数可以计算光系统II (PSII)的最大光化学效率(Fv/Fm) = (Fm - F0)/Fm和光适应下PSII的实际光化学效率(ΦPSII) = Fm° - Ft/Fm° (
... 光合生理是植物最重要的生理过程, 而叶绿素荧光特性能从内在揭示植物光合生理的表现, 其中荧光参数Fv/Fm及ΦPSII能够反映出PSII光能转化效率, 其变化可以直接体现植物受逆境胁迫的状况(
模拟降雨对齿肋赤藓(Syntrichiacaninervis)生理特性的影响.
3
2014
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... 水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(
... 当植物受到逆境胁迫时, 植物细胞内活性氧自由基产生和清除的平衡遭到破坏, 使活性氧大量富集, 它们对植物细胞具有损伤作用, 而植物细胞的抗氧化酶系统(CAT、POD、SOD)在消除或降低氧自由基带来的损伤中发挥着重要作用.CAT主要清除过氧化氢自由基, POD主要清除过氧化物自由基, SOD主要清除超氧自由基(
高温和干旱对鳞叶藓游离脯氨酸和可溶性糖含量的影响.
2
2004
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... 渗透调节是植物适应水分胁迫的一种重要机制, 包括脯氨酸、可溶性糖、可溶性蛋白等, 在植物遇到逆境胁迫时能够维持一定的细胞膨压, 从而维持植物细胞持续生长、气孔开放和光合作用等生理过程的进行.因此, 植物体内渗透调节物质含量的变化与植物受环境的胁迫程度紧密相关, 同时也能反映植物的抗逆特性(
Effects of artificially cultivated biological soil crusts on soil nutrients and biological activities in the Loess Plateau.
2
2014
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... 沙漠水分来源主要依靠降水, 而降水的分配具有明显的季节差异, 如古尔班通古特沙漠降雨多集中在早春夏末, 早春融雪及夏季降雨后土壤水分短暂时间内处于一个过饱和的状态且面临一定的水分胁迫, 其他季节干旱.研究区降水主要以0-5 mm的小雨量为主, 占该沙漠总降雨频次的89.8%, 平均降雨间隔在6天左右(
生物结皮光合作用对光温水的响应及其对结皮空间分布格局的解译——以黄土丘陵区为例.
1
2010
... MDA作为细胞膜发生膜脂过氧化作用的产物, 其浓度的高低表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱(
荒漠藓类结皮层中齿肋赤藓形态结构适应性及其原丝体发育特征.
2
2009
... 自然条件下藓类结皮的形成需要几年甚至数十年的时间(
... 水分是荒漠生态系统植物生长的主要限制因子, 作为典型的变水植物, 齿肋赤藓植株无真正的根、茎、叶结构, 植株含水量极易随外界环境的变化而变化(


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}