Metabolic responses of wheat roots to alkaline stress
GUORui1,2,*,, ZHOUJi3, YANGFan4, LIFeng1 1Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081, China2Key Laboratory of Dryland Agriculture, Ministry of Agriculture, Beijing 100081, China3Land Consolidation and Rehabilitation Centre, the Ministry of Land and Resources, Beijing 100034, Chinaand 4Jilin Academy of Forestry Science, Changchun 130033, China 通讯作者:*E-mail:guorui01@caas.cn 收稿日期:2017-04-5 接受日期:2016-04-14 网络出版日期:2017-07-28 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 国家自然科学基金(31200243和31570328)
关键词:小麦;碱胁迫;根系;生长特性;代谢调控 Abstract Aims The aim of this study was to investigate the effects of alkaline stress on primary, secondary metabolites and metabolic pathways in the roots of wheat (Triticum aestivum). The results were used to evaluate the physiological adaptive mechanisms by which wheat tolerated alkali stress.Methods A pot experiment was carried out in the greenhouse. For each plastic pot, five wheat seeds were planted. After germination, seedlings were allowed to grow under controlled water and nutrient conditions for two months, then seedlings were exposed to alkaline stress (NaHCO3-Na2CO3) for 12 days. The relative growth rate (RGR), absolute water content (AWC), metal elements, free cations and metabolites were measured.Important findings The alkaline stress caused the reduction of RGR and AWC. Alkaline stress caused a rapid increase of Na content with the concurrent decrease in K and Cl content, resulting in inhibited metal element accumulation and an ionic imbalance. In the present study, alkaline stress strongly enhanced Ca accumulation in wheat roots, suggesting that an increased Ca concentration can immediately trigger the salt overly sensitive (SOS)-Na exclusion system and reduce Na-associated injuries. Also, 70 metabolites, including organic acids, amino acids, sugars/polyols and others, behaved differently in the alkaline stress treatments according to a GC-MS analysis. The metabolic profiles of wheat were closely associated with alkaline-stress conditions. Alkaline stress caused the accumulation of organic acids, accompanied by the depletion of sugars/polyols and amino acids. Organic acids could play a central role in the regulation of intracellular pH by accumulating vacuoles to neutralize excess cations. Glycolysis and amino acid synthesis in roots were inhibited under salt stress while prolonged alkaline stress led to a progressive tricarboxylic acid (TCA) cycle. The severe negative effects of alkaline stress on sugar synthesis and storage may reflect the toxic levels of Na+ accumulating in plant cells in a high-pH environment, implying that the reactive oxygen species detoxification capacity was diminished by the high pH. A lack of NO3- in wheat roots can decrease synthase enzyme activities, limiting the synthesis of amino acids. Under salt stress, the TCA cycle and organic acid accumulation increased, but glycolysis and amino acid synthesis were inhibited in roots. Thus, energy levels and high concentrations of organic acids may be the key adaptive mechanisms by which wheat seedlings maintain their intracellular ion balance under alkaline stress.
碱胁迫下根系RGR随盐浓度增加而下降, 且下降幅度随盐浓度增加而加强(图1A, p < 0.05)。根系AWC变化趋势和RGR基本相同, 胁迫强度越高下降越显著(图1B, p < 0.05)。 显示原图|下载原图ZIP|生成PPT 图1碱浓度对小麦根系相对生长率(RGR)和绝对含水量(AWC)的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。 -->Fig. 1Effects of alkaline concentration on root relative growth rate (RGR) and absolute water content (AWC) (mean ± SE, n = 5). Different lowercase letters indicate significant differences among the various treatments (p < 0.05). -->
2.3 碱胁迫对小麦矿质元素吸收和游离阴离子的影响
如表2所示, 在碱胁迫12天后小麦根系中矿质元素含量发生了显著变化。与对照相比较, 碱胁迫下小麦根系中Na元素含量随盐度增加呈现上升趋势, 但K元素含量呈现下降趋势, 且随着盐度的增加变化幅度加强(表2, p < 0.05)。当盐度为50 mmol·L-1时根系中Ca含量缓慢增加, 但盐度达到100 mmol·L-1后Ca含量急剧增加(表2, p < 0.05)。根系中Mg、Cu、Fe、Zn元素对碱胁迫的响应相似, 都是随着盐度的增加, 含量呈现下降趋势, 但在中低碱胁迫下Cu和Fe变化不明显(表2, p < 0.05)。另外, 碱胁迫对Mn元素的影响不明显(表2, p < 0.05)。碱胁迫下小麦根系Cl-和NO3-含量均下降, 且随着盐度增加下降幅度增大(表3, p < 0.05)。碱胁迫下根系中H2PO4-含量均低于对照处理组(表3, p < 0.05)。而碱胁迫对根系中SO42-含量影响不大(表3, p < 0.05)。 Table 2 表2 表2碱胁迫对小麦根系干物质中矿质元素含量的影响(平均值±标准误差, n = 5) Table 2Effects of alkaline stress on the contents of metal elements in the roots of wheat seedlings (mean ± SE, n = 5)
处理 Treatment
碱胁迫浓度 Alkaline concentration (mmol)
矿质元素 Metal elements (mmol·g-1 dry mass)
Na
K
Ca
Mg
Cu
Fe
Zn
Mn
对照 Control
0
4.25 ± 1.01b
69.90 ± 3.03a
17.11 ± 1.01b
7.80 ± 0.29a
1.25 ± 0.04a
1.56 ± 0.08a
0.07 ± 0.01a
0.06 ± 0.00a
碱胁迫 Alkaline stress
50
15.89 ± 2.06 ab
45.20 ± 1.54b
20.53 ± 1.00b
6.01 ± 0.16b
1.02 ± 0.02a
1.26 ± 0.03a
0.05 ± 0.00b
0.05 ± 0.00a
100
19.09 ± 2.11a
16.83 ± 1.02c
32.85 ± 1.30a
4.96 ± 0.47c
0.55 ± 0.03b
0.84 ± 0.05b
0.03 ± 0.00c
0.06 ± 0.00a
Different lowercase letters indicate significant differences among the various treatments (p < 0.05).不同小写字母表示不同处理间差异显著(p < 0.05)。 新窗口打开 Table 3 表3 表3碱胁迫对小麦根系干物质中阴离子含量的影响(平均值±标准误差, n = 5) Table 3Effects of alkaline stress on the contents of Cl-, NO3-, H2PO4-, SO42- in the roots of wheat seedlings (mean ± SE, n = 5).
处理 Treatment
碱胁迫浓度 Alkaline concentration (mmol)
阴离子 Anions (mmol·g-1 dry mass)
Cl-
NO3-
H2PO4-
SO42-
对照 Control
0
0.12 ± 0.01a
0.41 ± 0.02a
0.05 ± 0.00a
0.03 ± 0.00a
碱胁迫 Alkaline stress
50
0.09 ± 0.00b
0.34 ± 0.01b
0.03 ± 0.01b
0.02 ± 0.00a
100
0.04 ± 0.01c
0.07 ± 0.00c
0.02 ± 0.00b
0.03 ± 0.00a
Different lowercase letters indicate significant differences among the various treatments (p < 0.05).不同小写字母表示不同处理间差异显著(p < 0.05)。 新窗口打开
2.4 碱胁迫下小麦根系代谢组变化轨迹
本研究利用GC-MS技术检测小麦根系对碱胁迫的生理响应过程, 主成分分析(PCA)的得分图显示, 3组样本——对照组(0 mmol·L-1)、中度碱胁迫(50 mmol·L-1)和重度碱胁迫(100 mmol·L-1)呈现显著性差异, 两个主成分得分为95%, 这表明本研究GC-MS的代谢组学方法可以有效地表征小麦根系碱胁迫下的生理代谢状态(图2A)。OPLS-DA结果进一步证实小麦根系对照组和碱胁迫组代谢发生显著性变化, 得分图显示对照组分别和50 mmol·L-1碱胁迫处理(建模有效性得分值(R2X) = 0.86, Q2 = 0.99)以及100 mmol·L-1碱胁迫处理(R2X = 0.91, Q2 = 0.99)都得到了很好的OPLS-DA模型(图2B、2C)。 显示原图|下载原图ZIP|生成PPT 图2小麦根系在不同碱胁迫强度下代谢变化过程的SIMCA得分图。PCA得分图 (A)。OPLS-DA得分图, 对照组vs 50 mmol·L-1碱胁迫(B), 对照组vs 100 mmol·L-1碱胁迫(C)。 -->Fig. 2SIMCA analyzed score plots showing the metabolomic trajectory of roots of wheat seedlings under different salinity concentration treatments. Principal component analysis (PCA) score plots (A). Orthogonal partial least squares discriminant analysis (OPLS-DA) scores: CK vs. AS-50 mmol·L-1 (B) and CK vs. AS-100 mmol·L-1 (C). -->
2.5 碱胁迫对小麦根系代谢产物的影响
根据主成分分析结果, 在50 mmol·L-1和100 mmol·L-1碱胁迫下小麦根系与对照相比较, 分别有33个和57个代谢物发生显著性变化, 其中相同的代谢物在高碱胁迫下变化倍数明显高于中低碱胁迫(表4, p < 0.05; p < 0.01)。如表4显示, 在三羧酸(TCA)循环中不同浓度碱胁迫均促进柠檬酸、琥珀酸和苹果酸的合成, 另外在100 mmol·L-1碱胁迫下其他参与TCA循环的代谢物都呈现极显著的上升(表4,p< 0.05; p < 0.01)。但是, 碱胁迫迫使参与糖酵解的代谢物含量降低, 尤其在高盐浓度下(表4, p < 0.05; p < 0.01)。不同浓度碱胁迫均造成参与莽草酸途径的苯丙氨酸和酪氨酸含量降低, 100 mmol·L-1浓度又使得色氨酸和肉桂酸含量降低, 但与此同时莽草酸和奎尼酸在高碱胁迫下显著增加(表4, p < 0.05; p < 0.01)。与对照相比, 碱胁迫使肌醇、甘氨酸、丝氨酸和乙醇胺含量降低, 这表明碱胁迫明显抑制细胞膜脂代谢过程, 尤其在高pH值条件下(表4, p < 0.05; p < 0.01)。碱胁迫导致氨基酸显著性减少, 另外没有引起脯氨酸的显著增加(表4, p < 0.05; p < 0.01)。碱胁迫造成小麦根系中糖类和多元醇代谢物含量降低, 包括葡萄糖、果糖、蔗糖等; 但是明显引起有机酸类物质含量显著升高, 包括柠檬酸琥珀酸乳酸等(p < 0.05; p < 0.01)。 Table 4 表4 表4不同碱胁迫下小麦根系提取物中主要代谢物相对含量和变化倍数 Table 4Relative concentration and changes of major metabolites in roots of wheat seedlings after alkaline stress treatment
代谢路径和代谢物名称 Metabolic pathways and metabolites
相对含量 Relative concentration
倍性变量Fold changes
CK
AS-50 mmol
AS-100 mmol
log2(50/CK)
log2(100/CK)
三羧酸循环 Tricarboxylic acid cycle
柠檬酸 Citric acid
75.73
150.14
231.23
0.99*
1.61**
乌头酸 Aconitic acid
1.34
1.70
5.13
0.34
1.93**
α-酮戊二酸 α-ketoglutaric acid
0.18
0.33
0.85
0.89
2.23**
琥珀酸 Succinic acid
19.05
61.45
120.69
1.69**
2.66**
延胡索酸 Fumaric acid
1.34
1.56
9.63
0.23
2.85**
苹果酸 Malic acid
10.47
23.10
30.35
1.14*
1.54**
糖酵解过程 Glycolysis
葡萄糖 Glucose
26.37
13.67
7.91
-0.95*
-1.74**
葡萄糖-6-磷酸 Fructose-6-phosphate
0.57
0.25
0.08
-1.16*
-2.75**
果糖-6-磷酸 Glucose-6-phosphate
0.22
0.13
0.04
-0.80
-2.58**
3-磷酸甘油酸 3-phosphoglyceric acid
0.50
0.34
0.11
-0.57
-2.19**
丙酮酸 Pyruvate
0.54
0.50
0.31
-0.10
-0.81
磷酸烯醇式丙酮酸 Enolphosphopyruvate
0.73
0.61
0.22
-0.26
-1.74**
莽草酸途径 Shikimic path way
莽草酸 Shikimic acid
1.84
1.22
3.84
-0.60
1.06*
奎尼酸 Quinic acid
4.17
6.17
23.11
0.58
2.47**
苯丙氨酸 Phenylalanine
0.65
0.17
0.11
-1.91*
-2.60**
色氨酸 Tryptophan
0.05
0.03
0.02
-0.71
-1.45*
酪氨酸 Tyrosine
1.59
0.03
0.02
-4.18**
-6.57**
肉桂酸 Cinnamic acid
0.25
0.15
0.12
-0.77
-1.12*
细胞膜脂代谢 Metabolism of plasma membrane
肌醇 Myo-inositol
19.17
9.63
3.07
-0.99*
-2.64**
甘氨酸 Glycine
0.74
0.34
0.12
-1.56**
-2.69**
丝氨酸 Serine
17.83
9.56
1.64
-0.90*
-3.44**
乙醇胺 Ethanolamine
20.23
11.31
3.26
-0.84*
-2.63**
氨基酸 Amino acid
γ-氨基丁酸 γ-aminobutyric acid
137.19
41.77
23.92
-1.72**
-2.52**
丙氨酸 Alanine
106.58
56.88
10.64
-0.91*
-3.32**
谷氨酸 Glutamate
24.93
13.73
8.40
-0.86*
-1.57**
天冬酰胺 Asparagine
12.65
4.38
0.91
-1.53**
-3.79**
天冬氨酸 Aspartic acid
7.60
4.06
3.34
-0.90*
-1.19*
脯氨酸 Proline
10.04
15.47
11.57
0.62
0.20
赖氨酸 Lysine
17.32
0.33
0.11
-5.72**
-7.26**
糖类及多元醇 Sugars and polyols
果糖 Fructose
814.31
574.62
173.68
-0.78*
-2.23**
蔗糖 Sucrose
136.11
92.04
65.17
-0.56
-1.06*
塔罗糖 Talose
114.17
75.07
23.13
-0.60
-2.30**
蔗果三糖 Kestose
57.63
42.47
19.69
-0.44
-1.55**
核糖 Ribose
8.24
4.25
1.38
-0.96*
-2.58**
The relative concentration of each metabolite is the average of data from five biological replicates obtained through gas chromatography-mass spectrometry. Changes were calculated using the formula log2(treatment/control). * indicates significant (p < 0.05) and ** highly significant difference (p < 0.01), respectively.每个代谢产物的相对含量都是5次生物学重复测量的平均值。变化倍数来自于公式log2(胁迫/对照), *表示显著性差别(p < 0.05); **表示极显著性差别(p < 0.01)。 新窗口打开
DengRL, XuHR, CaoYF, XiaoK (2007). The molecular basis of ammonium transporters in plants. Plant Nutrition and Fertilizer Science, 13, 512-519. (in Chinese with English abstract)[邓若磊, 徐海荣, 曹云飞, 肖凯 (2007). 植物吸收铵态氮的分子生物学基础 . 植物营养与肥料学报, 13, 512-519.] [本文引用: 1]
GuoR, LiF, ZhouJ, LiHR, XiaX, LiuQ (2016). Eco-physiological responses of linseed ( Linum usitatissimum) to salt and alkali stresses. Chinese Journal of Plant Ecology, 40, 69-79. (in Chinese with English abstract)[郭瑞, 李峰, 周际, 李昊儒, 夏旭, 刘琪 (2016). 亚麻响应盐、碱胁迫的生理特征 . 植物生态学报, 40, 69-79.] [本文引用: 1]
[4]
GuoW, YuLH (2012). Effects of salinity-alkalinity stress on root activity and phenylalanine ammonia-lyase activity of wheat seedlings. Crops, (1), 31-34. (in Chinese with English abstract)[郭伟, 于立河 (2012). 盐碱胁迫对小麦幼苗根系活力和苯丙氨酸解氨酶活性的影响 . 作物杂志, (1), 31-34.] [本文引用: 1]
[5]
HuHY, HeJ, ZhaoJJ, RuZG (2013). Dynamic change of cell defense enzyme activity in wheat seedling under different alkaline pH conditions. Journal of Henan Institute of Science and Technology (Natural Science Edition), 41(5), 1-5. (in Chinese with English abstract)[胡海燕, 贺杰, 赵俊杰, 茹振刚 (2013). 碱性pH条件下小麦幼苗保护酶活性的变化动态 . 河南科技学院学报(自然科学版), 41(5), 1-5.]
[6]
JianaerAH, YangCW, ShiDC, WangDL (2007). Physiological response of an alkali resistant halophyte Kochia sieversiana to salt and alkali stresses. Acta Botanica Boreali-Occidentalia Sinica, 27, 79-84. (in Chinese with English abstract)[贾娜尔·阿汗, 杨春武, 石德成, 王德利 (2007). 盐生植物碱地肤对盐碱胁迫的生理响应特点 . 西北植物学报, 27, 79-84. ] [本文引用: 1]
[7]
JiangSX, LiuDX, PangHX, LüJY (2014). Effects of PEG stress and recovery on activities of key enzymes involved in proline metabolism in wheat cultivars with difference in drought tolerance. Acta Botanica Boreali-Occidentalia Sinica, 34, 1581-1587. (in Chinese with English abstract)[姜淑欣, 刘党校, 庞红喜, 吕金印 (2014). PEG胁迫及复水对不同抗旱性小麦幼苗脯氨酸代谢关键酶活性的影响 . 西北植物学报, 34, 1581-1587.] [本文引用: 1]
[8]
KerepesiI, GalibaG (2000). Osmotic and salt stress induced alteration in soluble carbohydrate content in wheat seedlings. Crop Science, 40, 482-487. [本文引用: 1]
[9]
KingsburyRW, EpsteinE, PearyRW (1984). Physiological responses to salinity in selected lines of wheat. Plant Physiology, 74, 417-423. [本文引用: 2]
[10]
LiB, WangZC, SunZG, ChenY, YangF (2005). Resources and sustainable resource exploitation of salinized land in China. Agricultural Research in the Arid Areas, 23(2), 152-158. (in Chinese with English abstract)[李彬, 王志春, 孙志高, 陈渊, 杨福 (2005). 中国盐碱地资源与可持续利用研究 . 干旱地区农业研究, 23(2), 152-158.] [本文引用: 1]
[11]
LisecJ, SchauerN, KopkaJ, WillmitzerL, FernieAR (2006). Gas chromatography mass spectrometry-based metabolite profiling in plants. Nature Protocol, 1, 387-396. [本文引用: 1]
[12]
LuoYY, LiuSK (2009). Research progress of ammonium transporter in plants. Genomics and Applied Biology, 28, 373-379. (in Chinese with English abstract)[骆媛嫒, 柳参奎 (2009). 植物中铵转运蛋白的研究进展 . 基因组学与应用生物学, 28, 373-379.] [本文引用: 2]
[13]
MunnsR, TesterM (2008). Mechanisms of salinity tolerance. Annual Review of Plant Biology, 59, 651-681. [本文引用: 2]
[14]
ParidaAK, DasAB (2005). Salt tolerance and salinity effects on plants: A review. Ecotoxicology and Environmental Safety, 60, 324-349. [本文引用: 1]
[15]
ParvanehR, ShahrokhT, HosseiniSM (2012). Studying of salinity stress effect on germination, proline, sugar, protein, lipid and chlorophyll content in purslane ( Portulace oloracea L.) leaves. Stress Physiology & Biochemistry, 8, 182-193. [本文引用: 1]
[16]
PengZH, PengKQ, HuJJ, XiaoLT (2002). Research progress on accumulation of proline under osmotic stress in plants. Chinese Agriculture Science Bulletin, 18(4), 80-83. (in Chinese with English abstract)[彭志红, 彭克勤, 胡家金, 萧浪涛 (2002). 渗透胁迫下植物脯氨酸积累的研究进展 . 中国农学通报, 18(4), 80-83.]
[17]
SaRL, LiuJH, LiuL, BaiJH, WangZH (2014). Cation- responsive mechanisms of oats to alkali stress. Acta Agronomica Sinica, 40, 362-368. (in Chinese with English abstract)[萨如拉, 刘景辉, 刘伟, 白健慧, 王占海 (2014). 燕麦对碱胁迫的阳离子响应机制 . 作物学报, 40, 362-368.] [本文引用: 2]
[18]
ShiDC, WangDL (2005). Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.) Kitag. Plant and Soil, 271, 15-26.
[19]
ShiLX, MaS, FangY, XuJ (2015). Crucial variations in growth and ion homeostasis of glycine gracilis seedlings under two types of salt stresses. Journal of Soil Science and Plant Nutrition, 15, 1-17.
[20]
WangH, WuZH, HanJY, ZhengW, YangCW (2012). Comparison of ion balance and nitrogen metabolism in old and young leaves of alkali-stressed rice plants. PLOS ONE, 7, e37817. doi: 10.1371/journal.pone.0037817.
WangXD, WangC, MaZH, HouRF, GaoQ, ChenQ (2011). Effect of short-term salt stress on the absorption of K+ and accumulation of Na+, K+ in seedlings of different wheat varieties. Acta Ecologica Sinica, 31, 2822-2830. (in Chinese with English abstract)[王晓冬, 王成, 马智宏, 侯瑞锋, 高权, 陈泉 (2011). 短期NaCl胁迫对不同小麦品种幼苗K+吸收和Na+、K+积累的影响 . 生态学报, 31, 2822-2830.]
[23]
WangXP, GengSJ, RiYJ, CaoDH, LiuJ, ShiDC, YangCW (2011). Physiological responses and adaptive strategies of tomato plants to salt and alkali stresses. Scientia Horticulturae, 130, 248-255. [本文引用: 2]
[24]
WuDZ, ShenQF, CaiSG, ChenZH, DaiF, ZhangGP (2013). Ionomic responses and correlations between elements and metabolites under salt stress in wild and cultivated barley. Plant & Cell Physiology, 54, 1976-1988. [本文引用: 5]
[25]
YangCW, LiCY, ZhangML, LiuJ, JuM, ShiDC (2008). pH and ion balance in wheat-wheatgrass under salt-or alkali stress. Chinese Journal of Applied Ecology, 19, 1000-1005. (in Chinese with English abstract)[杨春武, 李长有, 张美丽, 刘杰, 鞠淼, 石德成 (2008). 盐、碱胁迫下小冰麦体内的pH及离子平衡 . 应用生态学报, 19, 1000-1005.]
[26]
YangC, ShiD, WangD (2008). Comparative effects of salt stress and alkali stress on growth, osmotic adjustment and ionic balance of an alkali resistant halophyte Suaeda glauca(Bge.). Plant Growth Regulation, 56, 179-190. [本文引用: 1]
[27]
YangT, XieZX, YuQ, LiuXJ (2014). Effects of partial root salt stress on seedling growth and photosynthetic characteristics of winter wheat. Chinese Journal of Eco- Agrculture, 22, 1074-1078. (in Chinese with English abstract)[杨婷, 谢志霞, 喻琼, 刘小京 (2014). 局部根系盐胁迫对冬小麦生长和光合特征的影响 . 中国生态农业学报, 22, 1074-1078.] [本文引用: 1]
[28]
YuRP (1999). Saline soil resources in China and their exploitation. Chinese Journal of Soil Science, 30(4), l58-159. (in Chinese with English abstract)[俞仁培 (1999). 我国盐渍土资源及其开发利用 . 土壤通报, 30(4), l58-159.] [本文引用: 1]
[29]
ZhuJK (2003). Regulation of ion homeostasis under salt stress. Current Opinion in Plant Biology, 6, 441-445.
 , 周际
, 周际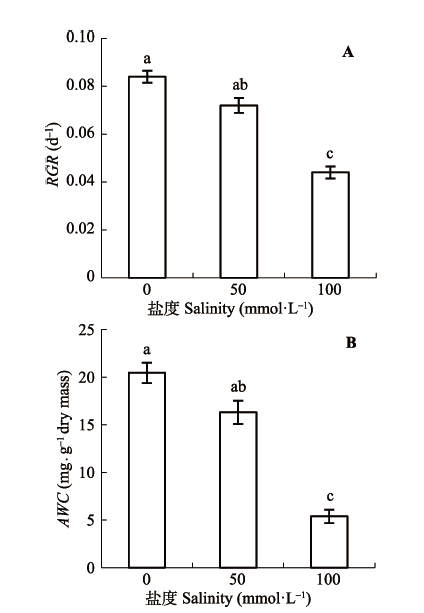 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT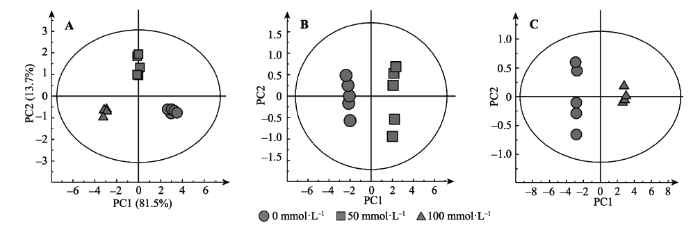 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT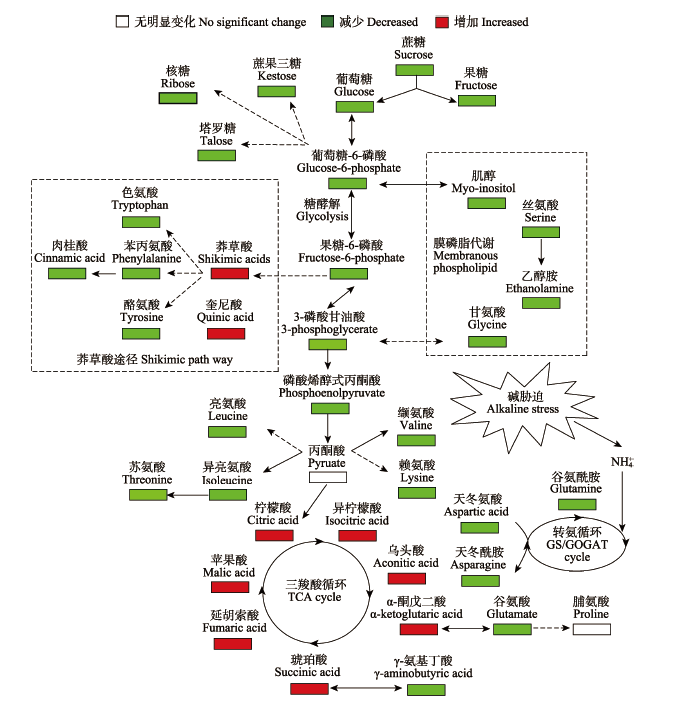 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}