Canopy structure and radiation interception of Salix matsudana: Stand density dependent relationships
CHENJing, ZHAOCheng-Zhang*,, WANGJi-Wei, ZHAOLian-Chun College of Geography and Environmental Science, Northwest Normal University, Research Center of Wetland Resources Protection and Industrial Development Engineering of Gansu Province, Lanzhou 730070, China 通讯作者:* 通信作者Author for correspondence (E-mail: zhaocz601@163.com) 收稿日期:2017-05-3 接受日期:2016-08-4 网络出版日期:2017-07-28 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:基金项目 国家自然科学基金(41461013和91125014)
关键词:光截获;冠层厚度;冠层面积;密度;旱柳 Abstract Aims Plants can enhance their photosynthetic efficiency and competitiveness by adjusting canopy structure and radiation interception. The objective of this paper was to quantify the relationship between canopy structure (crown depth and crown area) and light interception (LI) in a Salix matsudana stand under three different stand densities in a flood plain of Zhangye.Methods Our study site is located at the Heihe flood plain of Xichengyi in Ganzhou district, Zhangye City, Gansu Province in the middle Heihe River, where S. matsudana is the dominant species. Based on stand density (10 m × 10 m), the S. matsudana community is divided into three types: low density (I, 25-36 Ind.·plot-1), medium density (II, 37-48 Ind.·plot-1), and high density (III, 49-60 Ind.·plot-1). Community characteristics, soil physical and chemical properties of each type were measured. At each plot, we measured photosynthetically active radiation (PAR), LI, net photosynthetic rate (Pn), transpiration rate (Tr), crown depth, crown area, leaf area index (LAI), twig numbers, twig length, and bifurcation angle. The standardized major axis (SMA) estimation method was used to determine the relationships between LI and canopy structure.Important findings With increasing in stand density, we found that soil moisture increased, and soil electric conductivity decreased, while twig length and crown depth increased, and PAR, twig numbers, bifurcation angle and crown area decreased. LAI and LI, Pn and Tr reached their maximum at the stand of medium density. There was a significant, positive correlation and negative correlation (p < 0.01), respectively, between the LI, crown depth and crown area at low density (I), whereas low significant (p < 0.05) at high density (III), and high significantly positive correlation (p < 0.01) at the medium density (II). S. matsudana has more horizontal branches that reduce LI. Canopy thickness and increased crown area at low density. More vertical distribution of branches at high density, and a more balanced spacial distribution were found at medium density.
Keywords:light interception;crown depth;crown area;density;Salix matsudana -->0 PDF (1048KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 陈静, 赵成章, 王继伟, 赵连春. 不同密度旱柳的树冠构型与光截获. 植物生态学报, 2017, 41(6): 661-669 https://doi.org/10.17521/cjpe.2016.0257 CHENJing, ZHAOCheng-Zhang, WANGJi-Wei, ZHAOLian-Chun. Canopy structure and radiation interception of Salix matsudana: Stand density dependent relationships. Chinese Journal of Plant Ecology, 2017, 41(6): 661-669 https://doi.org/10.17521/cjpe.2016.0257 森林冠层结构决定了光能在冠层内的传输和分布方式(Goel, 1988), 对植物利用生态空间的能力和提高光合收益具有重要意义。光截获(LI)和冠层形态结构能够反映植物冠层适应特定生境的基本行为和光合生理功能, 体现了植物进行资源优化配置的生态适应与生存策略(Poorter et al., 2003)。LI作为冠层内部光分布特征的一个评价指标, 反映了树冠对太阳光的截取能力(Wunsche et al., 1996), 是度量群体内光照状况及光捕获能力的重要参数(李世莹等, 2013); 树冠构型可以描述为垂直和水平截面上的形状, 其中冠层厚度或冠长表示树冠垂直空间大小, 表征树冠的纵向生长与邻体间的竞争效应(Thorpe et al., 2010), 树冠面积是研究树冠水平伸展空间结构的指标, 反映植物在水平方向上的光资源竞争能力(Kind et al., 2006)。植物可以通过调整冠层厚度和面积来适应生境变化, LI与冠层构型相互影响、相互制约的生态学机制是理解冠层内光合作用过程和光合碳固定的重要依据。 密度是自然界植物种群普遍存在的重要选择压力(Japhet et al., 2009), 随着种群密度的增加, 林冠内光量减少, 邻株的物理或几何上的拥挤效应会加剧种内资源竞争, 影响到植物的光资源获取能力, 植物为获取最大光资源量, 在垂直生长与侧向扩展间物质营养投入的权衡发生偏移, 引起构型转变(Clark, 2010), 促使植物对叶面积指数(LAI)、分枝角度、分枝数和枝长度进行调整, 构建与光截获效率相适应的冠幅厚度与冠幅面积, 体现了最大化地吸取异质性分布资源的觅食行为和可塑性反应。因此, 研究不同密度下林冠LI和冠层构型的关系, 对于理解植物的光截获对策具有重要意义。 旱柳(Salix matsudana)是中国北方主要道路防护林及沙荒造林树种, 抗寒喜光、生长快、繁殖容易, 具有较好的防风固沙和保持水土功能, 在改善生态环境中发挥了重要作用。近年来, 国内外****关于冠层光照与冠层构型的关系(Poorter et al., 2008; 杨晓东等, 2013)、湿地环境因子对植物光合和形态的影响(Rodríguez-González et al., 2010)、不同密度冠层构型的生态生理响应机制(章建红等, 2014)以及旱柳的光合效率(夏江宝等, 2013)、生长潜力(Sixto et al., 2015)和生理特性(Song et al., 2016)等方面有较多研究, 解释了植物适应特定环境的光照、光合生理、形态变化与表型塑造, 但光分布和旱柳冠层构型的关联性研究不够深入, 特别是在湿地群落不同密度生境中, LI和冠层厚度、冠层面积关系对植物生理功能和异质性资源配置的响应机理尚不清晰。鉴于此, 本研究以黑河中游张掖段洪泛平原湿地的天然旱柳林为研究对象, 研究旱柳在密度梯度上LI与冠层厚度和面积的关系, 旨在揭示湿地植物密度制约下光分布规律与表型可塑性。
1 材料和方法
1.1 研究区概况
研究区位于甘肃省黑河流域中游张掖市甘州区西城驿的黑河滩地, 地理位置为38.98° N, 100.40° E, 海拔1 482.7 m, 属温带大陆性气候, 年平均气温 7.8 ℃, ≥0 ℃积温为2 734 ℃, 年降水量132.6 mm, 主要集中在6-9月, 年蒸发量1 986.5 mm, 年日照时间3 077 h (韩玲等, 2016)。土壤以砾石土、灰棕荒漠土和草甸土为主, 植被分布具有明显的水平分异特征, 以湿生植物、盐生植物与陆生乔灌林木为主。主要植物有: 旱柳、宽苞水柏枝(Myricaria bracteata)、多枝柽柳(Tamarix ramosissima)、沙枣(Elaeagnus angustifolia)、苦豆子(Sophora alopecuroides)、芨芨草(Achnatherum splendens)、冰草(Agropyron cristatum)、芦苇(Phragmites australis)、节节草(Equisetum ramosissimum)等(韩玲等, 2016)。该区属于张掖黑河国家湿地核心区, 没有人类活动的影响, 较为真实地反映了湿地的自然属性与植物生物学特性的关系。
依据密度将100个样方划分为3个密度组(分别为I、II、III), 各组的样方数和旱柳种群特征见表1。对旱柳种群样地划分出的3个密度梯度各平均性状进行统计分析。旱柳光截获、冠层厚度和冠层面积的实验数据先进行以10为底的对数转换, 使之符合正态分布后再进行分析。数据分析主要运用标准化 主轴估计(SMA)的方法(Warton et al., 2006)对旱柳光截获和冠层构型间的关系进行分析, 采用Excel软件和SPSS 16.0统计分析软件对数据进行相关分析和方差分析, 显著性水平设为0.05, 方差分析前对所有数据进行方差齐性检验和正态分布检验。 Table 1 表1 表1样地分组情况和旱柳种群的生物学特征(平均值±标准误差) Table 1Biological characteristics of Salix matsudana population (mean ± SE) at three stands of variable density
密度分组 Density fractions
密度区间 Density interval (Ind.·plot-1)
样方数 Number of plots
高度 Height (cm)
胸径 Diameter at breast height (cm)
郁闭度 Crown density (%)
低密度 Low density (I)
25-36
26
398.57 ± 20.63c
13.06 ± 0.61a
42.84 ± 2.72c
中密度 Medium density (II)
37-48
38
454.69 ± 24.05b
10.72 ± 0.58b
69.56 ± 3.93b
高密度 High density (III)
49-60
36
521.23 ± 27.48a
8.64 ± 0.39c
87.09 ± 4.37a
Different lowercase letters in the same column indicate significant difference among density at p < 0.01 level.同列不同小写字母表示密度间差异显著(p < 0.01)。 新窗口打开
2 结果和分析
2.1 不同密度旱柳群落土壤性质与光合性状
不同密度旱柳群落土壤理化性质与光合特性的变化如表2所示。旱柳土壤电导率、PAR、LI、Pn、Tr和WUE在不同密度存在显著差异(p < 0.05), 随种群密度的增加, 土壤电导率、PAR和水分利用率呈逐渐减小趋势, 分别降低了69.55%、55.70%和24.36%; LI、Pn和Tr呈倒U形变化趋势: LI和Tr表现为II > III > I, 增幅分别达到26.56%和42.05%, 而Pn为II > I > III, 增幅达到47.84%; 土壤质量含水量在不同密度均出现较大值, 无显著差异(p > 0.05)。
2.2 不同密度旱柳冠层构型的变化
如图1所示, 旱柳冠层构型在不同密度的变化存在显著差异(p < 0.05)。随着密度的逐渐增加, 旱柳的冠层厚度与枝长度呈逐渐增加趋势, 从密度I到密度III分别增加了66.67%和44.80%; 冠层面积、分枝数和分枝角度呈逐渐减小趋势, 密度I到密度III分别减小24.69%、60.89%和28.33%; LAI呈先增大后减小的趋势, 变化趋势为II > III > I, 即在密度II出现最大值, 密度I出现最小值, 密度II的LAI较密度I与密度III分别增大了132.39%和23.13%。 显示原图|下载原图ZIP|生成PPT 图1不同密度条件下旱柳种群冠层构型的变化(平均值±标准误差)。不同小写字母表示在密度间差异显著(p < 0.05)。I、II、III, 同表1。 -->Fig. 1Canopy structure of Salix matsudana by density (mean ± SE). The lowercase letters indicate significant differences among density (p < 0.05). I, II, III, see Table 1. -->
2.3 不同密度旱柳冠层光截获与构型的关系
2.3.1 光截获与冠层厚度的关系 如图2所示, 旱柳冠层LI与冠层厚度在低密度I和中密度II呈极显著正相关关系(p < 0.01, 图2I、图2II), 高密度III呈显著正相关关系(p < 0.05, 图2III)。通过SMA分析得出3个不同密度梯度间的斜率无显著差异(p > 0.05), LI与冠层厚度回归关系具有共同斜率0.74 (95%的置信区间CI = (0.67, 0.83)), 显著小于1 (p < 0.05), 表明LI增加比率大于冠层厚度的增加比率。随着种群密度逐渐增加, LI与冠层厚度的y轴截距显著下降(p < 0.05), 表明相同的LI条件下, 种群密度越低, 冠层厚度越小。 显示原图|下载原图ZIP|生成PPT 图2不同密度旱柳光截获与冠层厚度的关系。I、II、III, 同表1。 -->Fig. 2Relationship between light interception and crown depth of Salix matsudana by density. I, II, III, see Table 1. -->
2.3.2 光截获与冠层面积的关系 如图3所示, 旱柳冠层LI与冠层面积在低密度I和高密度III分别呈极显著负相关(p < 0.01, 图3I)与显著负相关(p < 0.05, 图3III), 中密度II则呈极显著正相关关系(p < 0.01, 图3II); 随着种群密度逐渐增加, 旱柳LI和冠层面积回归方程的SMA斜率分别为-0.55 (95%的置信区间CI = (-0.72, -0.39))、0.52 (95%的置信区间CI = (0.43, 0.62))、-0.63 (95%的置信区间CI = (-0.78, -0.45)), 低密度和高密度旱柳的SMA斜率均与-1存在极显著差异(p < 0.01), 中密度与1存在极显著差异(p < 0.01), 表明旱柳在低密度和高密度时, LI越大、冠层面积越小, 不同密度生境中呈不同的权衡关系; 在中密度时, 旱柳LI越大、冠层面积越大, LI增加的速度大于冠层面积增加的速度。 显示原图|下载原图ZIP|生成PPT 图3不同密度旱柳冠层光截获与冠层面积的关系。I、II、III, 同表1。 -->Fig. 3Relationship between light interception and crown area of Salix matsudana by density. I, II, III, see Table 1. -->
密度差异形成各种不同的光环境, 引起植株冠层结构的动态变化, 使得植株叶片和枝条的形态(Stenberg et al., 1998)和树冠形态(King, 1997)等都表现出高度的可塑性。本研究中, 低密度样地旱柳光截获与冠层厚度和面积分别呈极显著正相关和负相关, 形成小冠层厚度、大冠层面积的冠层构型(图1), 这符合段劼等(2010)提出的“林分密度较低时树冠水平表面积大、生长饱满”的观点。主要原因可能在于: (1)低密度种群的光、热和空间资源充足, 种群竞争较弱, 旱柳为了拓展生存生态位, 优先倾向于增加枝条数和分枝角度, 从而选择了减少枝长度和单位分枝上叶面积的资源投资, 促使枝条水平伸展、叶片松散排列在不同的分枝上表现为LAI很低(王俊峰和冯玉龙, 2004), 较小的LAI决定了植物冠层较少的光截获量(图1)(Depury & Farquhar, 1997); (2)邻体遮挡较少且PAR最大(表2), 常常遭受强光胁迫, 冠层内部枝条之间的自我荫蔽程度主要与植株冠层形态特征有关, 冠层越深、幅度越小的植株个体, 其自我荫蔽程度则越弱, 反之亦然(Osunkoya et al., 2007), 为了躲避强光胁迫, 植物选择增大胸径和冠幅而降低株高与冠幅深度, 这是旱柳在强光下的自我保护构造和安全性经济权衡策略; (3)结合表1、表2和图1资料认为, 低密度生境地表盐分积累尤为明显, 电导率最大, 盐分胁迫造成的“生理干旱”现象阻碍了植物的吸水作用(周洪华和李卫红, 2015), 使植物的生长高度和冠幅厚度都受到限制。随密度改变, 旱柳净光合速率的下降幅度小于蒸腾速率, 较高的水分利用率有利于旱柳将更多的光合产物投入到与木质部水分、养分输导有关的系统构建方面(史元春等, 2015), 横向扩展冠幅、加粗树干以维持机械支持与导管畅通, 为水分转移至树顶提供更大的接触面积和便捷的路径。因此, 旱柳在低密度时, 光和盐胁迫加剧, 促使植物趋向于矮状化, 采取减少光截获的水平空间拓展模式来应对环境胁迫。 Table 2 表2 表2不同密度旱柳群落土壤性质与光合性状(平均值±标准误差) Table 2Soil characteristics and photosynthetic characteristics of Salix matsudana stand by density (mean ± SE)
密度 Density
I
II
III
土壤质量含水量 Mass moisture of soil (%)
31.56 ± 1.52a
34.83 ± 1.87a
38.47 ± 2.01a
土壤电导率 Soil electric conductivity (ms·cm-1)
7.75 ± 0.41a
4.09 ± 0.21b
2.36 ± 0.13c
光合有效辐射 PAR (μmol·m-2·s-1)
981.26 ± 50.14a
740.51 ± 39.08b
434.67 ± 24.34c
光截获 LI
0.64 ± 0.03c
0.81 ± 0.05a
0.73 ± 0.04b
净光合速率 Pn (μmol·m-2·s-1)
10.27 ± 0.53b
13.69 ± 0.61a
9.26 ± 0.47c
蒸腾速率 Tr (mmol·m-2·s-1)
4.59 ± 0.23c
6.52 ± 0.34a
5.24 ± 0.27b
水分利用率 WUE (μmol·mmol-1)
2.34 ± 0.11a
2.10 ± 0.09b
1.77 ± 0.08c
PAR, photosynthetically active radiation; LI, light interception; Pn, net photosynthetic rate; Tr, transpiration rate; WUE, water use efficiency. Different lowercase letters in the same line indicate significant differences of different density for identical module (p < 0.05). I, II, III, see Table 1.同行不同小写字母表示密度间差异显著(p < 0.05)。I、II、III, 同表1。 新窗口打开
DepuryDGG, FarquharGD (1997). Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models. Plant, Cell & Environment, 20, 537-557. [本文引用: 1]
[4]
DuJ, ZhaoCZ, SongQH, ShiYC, WangJW, ChenJ (2016). Plant size differences with twig and leaf traits of Zygophyllum xanthoxylum in the northern slope of Qilian Mountains, China . Chinese Journal of Plant Ecology, 40, 212-220. (in Chinese with English abstract)[杜晶, 赵成章, 宋清华, 史元春, 王继伟, 陈静 (2016). 祁连山北坡霸王枝-叶性状关系的个体大小差异 . 植物生态学报, 40, 212-220.] [本文引用: 1]
[5]
DuanJ, MaLY, JiaLM, XuCY, JiaZK, CheWR (2010). The density effect of Platycladus orientalis plantation in Beijing area. Acta Ecologica Sinica, 30, 3206-3214. (in Chinese with English abstract)[段劼, 马履一, 贾黎明, 徐程扬, 贾忠奎, 车文瑞 (2010). 北京地区侧柏人工林密度效应 . 生态学报, 30, 3206-3214.]
[6]
GoelNS (1988). Models of vegetation canopy rellectance and their use in estimation of biophysical parameters from rellectance data. Remote Sening Reviews, 4, 1-221. [本文引用: 1]
[7]
GongR, GaoQ (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions. Chinese Journal of Plant Ecology, 39, 300-308. (in Chinese with English abstract)[龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展 . 植物生态学报, 39, 300-308.] [本文引用: 1]
[8]
GuoJ, XiaoK, GuoXY, ZhangFL, ZhaoCZ (2005). Review on maize canopy structure, light distributing and canopy photosynthesis. Journal of Maize Science, 13(2), 55-59. (in Chinese with English abstract)[郭江, 肖凯, 郭新宇, 张凤路, 赵春江 (2005). 玉米冠层结构、光分布和光合作用研究综述 . 玉米科学, 13(2), 55-59.] [本文引用: 1]
[9]
HanL, ZhaoCZ, XuT, FengW, DuanBB, ZhengHL.Trade-off between leaf size and vein density ofAchnatherum splendens in Zhangye wetland .Chinese Journal of Plant Ecology, 40, 788-797. (in Chinese with English abstract)[韩玲, 赵成章, 徐婷, 冯威, 段贝贝, 郑慧玲 (2016). 张掖湿地芨芨草叶大小和叶脉密度的权衡关系 . 植物生态学报, 40, 788-797.] [本文引用: 3]
[10]
JaphetW, ZhouDW, ZhangHX, ZhangHX, YuT (2009). Evidence of phenotypic plasticity in the response of Fagopyrum esculentum to population density and sowing date. Journal of Plant Biology, 52, 303-311. [本文引用: 1]
[11]
KanniahKD, BeringerJ, NorthP, HutleyL (2012). Control of atmospheric particles on diffuse radiation and terrestrial plant productivity: A review. Progress in Physical Geography, 36, 209-237. [本文引用: 1]
[12]
KindDA, DaviesSJ, NoorNSM (2006). Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest. Forest Ecology and Management, 223, 152-158. [本文引用: 1]
[13]
KingDA (1997). Branch growth and biomass allocation in Abies amabilis saplings in contrasting light environments. Tree Physiology, 17, 251-258. [本文引用: 1]
[14]
LiL, ZhouDW, ShengLX (2011). Density dependence- determined plant biomass allocation pattern. Chinese Journal of Ecology, 30, 1579-1589. (in Chinese with English abstract)[黎磊, 周道玮, 盛连喜 (2011). 密度制约决定的植物生物量分配格局 . 生态学杂志, 30, 1579-1589.] [本文引用: 1]
[15]
LiSY, FengW, WangYH, WangCY, GuoTC (2013). Effects of spacing interval of wide bed planting on canopy characteristics and yield in winter wheat. Chinese Journal of Plant Ecology, 37, 758-767. (in Chinese with English abstract)[李世莹, 冯伟, 王永华, 王晨阳, 郭天财 (2013). 宽幅播种带间距对冬小麦冠层特征及产量的影响 . 植物生态学报, 37, 758-767.] [本文引用: 1]
[16]
MaZQ, LiuQJ, ZengHQ, LiXR, ChenYR, ZhangSH, YangFD, WangHQ (2008). Estimation of leaf area index of planted forests in subtropical China by photogrammetry. Acta Ecologica Sinica, 28, 1971-1980. (in Chinese with English abstract)[马泽清, 刘琪憬, 曾慧卿, 李轩然, 陈永瑞, 林耀明, 张时煌, 杨风亭, 汪宏清 (2008). 南方人工林叶面积指数的摄影测量 . 生态学报, 28, 1971-1980.] [本文引用: 1]
[17]
OsunkoyaOO, Omar-AliK, AmitN, DayanJ, DaudDS, ShengTK (2007). Comparative height crown allometry and mechanical design in 22 tree species of Kuala Belalong rainforest, Brunei, Borneo. American Journal of Botany, 94, 1951-1962. [本文引用: 2]
[18]
PoorterL, BongersF, SterckFJ, WollH (2003). Architecture of 53 rain forest tree species differing in adult stature and shade tolerance. Ecology, 84, 602-608. [本文引用: 1]
[19]
PoorterL, WrightSJ, PazH, AckerlyDD, ConditR, IbarraManriquezG, HarmsKE, LiconaJC, Martínez-RamosM, MazerSJ, Muller-LandauHC, Peña-ClarosM, WebbCO, WrightIJ (2008). Are functional traits good predictors of demographic rates? Evidence from five neotropical forests. Ecology, 89, 1908-1920. [本文引用: 1]
[20]
Rodríguez-GonzálezPM, StellaJC, CampeloF, Ferreira MT AlbuquerqueA (2010). Subsidy or stress? Tree structure and growth in wetland forests along a hydrological gradient in Southern Europe. Forest Ecology & Management, 259, 2015-2025. [本文引用: 1]
[21]
ShiYC, ZhaoCZ, SongQH, DuJ, WangJW (2015). Allometric relationship between height and crown width or diameter of Platycladus orienalis on different slope aspects of Lanzhou northern mountains. Chinese Journal of Ecology, 34, 1879-1885. (in Chinese with English abstract)[史元春, 赵成章, 宋清华, 杜晶, 王继伟 (2015). 兰州北山侧柏株高与冠幅、胸径异速生长关系的坡向差异性 . 生态学杂志, 34, 1879-1885.] [本文引用: 1]
[22]
SixtoH, CañellasI, ArendonkaJV, CiriaP, CampsF, SánchezM, Sánchez-GonzálezM (2015). Growth potential of different species and genotypes for biomass production in short rotation in Mediterranean environments. Forest Ecology and Management, 354, 291-299. [本文引用: 1]
[23]
SongX, FangJ, HanX, HeX, LiuM, HuJ, ZhuoR (2016). Overexpression of quinone reductase from Salix matsudana Koidz enhances salt tolerance in transgenic Arabidopsis thaliana. Gene, 576, 520-527. [本文引用: 1]
[24]
StenbergP, SmolanderH, SprugelD, SmolanderS (1998). Shoot structure, light interception, and distribution of nitrogen in an Abies amabilis canopy. Tree Physiology, 18, 759-767. [本文引用: 1]
[25]
SunHG, ChenYT (2010). Root growth patterns of four coastal shelter forest tree species in response to salt stress. Chinese Journal of Ecology, 29, 2365-2372. (in Chinese with English abstract)[孙洪刚, 陈益泰 (2010). 沿海防护林四个树种根系分布对盐胁迫的响应 . 生态学杂志, 29, 2365-2372.] [本文引用: 1]
[26]
ThorpeHC, AstrupR, TrowbridgeA, CoatesKD (2010). Competition and tree crowns: A neighborhood analysis of three boreal tree species. Forest Ecology and Management, 259, 1586-1596. [本文引用: 1]
[27]
WangJF, FengYL (2004). The effect of light intensity on blomass allocation, leaf morphology and relative growth rate of two invasive plants. Acta Phytoecologica Sinica, 28, 781-786. (in Chinese with English abstract)[王俊峰, 冯玉龙 (2004). 光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响 . 植物生态学报, 28, 781-786.] [本文引用: 1]
WunscheJN, LaksoAN, RobinsonTL, LenzF, DenningSS (1996). The bases of productivity in apple production systems: The role of light interception by different shoot types. Journal of the American Society for Horticultural Science, 121, 886-893. [本文引用: 1]
[30]
XiaJB, ZhangSY, ZhaoZG, ZhaoYY, GaoY, GuGY, SunJK (2013). Critical effect of photosynthetic efficiency in Salix matsudana to soil moisture and its threshold grade in shell ridgei sland. Chinese Journal of Plant Ecology, 37, 851-860. (in Chinese with English abstract)[夏江宝, 张淑勇, 赵自国, 赵艳云, 高源, 谷广义, 孙景宽 (2013). 贝壳堤岛旱柳光合效率的土壤水分临界效应及其阈值分级 . 植物生态学报, 37, 851-860.] [本文引用: 1]
[31]
YangXD, YanER, ZhangZH, SunBW, HuangHX, ARSHADA, MaWJ, ShiQR (2013). Tree architecture of overlapping species among successional stages in evergreen broad-leaved forests in Tiantong region, Zhejiang Province, China. Chinese Journal of Plant Ecology, 37, 611-619. (in Chinese with English abstract)[杨晓东, 阎恩荣, 张志浩, 孙宝伟, 黄海侠, ARSHADA, 马文济, 史青茹 (2013). 浙江天童常绿阔叶林演替阶段共有种的树木构型 . 植物生态学报, 37, 611-619.] [本文引用: 2]
[32]
ZhangH, LiJN, WangGX, DaiXF, QiuMQ, ZhengKF. (2008). Plants interactions between Suaeda salsa individuals are mediated by salinity stress. Acta Physiologiae Plantarum, 30, 99-104. [本文引用: 1]
[33]
ZhangJH, ShiQR, XuMS, ZhaoYT, ZhongQ, ZhangFJ, YanER (2014). Testing of Corner’s rules across woody plants in Tiantong region, Zhejiang Province: Effects of individual density. Chinese Journal of Plant Ecology, 38, 655-664. (in Chinese with English abstract)[章建红, 史青茹, 许洺山, 赵延涛, 仲强, 张富杰, 阎恩荣 (2014). 浙江天童木本植物Corner法则的检验: 个体密度的影响 . 植物生态学报, 38, 655-664.] [本文引用: 3]
[34]
ZhangWF, WangZL, YuSL, LiSK, FangJ, TongWS (2004). Effects of planting density on canopy photosynthesis, canopy structures and yield formation of high-yield cotton in Xinjiang, China. Acta Phytoecologica Sinica, 28, 164-171. (in Chinese with English abstract)[张旺锋, 王振林, 余松烈, 李少昆, 房建, 童文崧 (2004). 种植密度对新疆高产棉花群体光合作用、冠层结构及产量形成的影响 . 植物生态学报, 28, 164-171.] [本文引用: 1]
[35]
ZhangYQ, LiangCZ, WangW, WangLX, PengJT, YanJC, JiaJC (2010). Soil salinity and Achnatherum splendens distribution. Chinese Journal of Ecology, 29, 2438-2443. (in English with Chinese abstract)[张雅琼, 梁存柱, 王炜, 王立新, 彭江涛, 闫建成, 贾成朕 (2010). 芨芨草群落土壤盐分特征 . 生态学杂志, 29, 2438-2443.] [本文引用: 1]
[36]
ZhouHH, LiWH (2015). Responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica. Chinese Journal of Plant Ecology, 39, 81-91 (in English with Chinese abstract)[周洪华, 李卫红 (2015). 胡杨木质部水分传导对盐胁迫的响应与适应 . 植物生态学报, 39, 81-91.] [本文引用: 1]
Individuals and the variation needed for high species diversity in forest trees. 1 2010
 , 王继伟, 赵连春
, 王继伟, 赵连春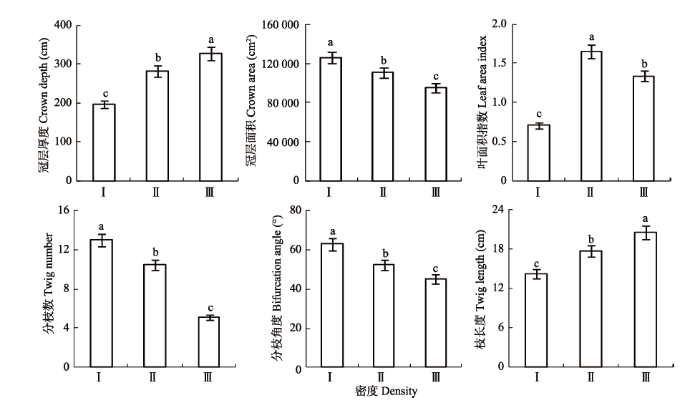 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT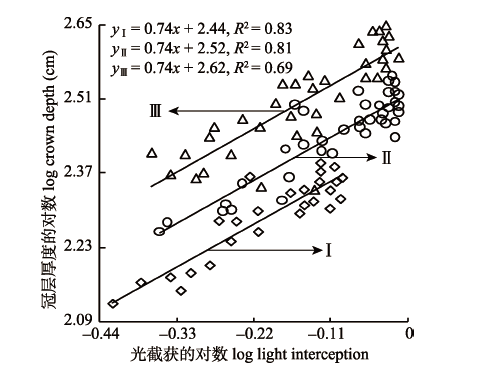 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT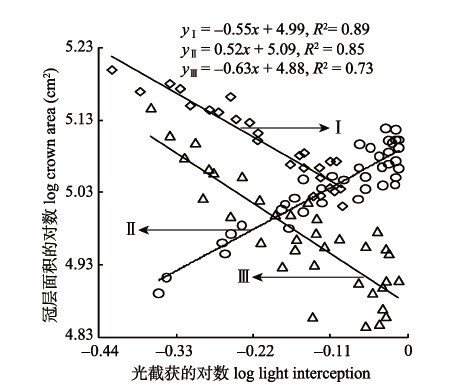 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}