Responses of apical ion fluxes to NaCl stress in Elaeagnus angustifolia seedlings
YANGSheng1,2,*, ZHANGHua-Xin2, CHENQiu-Xia1, YANGXiu-Yan2 1 Zhejiang Institute of Subtropical Crops, Wenzhou, Zhejiang 325005, Chinaand 2Research Center of Saline and Alkali Land of State Forestry Administration, Beijing 100091, China 责任编辑:YANGShengZHANGHua-XinCHENQiu-XiaYANGXiu-Yan 收稿日期:2016-03-9 接受日期:2016-09-21 网络出版日期:2017-04-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家“十二五”科技支撑计划(2011BAD38B0102)和中央公益科研院所基本科研业务费专项资金(CAFYBB2012009)
关键词:沙枣;NaCl胁迫;离子流;种源;Na+/H+逆向转运体;K+/Na+平衡 Abstract AimsElaeagnus angustifolia is one of the most salt-tolerant species. The objective of this study was to understand the mechanisms of ion transporation in E. angustifolia exposed to different salt concentrations through manipulations of K+/Na+ homeostasis. Methods Seedlings of two variants of the species, Yinchuan provenance (YC, salt-sensitive type) and the Alaer provenance (ALE, salt-tolerant type), were treated with three different NaCl application modes, and the ion fluxes in the apical regions were measured using non-invasive micro-test technology (NMT). In mode 1, Na+ and K+ fluxes were measured after 150 mmol·L-1 NaCl stress lasted for 24 h. In mode 2, K+ and H+ fluxes were quantified with a transient stimulation of NaCl solution. In mode 3, Amiloride (Na+/H+ antiporters inhibitor) and tetraethylammonium (TEA, K+ channel inhibitor) was used to treat apical regions of E. angustifolia seedlings after NaCl stress for 24 h, respectively. Important findings Under NaCl stress for 24 h, net effluxes of Na+ and K+ were increased significantly. The net Na+ effluxes of YC provenance seedlings (720 pmol·cm-2•s-1) were lower than that of ALE provenance (912 pmol·cm-2·s-1), but the net K+ efflux was higher in YC provenance. Under the instantaneous NaCl stimulation, net K+ efflux was remarkably increased, with the net K+ efflux of YC provenance always higher than that of ALE provenance. Interestingly, H+ at the apical regions was found from influx to efflux, with the net H+ efflux of ALE provenance greater than that of the YC provenance. Under the NaCl and NaCl + Amiloride treatment, the net Na+ efflux of ALE provenance seedlings was higher than that of YC provenance, while the net K+ efflux was less in ALE provenance seedlings. On the other hand, the differences in net Na+ and K+ effluxes were insignificant between the two provenances under the control group and NaCl + TEA treatment. In conclusion, NaCl stress caused Na+ accumulation and K+ outflows of E. angustifolia seedlings; The E. angustifolia seedlings utilize Na+/H+ antiporters to reduce Na+ accumulation by excretion; and the maintenance of K+/Na+ homeostasis in salt-tolerant E. angustifolia provenance seedlings roots accounted for a greater Na+ extrusion and a lower K+ efflux under NaCl stress. Results from this study provide a theoretical basis for further exploring salt-tolerant E. angustifolia germplasm resource.
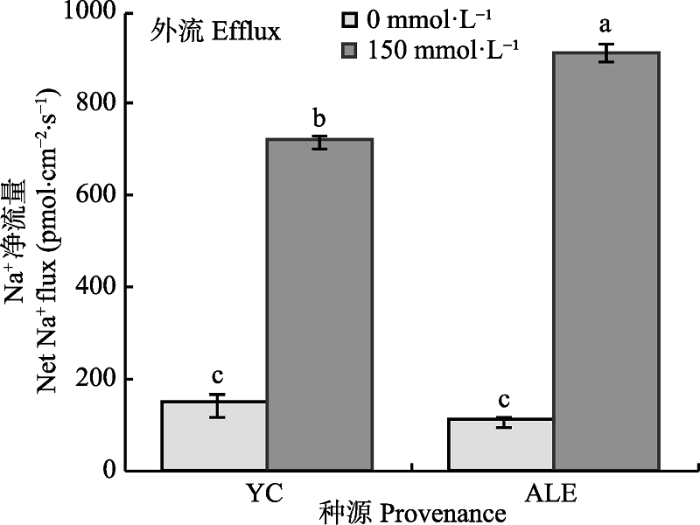
2.1.1 稳态Na+流 从图1可以看出, 在对照条件下, 两个种源沙枣幼苗根系的Na+流均为外流, 其净流量均低于 200 pmol·cm-2·s-1, 两个种源间无显著性差异, 而在150 mmol·L-1 NaCl胁迫处理24 h后, 与对照相比, 沙枣根系的Na+外流的净流量显著增加, 并且两个种源间差异明显。NaCl胁迫下, 阿拉尔种源根系Na+外流的净流量显著高于银川种源, 分别为912和720 pmol·cm-2·s-1。由此说明, 盐胁迫下, 沙枣幼苗根系Na+的外排能力显著提高, 并且与银川种源相比, 阿拉尔种源沙枣幼苗表现出更强的排Na+能力。 显示原图|下载原图ZIP|生成PPT 图1NaCl胁迫对两个种源沙枣幼苗根系稳态Na+流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示不同处理和种源间差异显著(p < 0.05)。 -->Fig. 1Effects of NaCl stress on steady-state Na+ flux at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences between different treatments and provenances (p < 0.05). -->
2.1.2 稳态K+流 K+是植物代谢过程中的重要物质, 限制体内Na+积累, 减少体内K+流失, 可以提高植物耐盐性(Gouiaa et al., 2012)。如图2所示, 在对照条件下, 银川和阿拉尔两个种源沙枣根系在对照条件下K+外流的净流量仅分别为180和159 pmol·cm-2·s-1, 而在150 mmol·L-1 NaCl胁迫下, 两个种源沙枣根系K+外流的净流量显著增大, 分别为469和360 pmol·cm-2·s-1,并且银川种源沙枣幼苗K+外流净流量明显高于阿拉尔种源, 这表明NaCl胁迫能够激发沙枣根系中K+外流, 并且银川种源沙枣幼苗根系保留K+能力弱于阿拉尔种源。 显示原图|下载原图ZIP|生成PPT 图2NaCl胁迫对两个种源沙枣幼苗根系稳态K+流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示不同处理和种源间差异显著(p < 0.05)。 -->Fig. 2Effects of NaCl stress on steady-state K+ flux at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences between different treatments and provenances (p < 0.05). -->
2.2 NaCl胁迫对不同种源沙枣幼苗根系离子流动力学的影响
2.2.1 瞬时K+流的动态特性 由图3可知, NaCl胁迫前, 两个种源沙枣幼苗根系K+流表现出平稳的轻微外流。在瞬时加入NaCl (150 mmol·L-1)后, 沙枣幼苗根系K+的外流立刻显著增加, 但是外排强度和变化趋势种源间存在差异。加入NaCl后, 银川种源沙枣幼苗根系的K+外排始终高于阿拉尔种源, 并且在10-17 min间, 银川种源K+的外流逐渐增加, 其最大值为701 pmol·cm-2·s-1, 随后呈缓慢下降趋势, 而NaCl诱导阿拉尔种源K+外排在前5 min内呈现出下降趋势, 最终维持在200- 320 pmol·cm-2·s-1。这说明在受到NaCl刺激后, 沙枣幼苗根系的K+外流立刻显著增加, 引起K+损失, 并且阿拉尔种源沙枣幼苗根系的保留K+能力强于银川种源。 显示原图|下载原图ZIP|生成PPT 图3NaCl胁迫诱导两个种源沙枣幼苗根系瞬时K+流的动态差异(平均值±标准误差, n = 4)。不同小写字母表示同一时间点种源间差异显著(p < 0.05)。 -->Fig. 3Differences of NaCl stress on transient K+ kinetics at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). Different lowercase letters indicate significant differences between provenances at the same time (p < 0.05). -->
2.2.2 瞬时H+流动态特性 由图4可知, 两个种源沙枣幼苗根系的H+净流量在NaCl胁迫前呈现出轻微的内流, 但NaCl胁迫诱导后, H+流马上由内流转为外流, 并且阿拉尔种源的H+外流大于银川种源。随着时间的延长, 沙枣幼苗根系的H+净流量逐渐下降, 大约胁迫15 min后, H+净流量趋于平稳, 但阿拉尔种源根系维持在6.38-9.41 pmol·cm-2·s-1之间, 高于银川种源(2.21- 5.50 pmol·cm-2·s-1), 并且随着时间延长, 差异越来越明显。由此说明, NaCl胁迫能够诱导沙枣根系的H+外流, 并且耐盐型沙枣种源的外流量大于盐敏感型种源, 进而表明耐盐型沙枣种源根系具有更强的质子泵活性, 由此抑制膜电位去极化, 减少K+流失。 显示原图|下载原图ZIP|生成PPT 图4NaCl胁迫诱导两个种源沙枣幼苗根系瞬时H+流的动态差异(平均值±标准误差, n = 4)。不同小写字母表示同一时间点种源间差异显著(p < 0.05)。 -->Fig. 4Differences of NaCl stress on transient H+ kinetics at apical regions of two provenances of Elaeagnus angustifolia (mean ± SE, n = 4). Different lowercase letters indicate significant differences between provenances at the same time (p < 0.05). -->
2.3 NaCl胁迫下两个种源沙枣幼苗根系离子流的药理学特性
2.3.1 两个种源沙枣幼苗根系在不同处理下的Na+流 如图5所示, 银川种源和阿拉尔种源沙枣幼苗根系在各处理下, Na+净流量均为外流, 在对照条件下, 外流量最小, 而NaCl胁迫下最大。在TEA和Amiloride处理后, Na+的净流量均显著减少。同时, Amiloride作为Na+/H+逆向转运蛋白抑制剂, 对Na+外流的限制作用比K+通道蛋白抑制剂TEA显著。在NaCl胁迫下, 阿拉尔种源的Na+净流量处于1β000-1β200 pmol·cm-2·s-1之间, 而银川种源则维持在800-1β000 pmol·cm-2·s-1。在NaCl + Amiloride处理后, 银川种源的Na+净外流量仍然显著低于阿拉尔种源, 与仅NaCl处理相比, 分别下降了 449 pmol·cm-2·s-1和429 pmol·cm-2·s-1。在NaCl + TEA处理后, 沙枣幼苗根系的Na+净流量在阿拉尔和银川种源间差异不明显, 分别为775和727 pmol·cm-2·s-1。进一步说明在NaCl胁迫下, 阿拉尔种源沙枣幼苗质膜Na+外排能力强于银川种源。 显示原图|下载原图ZIP|生成PPT 图5两种抑制剂对NaCl胁迫后两个种源沙枣根系Na+流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示同一种源不同处理间差异显著(p < 0.05); 不同大写字母表示相同处理不同种源间差异显著(p < 0.05)。 -->Fig. 5Effects of two kinds of inhibitors on net Na+ flux at apical regions of two provenances of Elaeagnus angustifolia treated by NaCl stress (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences among different treatments in the same provenance (p < 0.05), While different capital letters indicate significant difference between provenances at the same treatment (p < 0.05). -->
2.3.2 两个种源沙枣幼苗根系在不同处理下的K+流 如图6所示, 在各处理下, 银川和阿拉尔种源沙枣幼苗根系的K+净流量均呈现出外流, K+净流量在对照条件下分别仅为117和87 pmol·cm-2·s-1, 当NaCl胁迫24 h后, 分别增加到859和560 pmol·cm-2·s-1。当加入抑制剂处理后, K+的外流均显著减少, 而Amiloride的抑制效果弱于TEA。在NaCl和NaCl+ Amiloride处理条件下, 银川种源沙枣幼苗根尖的K+净流量均明显高于阿拉尔种源, 而在CK和NaCl+ TEA处理下, 两个种源间无明显差异。表明NaCl胁迫环境可以引起沙枣体内K+的流失, 银川种源沙枣幼苗根系保留K+的能力弱于阿拉尔种源, 并且K+通道蛋白和Na+/H+逆向转运体活性能影响质膜上的K+运移。 显示原图|下载原图ZIP|生成PPT 图6两种抑制剂对NaCl胁迫后两个种源沙枣根系K+流的影响(平均值±标准误差, n = 4)。ALE, 阿拉尔; YC, 银川。不同小写字母表示同一种源不同处理间差异显著(p < 0.05); 不同大写字母表示相同处理不同种源间差异显著(p < 0.05)。 -->Fig. 6Effects of two kinds of inhibitors on net K+ flux at apical regions of two provenances of Elaeagnus angustifolia treated by NaCl stress (mean ± SE, n = 4). ALE, Alaer; YC, Yinchuan. Different lowercase letters indicate significant differences among different treatments in the same provenance (p < 0.05), While different capital letters indicate significant difference between provenances at the same treatment (p < 0.05). -->
3 讨论
植物维持细胞溶质中较高的K+/Na+是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+吸收, 增加Na+外排和区隔化, 以及控制K+外流量来实现较高的K+/Na+ (Maathuis & Amtmann, 1999; Chen et al., 2007a; Cuin et al., 2012)。有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+能力或保留K+能力(Chen et al., 2007b, 2008; Cuin et al., 2008; Sun et al., 2009)。但是也有研究显示K+外流不能预测不同基因型水稻(Oryza sativa)的耐盐性(Coskun et al., 2013)。在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+和K+外流量均显著增加, 并且耐盐型沙枣种源的Na+净流量明显大于盐敏感型种源, 而K+外流的净流量小于盐敏感型种源。这表明NaCl胁迫引起沙枣幼苗Na+积累而K+流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+外排能力和保留K+能力。 在高等植物中, 质膜Na+/H+逆向转运蛋白除了负责调节根部Na+外排和体内Na+长距离运输外, 还可以调控细胞内的K+平衡、pH值和Ca2+信号转导 (马清等, 2011; 孙健, 2011)。质膜Na+/H+逆向转运体在盐胁迫下被诱导合成或者激活, 进而将Na+从细胞质外排到细胞外, 但是排Na+需要质膜两侧产生的H+电化学势梯度来完成, 而质膜H+-ATPase利用水解ATP产生的能量将H+泵出细胞质能形成H+电化学势梯度(Rausch et al., 1996; 陈观平等, 2006; 马清等, 2011)。同时, Olias等(2009)对番茄(Solanum lycopersicum)研究发现质膜Na+/H+逆向转运体(SlSOS1)不仅在NaCl胁迫下维持离子平衡起重要作用, 而且对于Na+在各器官间的区隔化也发挥关键作用。Cuin等(2011)研究认为Na+/H+逆向转运体可以作为小麦耐盐性筛选的一个量化指标。大量转基因试验证实质膜Na+/H+逆向转运体的转基因植物也都表现出更强的耐盐能力(Yang et al., 2009; Yue et al., 2012; Feki et al., 2014)。在本试验中, NaCl胁迫下, 沙枣幼苗根系Na+外排显著增加, 耐盐型沙枣种源的Na+外流速率大于盐敏感型种源, 并且瞬时盐胁迫下, H+由内流变为外流, 耐盐型种源沙枣幼苗根系的H+外流量明显大于盐敏感型种源, 说明耐盐型种源沙枣幼苗在盐胁迫下具有更强的Na+/H+逆向转运体和质膜H+-ATPase活性。本研究的药理学试验显示, Na+/H+逆向转运体抑制剂Amiloride显著减少Na+的外排, 并且阿拉尔种源在NaCl和NaCl+ Amiloride处理下Na+净外排量均显著高于银川种源, 这进一步表明质膜Na+/H+逆向转运蛋白在沙枣根尖外排Na+过程中起着非常重要的作用, 但是两个种源在抑制量间无显著差异(449 pmol·cm-2·s-1和 429 pmol·cm-2·s-1), 并且在NaCl + TEA处理后两个种源根尖的Na+净流量也无明显差异, 这可能与抑制剂浓度, 处理时间和试验材料有关, 也可能沙枣的排Na+机制有其未知的特殊性,有待进一步研究探讨。 另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+通道和非选择性阳离子通道外流的K+增加(Chen et al., 2007a; Shabala & Cuin, 2008; Sun et al., 2009)。在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+外流量显著增加, 但受到K+通道抑制剂TEA的限制, 这与对大麦(Chen et al., 2007a)的研究结论相同。另外, 在TEA处理后, 两个种源间的K+外流差异不明显, 由此推测两个种源的K+外流在其他通道途径间无差异, 经K+通道而发生的K+外流是引起种源间K+外流和保持K+/Na+平衡能力差异的一个主要原因。 综上所述, 在NaCl胁迫环境下, 沙枣幼苗为了降低体内Na+的积累, 通过质膜Na+/H+逆向转运体将Na+排到细胞外, 同时, NaCl胁迫会使细胞去极化程度加强, 增加K+外流。阿拉尔种源的沙枣幼苗根系表现出更强的Na+/H+逆向转运体活性, 更多的Na+外排, 从而减少Na+向叶片组织的运输, 并减少NaCl胁迫下根系细胞中的K+外排, 进而更好地保持细胞质中K+/Na+平衡, 由此表现出更强的耐盐能力。 致谢 感谢温州市森林生态科技创新团队项目(C20150002)资助。 The authors have declared that no competing interests exist. 作者声明没有竞争性利益冲突.
ChenGP, WangHZ, ShiNN, ChenSY (2006). Na+/H+ antiporter and its relationship with plant salt tolerance .China Biotechnology, 26(5), 101-106. (in Chinese with English abstract)[陈观平, 王慧中, 施农农, 陈受宜 (2006). Na+/H+逆向转运蛋白与植物耐盐性的关系研究进展 . 中国生物工程杂志, 26(5), 101-106.] [本文引用: 1]
[4]
ChenZ, NewmanI, ZhouM, MendhamN, ZhangG, ShabalaS (2005). Screening plants for salt tolerance by measuring K+ flux: A case study for barely .Plant, Cell & Environment, 28, 1230-1246. [本文引用: 1]
ChenZ, ShabalaS, MendhamN, NewmanI, ZhangGP, ZhouMX (2008). Combining ability of salinity tolerance on the basis of NaCl-induced K flux from roots of barley .Crop Science, 48, 1382-1388. [本文引用: 1]
[7]
ChenZ, ZhouM, NewmanIA, MendhamNJ, ZhangGP, ShabalaS (2007b). Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance .Functional Plant Biology, 34, 150-162. [本文引用: 1]
[8]
CoskunD, BrittoDT, JeanYK, KabirI, TolayI, TorunAA, KronzuckerHJ (2013). K+ efflux and retention in response to NaCl stress do not predict salt tolerance in contrasting genotypes of rice (Oryza sativa L.) .PLOS ONE, 8(2), e57767. doi: 10.1371/journal.pone.0057767. [本文引用: 1]
[9]
CuinTA, BettsSA, ChalmandrierR, ShabalaS (2008). A root’s ability to retain K+ correlates with salt tolerance in wheat .Journal of Experimental Botany, 59, 2697-2706. [本文引用: 2]
[10]
CuinTA, BoseJ, StefanoG, JhaD, TesterM, MancusoS, ShabalaS (2011). Assessing the role of root plasma membrane and tonoplast Na+/H+ exchangers in salinity tolerance in wheat: In planta quantification methods .Plant, Cell & Environment, 34, 947-961.
DemidchikV, MaathuisFJM (2007). Physiological roles of nonselective cation channels in plants: From salt stress to signaling and development .New Phytologist, 175, 384-404. [本文引用: 1]
[13]
FekiK, QuinteroFJ, KhoudiH, LeidiEO, MasmoudiK, PardoJM, BriniF (2014). A constitutively active form of a durum wheat Na+/H+ antiporter SOS1 confers high salt tolerance to transgenic Arabidopsis . Plant Cell Reportsl Reports, 33, 277-288. [本文引用: 2]
[14]
GouiaaS, KhoudiH, LeidiEO, PardoJM, MasmoudiK (2012). Expression of wheat Na+/H+ antiporter TNHXS1 and H+-pyrophosphatase TVP1 genes in tobacco from a bicistronic transcriptional unit improves salt tolerance . Plant Molecular Biology, 79, 137-155. [本文引用: 1]
[15]
GuanWK, XuN (2012). Research situation and resources utilization of Elaeagnus angustifolia . Anhui Agricultural Science Bulletin, 18(19), 119-121. (in Chinese with English abstract)[管文轲, 徐娜 (2012). 沙枣资源利用研究与开发现状述评 . 安徽农学通报, 18(19), 119-121.] [本文引用: 1]
[16]
GuoLJ, WangYT (2008). Conservation research and prospects of Elaeagnus germplasm resources and utilization values .Chinese Wild Plant Resources, 27(5), 32-34. (in Chinese with English abstract)[郭丽君, 王玉涛 (2008). 沙枣种质资源特性及利用价值 . 中国野生植物资源, 27(5), 32-34.] [本文引用: 1]
[17]
KongX, LuoZ, DongH, EnejiAE, LiW (2012). Effects of non-uniform root zone salinity on water use, Na+ recirculation, and Na+ and H+ flux in cotton .Journal of Experimental Botany, 63, 2105-2116. [本文引用: 3]
[18]
LiuBY (2007). Study on Ecophysiological Response and Ion Distribution of Elaeagnus angustifolia to Salt Stress . Master degree dissertation, Tianjin Normal University, Tianjin. 27-53. (in Chinese with English abstract)[刘宝玉 (2007). 盐胁迫下沙枣生理生态响应与离子分配研究 . 硕士学位论文, 天津师范大学, 天津. 27-53.] [本文引用: 1]
[19]
LiuZX (2013). Physiological Mechanism of Heterogeneous Responses of Elaeagnus angustifolia to NaCl and Na2SO4 Stress . PhD dissertation, Chinese Academy of Forestry, Beijing. 100-104. (in Chinese with English abstract)[刘正祥 (2013). 沙枣对氯化钠和硫酸钠胁迫异质性响应的生理机制 . 博士学位论文, 中国林业科学研究院, 北京. 100-104.] [本文引用: 3]
[20]
LiuZX, ZhangHX, YangXY, LiuT, DiWB (2014). Growth, and cationic absorption, transportation and allocation of Elaeagnus angustifolia seedlings under NaCl stress .Acta Ecologica Sinica, 34, 326-336. (in Chinese with English abstract)[刘正祥, 张华新, 杨秀艳, 刘涛, 狄文彬 (2014). NaCl胁迫下沙枣幼苗生长和阳离子吸收、运输与分配特性 . 生态学报, 34, 326-336.] [本文引用: 1]
[21]
MaQ, BaoAK, WuGQ, WangSM (2011). Plasma membrane Na+/H+ antiporter is involved in plant salt tolerance .Chinese Bulletin of Botany, 46, 206-215. (in Chinese with English abstract)[马清, 包爱科, 伍国强, 王锁民 (2011). 质膜Na+/H+逆向转运蛋白与植物耐盐性 . 植物学报, 46, 206-215.] [本文引用: 2]
[22]
MaathuisFJM (2006). The role of monovalent cation transporters in plant responses to salinity .Journal of Experimental Botany, 57, 1137-1147. [本文引用: 1]
[23]
MaathuisFJM, AmtmannA (1999). K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios .Annals of Botany, 84, 123-133. [本文引用: 1]
[24]
OliasR, EljakaouiZ, LiJU, MoralesPD, Marin-manzanoMC, PardoJM, BelverA (2009). The plasma membrane Na+/H+ antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+ between plant organs .Plant, Cell & Environment, 32, 904-916.
[25]
RauschT, KirschM, LöwR, LehrA, ViereckR, AnZG (1996). Salt stress responses of higher plants: The role of proton pumps and Na+/H+-antiporters .Journal of Plant Physiology, 148, 425-433. [本文引用: 1]
[26]
ShabalaS, CuinTA (2008). Potassium transport and plant salt tolerance .Physiologia Plantarum, 133, 651-669. [本文引用: 2]
[27]
ShiHZ, LeeB, WuSJ, ZhuJK (2003). Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana . Nature Biotechnology, 21, 81-85. [本文引用: 1]
[28]
ShiHZ, IshitaniM, KimC, ZhuJK (2000). The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter .Proceedings of the National Academy of Sciences of the United States of America, 97, 6896-6901. [本文引用: 1]
[29]
SunJ (2011). Signaling Network in the Perception of Salt Stress and Ionic Homeostasis Regulation in Populus euphratica . PhD dissertation, Beijing Forestry University, Beijing. 26-28. (in Chinese with English abstract)[孙健 (2011). 胡杨响应盐胁迫与离子平衡调控信号网络研究 . 博士学位论文, 北京林业大学, 北京. 26-28.] [本文引用: 4]
[30]
SunJ, ChenS, DaiS, WangR, LiN, ShenX, ZhouX, LuC, ZhengX, HuZ, ZhangZ, SongJ, XuY (2009). NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species .Plant Physiology, 149, 1141-1153. [本文引用: 2]
[31]
WegnerLH, StefanoG, ShabalaL, RossiM, MancusoM, ShabalaS (2011). Sequential depolarization of root cortical and stelar cells induced by an acute salt shock—Implications for Na+ and K+ transport into xylem vessels . Plant, Cell & Environment, 34, 859-869. [本文引用: 1]
[32]
YangQ, ChenZZ, ZhouXF, YinHB, LiX, XinXF, HongXH, ZhuJK, GongZ (2009). Overexpression of SOS (salt overly sensitive) genes increases salt tolerance in transgenic Arabidopsis . Molecular Plant, 2, 22-31. [本文引用: 1]
[33]
YangS, LiuT, ZhangHX, LiHY, ZhangL (2014). Growth and physiological characteristics of Elaeagnus angustifolia L. under salt stress .Journal of Fujian College of Forestry, 34(1), 64-70. (in Chinese with English abstract)[杨升, 刘涛, 张华新, 李焕勇, 张丽 (2014). 盐胁迫下沙枣幼苗的生长表现和生理特性 . 福建林学院学报, 34(1), 64-70.] [本文引用: 2]
[34]
YangS, ZhangHX, LiuT, WuHW, YangXY, NiJW, ChenQX (2016). Study on ion metabolism characteristics of Elaeagnus angustifolia L. seedlings under NaCl stress .Forest Research, 29(1), 140-146. (in Chinese with English abstract)[杨升, 张华新, 刘涛, 武海雯, 杨秀艳, 倪建伟, 陈秋夏 (2016). NaCl胁迫下沙枣幼苗的离子代谢特性 . 林业科学研究, 29(1), 140-146.] [本文引用: 2]
[35]
YangS, ZhangHX, YangXY, ChenQX, WuHW (2015). Differential growth performance ofElaeagnus angustifolia L. provenances under NaCl stress .Scientia Silvae Sinicae, 51(9), 51-58. (in Chinese with English abstract)[杨升, 张华新, 杨秀艳, 陈秋夏, 武海雯 (2015). NaCl胁迫下不同种源沙枣的生长表现差异 . 林业科学, 51(9), 51-58.] [本文引用: 1]
[36]
YueY, ZhangM, ZhangJ, DuanL, LiZ (2012). SOS1 gene overexpression increased salt tolerance in transgenic tobacco by maintaining a higher K+/Na+ ratio .Journal of Plant Physiology, 169, 255-261. [本文引用: 1]
[37]
ZhangBZ, CaoZY, ZhaoKF (1992). A study on some physiological properties ofElaeagnus angustifolia under salt stress condition .Scientia Silvae Sinicae, 28(2), 187-189. (in Chinese with English abstract)[张宝泽, 曹子谊, 赵可夫 (1992). 盐分胁迫下沙枣某些生理特性的研究 . 林业科学, 28(2), 187-189.] [本文引用: 1]
Cellular mechanisms of potassium transport in plants 1 2008
... 维持体内K+/Na+平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+的进入和增加Na+的外排或区隔化, 以减少体内或细胞质中的Na+浓度, 另一方面限制体内K+流失, 从而实现体内K+/Na+平衡(Chao et al., 2013; Feki et al., 2014).早期的研究结果显示细胞质中的Na+外排主要由质膜上的Na+/H+逆向转运蛋白(SOS1)负责, 并由H+-ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+的流入(Shi et al., 2000, 2003; Maathuis, 2006).另外, K+的吸收会受到Na+的竞争性抑制, 并且诱导通过外向K+通道而外流的K+增加(Demidchik & Maathuis, 2007; Britto & Kronzucker, 2008; Shabala & Cuin, 2008).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare)和小麦(Triticum aestivum)耐盐型品种根系的K+外流量均小于盐敏感型品种(Chen et al., 2005; Cuin et al., 2008).因此, 了解植物根系区域Na+和K+离子流情况, 对于理解植物耐盐能力非常重要. ...
Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis 1 2013
... 维持体内K+/Na+平衡是植物适应盐胁迫环境的一个关键手段, 利用先进技术手段和转基因技术, 人们对其机制开展了大量有益的探索研究, 发现植物一方面通过限制Na+的进入和增加Na+的外排或区隔化, 以减少体内或细胞质中的Na+浓度, 另一方面限制体内K+流失, 从而实现体内K+/Na+平衡(Chao et al., 2013; Feki et al., 2014).早期的研究结果显示细胞质中的Na+外排主要由质膜上的Na+/H+逆向转运蛋白(SOS1)负责, 并由H+-ATPase酶负责提供驱动力, 同时引起细胞膜电位重新极化, 减少Na+的流入(Shi et al., 2000, 2003; Maathuis, 2006).另外, K+的吸收会受到Na+的竞争性抑制, 并且诱导通过外向K+通道而外流的K+增加(Demidchik & Maathuis, 2007; Britto & Kronzucker, 2008; Shabala & Cuin, 2008).同时, 有研究显示在盐胁迫下大麦(Hordeum vulgare)和小麦(Triticum aestivum)耐盐型品种根系的K+外流量均小于盐敏感型品种(Chen et al., 2005; Cuin et al., 2008).因此, 了解植物根系区域Na+和K+离子流情况, 对于理解植物耐盐能力非常重要. ...
Expression of wheat Na+/H+ antiporter TNHXS1 and H+-pyrophosphatase TVP1 genes in tobacco from a bicistronic transcriptional unit improves salt tolerance 1 2012
NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species 2 2009
... 植物维持细胞溶质中较高的K+/Na+是评判植物耐盐性的一个重要因素, 植物主要通过限制Na+吸收, 增加Na+外排和区隔化, 以及控制K+外流量来实现较高的K+/Na+ (Maathuis & Amtmann, 1999; Chen et al., 2007a; Cuin et al., 2012).有研究发现, 耐盐性较强的品种在盐胁迫下具有更强的外排Na+能力或保留K+能力(Chen et al., 2007b, 2008; Cuin et al., 2008; Sun et al., 2009).但是也有研究显示K+外流不能预测不同基因型水稻(Oryza sativa)的耐盐性(Coskun et al., 2013).在本试验中, NaCl胁迫后, 沙枣幼苗根系的稳态Na+和K+外流量均显著增加, 并且耐盐型沙枣种源的Na+净流量明显大于盐敏感型种源, 而K+外流的净流量小于盐敏感型种源.这表明NaCl胁迫引起沙枣幼苗Na+积累而K+流失, 但耐盐型沙枣种源幼苗根系具有更好的Na+外排能力和保留K+能力. ...
... 另外, 在NaCl胁迫下, 细胞去极化程度会加强, 由此促使通过外向K+通道和非选择性阳离子通道外流的K+增加(Chen et al., 2007a; Shabala & Cuin, 2008; Sun et al., 2009).在本试验中, 在NaCl胁迫 24 h和瞬时NaCl胁迫后, K+外流量显著增加, 但受到K+通道抑制剂TEA的限制, 这与对大麦(Chen et al., 2007a)的研究结论相同.另外, 在TEA处理后, 两个种源间的K+外流差异不明显, 由此推测两个种源的K+外流在其他通道途径间无差异, 经K+通道而发生的K+外流是引起种源间K+外流和保持K+/Na+平衡能力差异的一个主要原因. ...
Sequential depolarization of root cortical and stelar cells induced by an acute salt shock—Implications for Na+ and K+ transport into xylem vessels 1 2011
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT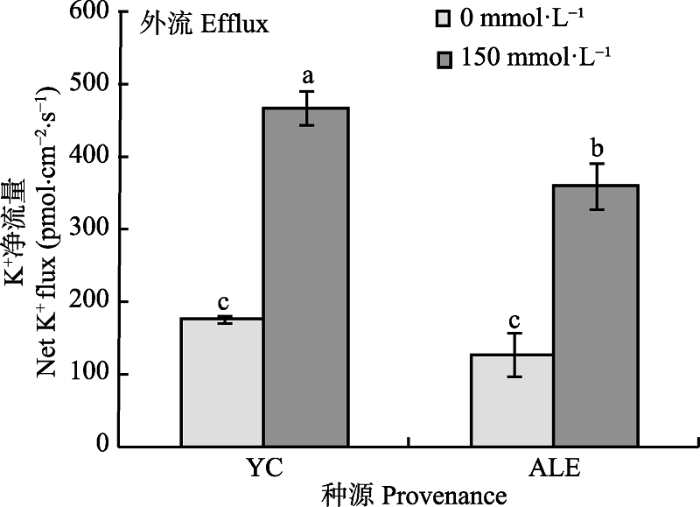 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT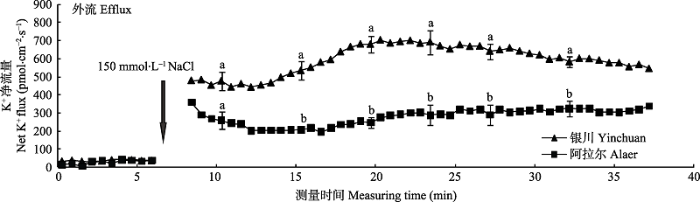 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT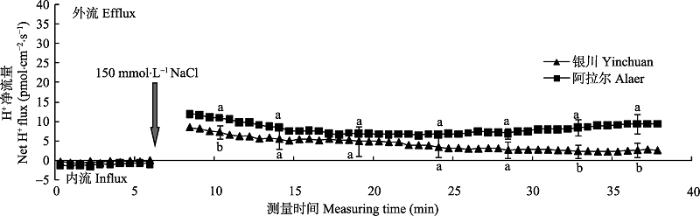 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT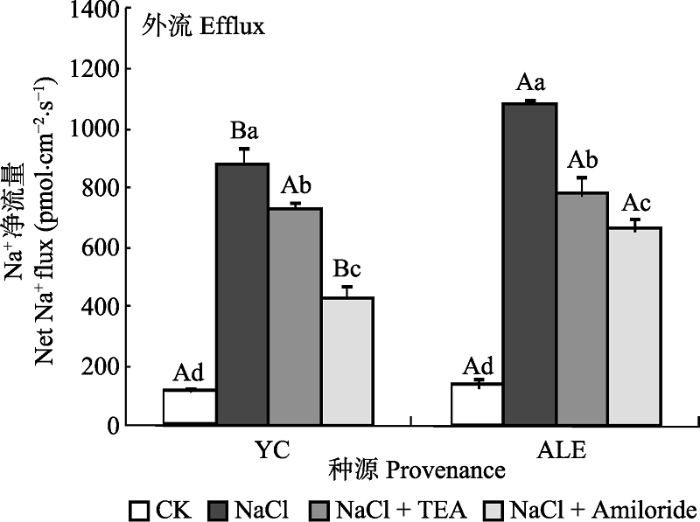 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT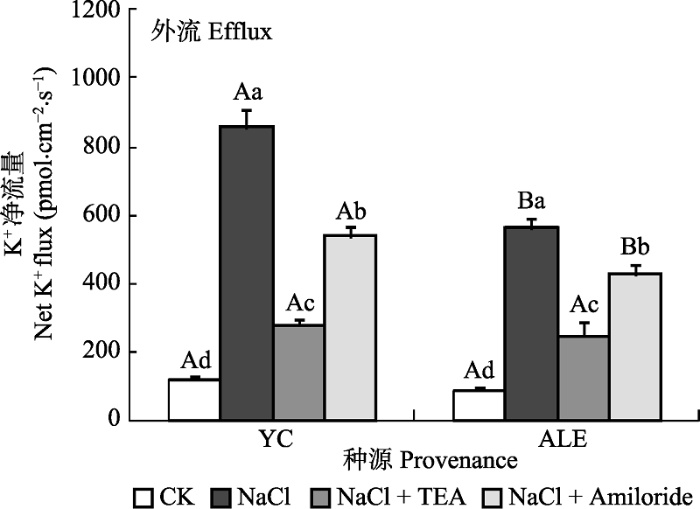 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}