Effects of tree mortality on the spatial patterns and interspecific associations of plant species in a Quercus aliena var. acuteserrata forest in Baotianman, Henan, China
WEIBo-Liang1,*, YUANZhi-Liang1,*, NIUShuai1, LIUXia2, JIAHong-Ru3, YEYong-Zhong1,**, 1College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China2Huaxian Environmental Protection Agency, Huaxian, Henan 456400, Chinaand 3Henan College of Finance and Taxation, Zhengzhou 451464, China 通讯作者:** 通信作者Author for correspondence (E-mail: yeyzh@163.com) 责任编辑:WEIBo-LiangYUANZhi-LiangNIUShuaiLIUXiaJIAHong-RuYEYong-Zhong 收稿日期:2016-09-26 接受日期:2017-01-3 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家科技支撑计划(2008BAC39802)
关键词:死亡;随机死亡假说;空间格局;种间相关性;锐齿槲栎 Abstract Aims Tree mortality is an important ecological process in forest ecosystems. The aims of this study were to determine how tree mortality influences the spatial patterns and interspecific associations of plant species, and what are the causes of tree mortality in a 1 hm2 permanent plot in Baotianman National Nature Reserve, Nanyang City, Henan Province. Methods We conducted field investigations in the plot and used spatial point pattern analysis to examine the spatial patterns and interspecific associations of 17 species prior to and following mortality. Important findings (1) Most of the species in the study plot showed an aggregated distribution both pre- and post-mortality. However, the number of species showing aggregated distribution decreased and the number of species showing random distribution increased following the mortality event. (2) Most species were positively associated with Quercus aliena var. acuteserrata both pre- and post-mortality, while some had no apparent association. Following tree mortality, on fine scales, the number of species with positive associations increased, and the number of species with negative associations decreased. (3) Tree mortality was in consistency with the random death hypothesis. The interspecific associations of four species with Q. aliena var. acuteserrata completely changed following death. For most species, the spatial patterns and the interspecific association with Q. aliena var. acuteserrata either changed at minor scales or did not change. The variations in spatial patterns or interspecific associations were inconsistent among species. (4) The dead trees of Q. aliena var. acuteserrata were significantly associated with the living trees in 13 species, but the associations between dead and living trees were not in agreement with the changes in interspecific association following mortality. Only five living tree species competed with the dead trees of Q. aliena var. acuteserrata, and the competition between each of these species and Q. aliena var. acuteserrata intensified after individual death. Tree mortality is the result of a variety of factors. Although the tree mortality in the study plot was in accordance with the random death hypothesis, there were also a few individuals which were dead from competition.
Keywords:mortality;random death hypothesis;spatial patterns;interspecific associations;Quercus aliena var. acuteserrata -->0 PDF (3176KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 韦博良, 袁志良, 牛帅, 刘霞, 贾宏汝, 叶永忠. 河南省宝天曼锐齿槲栎林树木死亡对空间格局及种间相关性的影响. 植物生态学报, 2017, 41(4): 430-438 https://doi.org/10.17521/cjpe.2016.0297 WEIBo-Liang, YUANZhi-Liang, NIUShuai, LIUXia, JIAHong-Ru, YEYong-Zhong. Effects of tree mortality on the spatial patterns and interspecific associations of plant species in a Quercus aliena var. acuteserrata forest in Baotianman, Henan, China. Chinese Journal of Plant Ecology, 2017, 41(4): 430-438 https://doi.org/10.17521/cjpe.2016.0297 种群空间格局是种群自身特性的体现, 在一定程度上反映了物种的资源利用方式、种内种间关系以及物种与环境的相互作用(刘妍妍和金光泽, 2010; 缪宁等, 2011)。物种空间格局的形成是多种生态过程相互作用的结果(范娟等, 2012)。随着种群内个体的不断生长、繁殖和死亡, 种群的空间格局以及种间关系会发生一定变化(李林等, 2014)。 树木死亡是森林群落动态的重要环节, 它对群落内物种组成、群落环境、森林生态系统物质循环以及物种的空间分布有较大影响(Domke et al., 2011; 王利伟等, 2011; 郭屹立等, 2016), 并在群落更新及演替过程中发挥着重要作用(Rouvinen et al., 2002)。树木死亡的原因很多, 例如树木衰老、自然灾害、病害虫害、动物取食以及个体之间的竞争等(Metcalf et al., 2009; 卢志军等, 2015)。 树木的死亡直接影响了种群的数量特征, 并改变了物种的空间点位(王斌等, 2014)。不同的死亡原因使物种的空间格局和种间相关性发生不同的变化。多数研究认为: 若树木是随机死亡的, 则物种的空间格局不会发生明显改变(缪宁等, 2011); 若树木的死亡是由密度制约所致, 则死亡后物种的空间格局更加随机(He & Duncan, 2000; Zhang et al., 2009); 而竞争导致的死亡会使物种种内种间相关性发生显著改变(徐嘉等, 2007)。 对于死亡树木的空间格局以及树木的死亡原因已有一些研究(郭屹立等, 2016)。然而这些研究通常以死亡树木为研究对象, 少有研究检验树木死亡前后种群空间格局和种间相关性的变化, 而对死树和活树的关系以及个体死亡后对物种种间关系影响的研究更加少见。为此, 本研究以宝天曼国家级自然保护区内一个1 hm2动态监测样地作为研究对象, 运用点格局分析的相关方法对数据进行分析, 试图回答以下几个问题: (1)在树木死亡前后, 不同物种的空间格局以及种间相关性有何变化?(2)死树的种间相关性能否反映树木死亡前后种群的种间相关性的变化?(3)样地内树木死亡的原因是什么?本研究希望通过对以上几个问题的研究深入探讨死亡对群落动态变化的影响, 并希望为进一步揭示树木的死亡机制提供更多理论依据。
1 研究地区和研究方法
1.1 研究地区概况
宝天曼国家级自然保护区位于河南省西南部内乡县境内, 地理位置为111.78°-112.06° E, 33.34°- 33.60° N, 总面积23 198 hm2, 最高峰海拔1 830 m (陈云等, 2014)。宝天曼自然保护区地处暖温带向北亚热带过渡地带, 植被属暖温带落叶林向北亚热带常绿阔叶林过渡的典型代表, 保护区内森林覆盖率高达97.8%, 是我国同纬度天然阔叶林保存最为完好的地区。宝天曼年平均气温15.2 ℃, 年降水量885.6 mm, 年蒸发量991.6 mm, 相对湿度68% (袁志良等, 2011)。
1.2 样地调查及样地概况
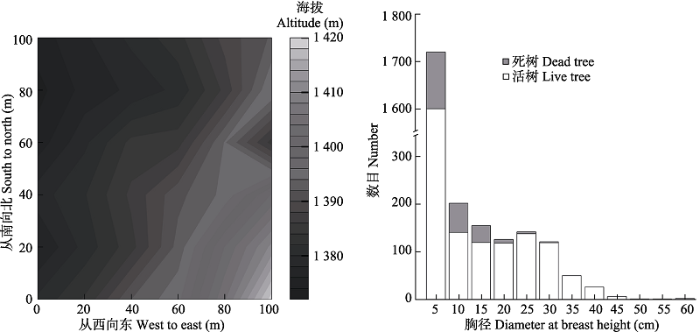
本研究于宝天曼内一个1 hm2长期监测样地中进行(图1)。参照巴拿马巴洛科罗拉多岛50 hm2热带雨林样地建设的技术规范(袁志良等, 2011), 用全站仪将样地分成25个20 m × 20 m的样方, 每个20 m × 20 m的样方又进一步分为16个5 m × 5 m的小样方。逐一测量记录样方内所有胸径(DBH)大于等于1 cm的木本植物(含死亡树木)的编号、名称、胸径、坐标和生长状况, 获得植物的调查数据。 显示原图|下载原图ZIP|生成PPT 图1样地地形图及17个物种整体的径级结构。 -->Fig. 1Topographic map in the plot and the frequency distribution of diameter at breast height of 17 species. -->
研究样地为锐齿槲栎(Quercus aliena var. acuteserrata)林, 该林型是河南落叶阔叶林区的主要森林植被类型(刘玉萃等, 2001)。样地内乔木层主要由锐齿槲栎、华东椴(Tilia japonica)、灯台树(Bothrocaryum controversum)等组成。灌木层主要由连翘(Forsythia suspensa)、白檀(Symplocos paniculata)、海州常山(Clerodendrum trichotomum)等组成。样地内个体数大于30的物种共有20种, 其中有17个物种有死亡个体。而死亡个体数大于10的物种仅有锐齿槲栎和连翘, 其余物种的死亡数均小于等于10 (表1)。 Table 1 表1 表1样地内物种活树和死树的基本特征 Table 1Basic characteristics of living and dead trees of species in the study plot
在个体死亡前后, 样地内17个物种的空间格局均为聚集分布。虽然死亡前后这些物种空间格局的函数值g(r)值发生一定变化, 但物种的格局仅在个别尺度上发生显著改变(附录I)。在格局出现显著变化的11个物种中, 大椴(Tilia nobilis)、海州常山、华榛(Corylus chinensis)、金银忍冬(Lonicera maackii)、秦岭木姜子(Litsea tsinlingensis)、锐齿槲栎、水榆花楸(Sorbus alnifolia)、四照花(Dendrobenthamia japonica var. chinensis)和卫矛(Euonymus alatus)的格局在少数尺度上由聚集分布变为随机分布; 桦叶荚蒾(Viburnum betulifolium)和三桠乌药(Lindera obtusiloba)在少数尺度上由随机分布变为聚集分布。此外, 白檀、鹅耳枥(Carpinus turczaninowii)、华东椴、连翘、山梅花(Philadelphus incanus)和崖樱桃(Cerasus scopulorum)这6个物种的空间格局无明显变化。 从图2可知, 树木死亡前后, 物种在小于10 m的尺度上以聚集分布为主, 在大于10 m的尺度上以随机分布为主。死亡前后物种的格局在小于5 m的尺度上没有明显变化; 在7-12 m的尺度上聚集分布的物种数有所减少; 在18-25 m的尺度上聚集分布的物种数明显减少, 而随机分布的物种数增多。但在随机死亡模型下, 物种的空间格局多数表现为随机分布, 仅个别物种在个别尺度上表现出聚集分布或规则分布。 显示原图|下载原图ZIP|生成PPT 图2物种个体死亡前(A)、死亡后(B)以及随机死亡(C)的空间分布格局百分比。 -->Fig. 2The percentage of spatial distribution patterns of species before individual death (A), after individual death (B), and random individual death (C). -->
2.2 死亡前后锐齿槲栎与其他物种的种间相关性
树木死亡前后锐齿槲栎与其他物种的相关性变化较小: 锐齿槲栎与白檀、桦叶荚蒾、华东椴和华榛这4个物种的种间相关性未发生改变, 与其他12个物种的种间相关性仅在个别尺度上发生改变(表2)。但由于死亡前锐齿槲栎与海州常山、金银忍冬、山梅花、卫矛的种间相关性多为不相关或仅在个别尺度上显著相关, 虽然死亡后锐齿槲栎与这4个物种的种间相关性仅在个别尺度上发生改变, 但死亡后的种间相关性与死亡前完全不同。此外锐齿槲栎与不同物种的相关性存在较大差异, 而死亡前后种间相关性的变化没有明显规律。 Table 2 表2 表2树木死亡前后锐齿槲栎和其他物种种间相关性变化及锐齿槲栎死树和其他物种活树的关系 Table 2Variations in interspecific associations between Quercus aliena var. acuteserrata and other species both pre- and post-mortality, and associations between the dead trees of Q. aliena var. acuteserrata and the living trees of other species
死树和活树的关系 Associations between dead trees and living trees
海州常山 Clerodendrum trichotomum
/
-
↓
-
连翘 Forsythia suspensa
-
-
↓
-
三桠乌药 Lindera obtusiloba
-
-
↓
-
四照花Dendrobenthamia japonica var. chinensis
-
-
↓
-
卫矛 Euonymus alatus
-+
-
↓
-
金银忍冬 Lonicera maackii
/
-
↓
/
白檀 Symplocos paniculata
-
-
/
+
桦叶荚蒾 Viburnum betulifolium
-
-
/
+
华东椴 Tilia japonica
+
+
/
-
华榛 Corylus chinensis
/
/
/
/
大椴 Tilia nobilis
-
-
↑
-+
山梅花 Philadelphus incanus
/
+
↑
-+
秦岭木姜子 Litsea tsinlingensis
+
+
↑
-
水榆花楸 Sorbus alnifolia
+
+
↑
-
崖樱桃 Cerasus scopulorum
+
+
↑
/
鹅耳枥 Carpinus turczaninowii
-+
-+
↓↑
+
“-” indicates negative association between species. “+” indicates positive association between species. “-+” indicates association between species is negative on some scales, and positive on some other scales."/" indicates no variation in the interspecific associations between species pre and postmortality, or there was no significant association between species. "↓" indicates interspecific competition becomes stronger or facilitation becomes weaker post-mortality. "↑" indicates interspecific competition becomes weaker or facilitation becomes stronger post-mortality. "↓↑" indicates interspecific competition becomes stronger or facilitation becomes weaker on some scales, and interspecific competition becomes weaker or facilitation becomes stronger on some other scales post-mortality.“-”表示物种之间存在竞争, “+”表示物种之间存在互助, “-+”表示物种之间在部分尺度上存在竞争, 在其他一些尺度上存在互助。“/”表示物种之间无明显变化, 或无明显相关性。“↓”表示死亡后种间竞争增强或互助减弱; “↑”表示死亡后种间竞争减弱或互助增强; “↓↑”表示死亡后种间关系在部分尺度上竞争增强或互助减弱, 在其他一些尺度上竞争减弱或互助增强。 新窗口打开 此外, 从种间相关性百分比(图3)来看, 物种之间的相关性以不相关为主, 并且在大部分尺度上表现为正相关的物种对数多于负相关, 仅在大于20 m的尺度上负相关的物种对数多于正相关。死亡前后种间相关性百分比变化较小, 仅在个别尺度有较大变化: 死亡后在2 m左右的尺度上表现为正相关的物种对数明显增加, 不相关的物种对数减少; 在8-15 m的尺度上, 表现出负相关的物种对数有所减少, 而不相关的物种对数增多; 在大于22 m的尺度上, 表现为正相关的物种对数有所减少, 而不相关的物种对数增多。 显示原图|下载原图ZIP|生成PPT 图3个体死亡前(A)和死亡后(B)锐齿槲栎与其他物种种间相关性的百分比。 -->Fig. 3The percentage of interspecific spatial associations between Quercus aliena var. acuteserrata and other species before individual death (A) and after individual death (B). -->
AakalaT, FraverS, PalikBJ, D AmatoAW (2012). Spatially random mortality in old-growth red pine forests of northern Minnesota .Canadian Journal of Forest Research, 42, 899-907. [本文引用: 1]
[2]
BaddeleyA, TurnerR (2005). Spatstat: An R package for analyzing spatial point patterns .Journal of Statistical Software, 12(i06), 1-42. [本文引用: 1]
[3]
BinY, LianJ, WangZ, YeW, CaoH (2011). Tree mortality and recruitment in a subtropical broadleaved monsoon forest in South China .Journal of Tropical Forest Science, 23(1), 57-66. [本文引用: 2]
[4]
ChenY, YuanZL, RenSY, WeiBL, JiaHR, YeYZ (2014). Correlation analysis of soil and species of different life forms in Baotianman Nature Reserve .Chinese Science Bulletin, 59, 2367-2376. (in Chinese)[陈云, 袁志良, 任思远, 韦博良, 贾宏汝, 叶永忠 (2014). 宝天曼自然保护区不同生活型物种与土壤相关性分析 . 科学通报, 59, 2367-2376.] [本文引用: 1]
[5]
DiggleP (2013). Statistical Analysis of Spatial and Spatio-Temporal Point Patterns. CRC Press, Boca Raton, USA. [本文引用: 1]
[6]
DomkeGM, WoodallCW, SmithJE (2011). Accounting for density reduction and structural loss in standing dead trees: Implications for forest biomass and carbon stock estimates in the United States .Carbon Balance & Management, 6(1), 57-62. [本文引用: 1]
[7]
FanJ, ZhaoXH, WangJS, ZhangCY, HeJ, XiaFC (2012). Spatial patterns of dominant species in a subtropical evergreen broad-leaved forest in Jiulian Mountain Jiangxi Province, China .Acta Ecologica Sinica, 32, 2729-2737. (in Chinese with English abstract)[范娟, 赵秀海, 汪金松, 张春雨, 何俊, 夏富才 (2012). 江西九连山亚热带常绿阔叶林优势种空间分布格局 . 生态学报, 32, 2729-2737.] [本文引用: 1]
[8]
GetzinS, DeanC, HeF, TrofymowJA, WiegandK, WiegandT (2006). Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island .Ecography, 29, 671-682. [本文引用: 1]
[9]
GrayL, HeF (2009). Spatial point-pattern analysis for detecting density-dependent competition in a boreal chronosequence of Alberta .Forest Ecology & Management, 259, 98-106. [本文引用: 3]
[10]
GuoYL, WangB, XiangWS, DingT, LuSH, HuangFZ, WenSJ, LiDX, LiXK (2016). Spatial distribution and habitat-association of snags in the tropical karst seasonal rainforest of Southwest Guangxi, China .Guihaia, 36, 154-161. (in Chinese with English abstract)[郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄甫昭, 文淑均, 李冬兴, 李先琨 (2016). 桂西南喀斯特季节性雨林枯立木的空间格局及生境关联性分析 . 广西植物, 36, 154-161.] [本文引用: 2]
[11]
HeF, DuncanRP (2000). Density-dependent effects on tree survival in an old-growth Douglas fir forest .Journal of Ecology, 88, 676-688. [本文引用: 4]
[12]
HeJS, LiuF, ChenWL, ChenLZ (1999). History of disturbance and regeneration strategies of Fagus engleriana and Quercus aliena var. acuteserrata forests in Shennongjia, Hubei Province . Acta Botanica Sinica, 41, 887-892. (in Chinese with English abstract)[贺金生, 刘峰, 陈伟烈, 陈灵芝 (1999). 神农架地区米心水青冈林和锐齿槲栎林群落干扰历史及更新策略 . 植物学报, 41, 887-892.]
[13]
HouHY, WangLH (2013). Species composition and main populations spatial distribution pattern in Korean pine broadleaved forest in Xiaoxing’an Mountains of Northeast China .Chinese Journal of Applied Ecology, 24, 3043-3049. (in Chinese with English abstract)[侯红亚, 王立海 (2013). 小兴安岭阔叶红松林物种组成及主要种群的空间分布格局 . 应用生态学报, 24, 3043-3049.] [本文引用: 2]
[14]
LiL, WuCC, MoDQ, WeiSG, HuangZL (2014). Dynamics death of constructive species in monsoon evergreen broad leaved forest .Journal of Southwest Forestry University, 34(6), 29-34. (in Chinese with English abstract)[李林, 伍婵翠, 莫德清, 魏识广, 黄忠良 (2014). 季风常绿阔叶林建群种的死亡动态格局 . 西南林业大学学报, 34(6), 29-34.] [本文引用: 2]
[15]
LinY, RenJY, YueM (2008). Population structure and spatial analysis of Betula albo-sinensis at Taibai Mountain, Northwestern China .Journal of Plant Ecology (Chinese Version), 32, 1335-1345. (in Chinese with English abstract)[林玥, 任坚毅, 岳明 (2008). 太白山红桦种群结构与空间分析 . 植物生态学报, 32, 1335-1345.] [本文引用: 1]
[16]
LiuF, ChenWL, HeJS (2000). Population structure and regeneration of Quercus aliena var. acuteserrata in Shennongjia .Acta Phytoecologica Sinica, 24, 396-401.(in Chinese with English abstract) [刘峰, 陈伟烈, 贺金生 (2000). 神农架地区锐齿槲栎种群结构与更新的研究 . 植物生态学报, 24, 396-401.]
[17]
LiuYC, WuMZ, GuoZM, JiangYX, LiuSR, WangZY, LiuBD, ZhuXL (2001). Studies on biomass and net production of Quercus acutidentata forest in Baotianman Nature Reserve .Acta Ecologica Sinica, 21, 1450-1456. (in Chinese with English abstract)[刘玉萃, 吴明作, 郭宗民, 蒋有绪, 刘世荣, 王正用, 刘保东, 朱学凌 (2001). 内乡宝天曼自然保护区锐齿栎林生物量和净生产力研究 . 生态学报, 21, 1450-1456.] [本文引用: 1]
[18]
LiuYY, JinGZ (2010). Spatial point pattern analysis for coarse woody debris in a mixed broadleaved-Korean pine forest in Xiaoxing’an Mountains, China .Acta Ecologica Sinica, 30, 6072-6081. (in Chinese with English abstract)[刘妍妍, 金光泽 (2010). 小兴安岭阔叶红松林粗木质残体空间分布的点格局分析 . 生态学报, 30, 6072-6081.] [本文引用: 1]
[19]
LuZJ, LiuFL, WuH, JiangMX (2015). Species composition, size class, and spatial patterns of snags in the Badagongshan (BDGS) mixed evergreen and deciduous broad- leaved forest in central China .Biodiversity Science, 23, 167-173. (in Chinese with English abstract)[卢志军, 刘福玲, 吴浩, 江明喜 (2015). 八大公山常绿落叶阔叶混交林枯立木物种组成、大小级与分布格局 . 生物多样性, 23, 167-173.] [本文引用: 1]
[20]
MetcalfCJE, HorvitzCC, TuljapurkarS, ClarkDA (2009). A time to grow and a time to die: A new way to analyze the dynamics of size, light, age, and death of tropical trees .Ecology, 90, 2766-2778. [本文引用: 1]
[21]
MiaoN, LiuS, ShiZ, XueZ, YuH, HeF (2011). Spatial pattern analysis of a Rhododendron-Abies virginal forest near timberline on the eastern edge of Qinghai-Tibetan Plateau, China .Acta Ecologica Sinica, 31, 1-9. (in Chinese with English abstract)[缪宁, 刘世荣, 史作民, 薛泽冰, 喻泓, 何飞 (2011). 青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局 . 生态学报, 31, 1-9.] [本文引用: 2]
[22]
MiaoN, LiuSR, ShiZM, YuH, LiuXL (2009). Spatial patterns of dominant tree species in sub-alpine Betula-Abies forest in West Sichuan of China .Chinese Journal of Applied Ecology, 20, 1263-1270. (in Chinese with English abstract)[缪宁, 刘世荣, 史作民, 喻泓, 刘兴良 (2009). 川西亚高山红桦-岷江冷杉林优势种群的空间格局分析 . 应用生态学报, 20, 1263-1270.] [本文引用: 3]
[23]
NongY, ZhengL, JiaHY, LuLH, HuangDW, HuangBH, LeiLQ (2015). Community characteristics and spatial distribution of dominant tree species in a secondary forest of Daqing Mountains, southwestern Guangxi, China .Biodiversity Science, 23, 321-331. (in Chinese with English abstract)[农友, 郑路, 贾宏炎, 卢立华, 黄德卫, 黄柏华, 雷丽群 (2015). 广西大青山次生林的群落特征及主要乔木种群的空间分布格局 . 生物多样性, 23, 321-331.] [本文引用: 1]
[24]
RouvinenS, KuuluvainenT, SiitonenJ (2002). Tree mortality in a Pinus sylvestris dominated boreal forest landscape in Vienansalo wilderness, eastern Fennoscandia .Silva Fennica, 36, 127-145. [本文引用: 1]
[25]
SvátekM, MatulaR (2015). Fine-scale spatial patterns in oak sprouting and mortality in a newly restored coppice .Forest Ecology & Management, 348, 117-123. [本文引用: 1]
[26]
WangB, XiangWS, DingT, HuangFZ, WenSJ, LiDX, GuoYL, LiXK (2014). Spatial distribution of standing dead trees abundance and its impact factors in the karst seasonal rain forest, Nonggang, southern China .Chinese Science Bulletin, 59, 3479-3490. (in Chinese)[王斌, 向悟生, 丁涛, 黄甫昭, 文淑均, 李冬兴, 郭屹立, 李先琨 (2014). 弄岗喀斯特季节性雨林枯立木多度的空间分布及影响因子 . 科学通报, 59, 3479-3490.] [本文引用: 2]
[27]
WangLW, LiBH, YeJ, BaiXJ, YuanZQ, XingDL, LinF, ShiS, WangXG, HaoZQ (2011). Dynamics of short-term tree mortality in broad-leaved Korean pine mixed forest in the Changbai Mountains .Biodiversity Science, 19, 260-270. (in Chinese with English abstract)[王利伟, 李步杭, 叶吉, 白雪娇, 原作强, 邢丁亮, 蔺菲, 师帅, 王绪高, 郝占庆 (2011). 长白山阔叶红松林树木短期死亡动态 . 生物多样性, 19, 260-270.] [本文引用: 2]
[28]
WiegandT, GunatillekeS, GunatillekeN (2007). Species associations in a heterogeneous Sri Lankan dipterocarp forest .The American Naturalist, 170, E77-E95. [本文引用: 1]
[29]
WiegandT, MoloneyAK (2004). Rings, circles, and null- models for point pattern analysis in ecology .Oikos, 104, 209-229. [本文引用: 2]
[30]
XuJ, FeiSM, HeYP, JiangJM, ChenXM, TuDL (2007). The competition intensity between withered and living standing trees in the upper layer of overmature Pinus yunnanensis forest in the mountainous area of Southwest Sichuan. Journal of Sichuan Forestry Science & Technology, 28(6), 1-8. (in Chinese with English abstract)[徐嘉, 费世民, 何亚平, 蒋俊明, 陈秀明, 涂代伦 (2007). 川西南山地云南松过熟林上层枯立木和活立木竞争强度研究 . 四川林业科技, 28(6), 1-8.] [本文引用: 1]
[31]
YuanZL, WangT, ZhuXL, ShaYY, YeYZ (2011). Patterns of spatial distribution of Quercus variabilis in deciduous broadleaf forests in Baotianman Nature Reserve .Biodiversity Science, 19, 224-231. (in Chinese with English abstract)[袁志良, 王婷, 朱学灵, 沙迎迎, 叶永忠 (2011). 宝天曼落叶阔叶林样地栓皮栎种群空间格局 . 生物多样性, 19, 224-231.] [本文引用: 2]
[32]
ZhangJ, HaoZ, SunIF, SongB, YeJ, LiB, WangX (2009). Density dependence on tree survival in an old-growth temperate forest in northeastern China .Annals of Forest Science, 66, 204. [本文引用: 3]
[33]
ZhangZH, HuG, NiJ (2010). Interspecific segregation of old-growth karst forests in Maolan, Southwest China .Acta Ecologica Sinica, 30, 2235-2245. (in Chinese with English abstract)[张忠华, 胡刚, 倪健 (2010). 茂兰喀斯特森林群落的种间分离特征 . 生态学报, 30, 2235-2245.] [本文引用: 1]
Spatially random mortality in old-growth red pine forests of northern Minnesota 1 2012
Accounting for density reduction and structural loss in standing dead trees: Implications for forest biomass and carbon stock estimates in the United States 1 2011
... 树木死亡是森林群落动态的重要环节, 它对群落内物种组成、群落环境、森林生态系统物质循环以及物种的空间分布有较大影响(Domke et al., 2011; 王利伟等, 2011; 郭屹立等, 2016), 并在群落更新及演替过程中发挥着重要作用(Rouvinen et al., 2002).树木死亡的原因很多, 例如树木衰老、自然灾害、病害虫害、动物取食以及个体之间的竞争等(Metcalf et al., 2009; 卢志军等, 2015). ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT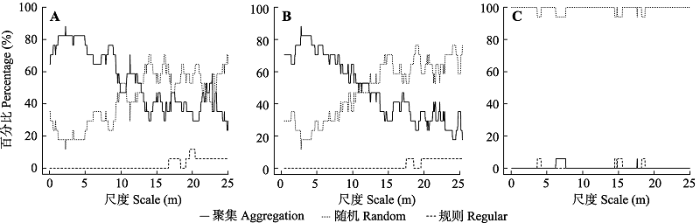 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT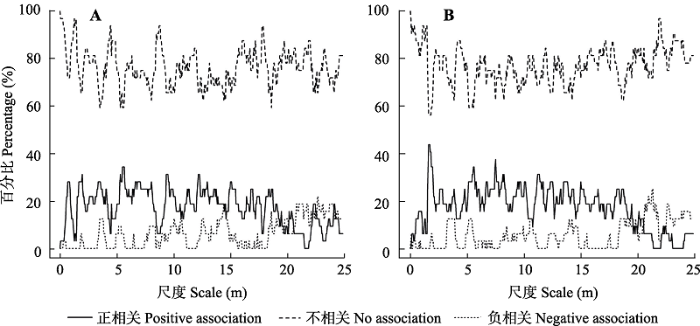 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}