 , 朱盛强1, 刘贞4, 邓蕙菁1, 郑有飞3
, 朱盛强1, 刘贞4, 邓蕙菁1, 郑有飞3
1. 复旦大学环境科学与工程系, 上海 200438;
2. 广东省环境资源利用与保护重点实验室, 广州 510640;
3. 南京信息工程大学江苏省大气环境与装备技术协同创新中心, 南京 210044;
4. 秦皇岛市气象局, 秦皇岛 066000
收稿日期: 2021-03-17; 修回日期: 2021-05-08; 录用日期: 2021-05-08
基金项目: 中国博士后科学基金(No.2020M681157);广东省省级科技计划项目(No.2020B1212060053);江苏省大气环境监测与污染控制高技术研究重点实验室开放基金项目(No.KHK2003);河北省气象局青年基金(No.20ky32)
作者简介: 赵辉(1990—), 男, E-mail: zhaohui_nuist@163.com
通讯作者(责任作者): 郑有飞, E-mail: zhengyf@nuist.edu.cn
摘要:以涡度相关技术为主要观测手段,连续观测冬小麦和水稻生态系统主要生长季净生态系统CO2交换(NEE)的变化规律,评估两种农田生态系统CO2的源/汇功能.结果表明,整个观测期间,两种作物生态系统CO2浓度的日变化曲线呈现白天低、晚上高的"一峰一谷"型,冬小麦生态系统变化较为平缓,而水稻生态系统变化则比较剧烈.冬小麦和水稻生态系统白天30 min CO2通量的平均值分别为-13.4 μmol·m-2·s-1和-12.9 μmol·m-2·s-1,通量最高值分别出现冬小麦的孕穗期与水稻的开花期.此外,两种作物生长季CO2通量表现出"U"形曲线的日变化特点,白天以吸收CO2为主,冬小麦和水稻生态系统分别于12:00和11:30达到吸收峰值;夜间CO2通量变化较为稳定,表现为呼吸排放CO2.两种农田生态系统均表现为碳汇,冬小麦与水稻生态系统净碳交换分别为188.2 g·m-2与233.8 g·m-2.
关键词:涡度相关法CO2通量冬小麦水稻净生态系统碳交换
Evaluation of carbon budget for farmland ecosystem based on the eddy correlation technology
ZHAO Hui1,2,3
, ZHU Shengqiang1, LIU Zhen4, DENG Huijing1, ZHENG Youfei31. Department of Environmental Science and Engineering, Fudan University, Shanghai 200438;
2. Guangdong Key Laboratory of Environmental Protection and Resources Utilization, Guangzhou 510640;
3. Jiangsu Collaborative Innovation Center of Atmospheric Environment and Equipment Technology(CICAEET), Nanjing University of Information Science & Technology, Nanjing 210044;
4. Qinhuangdao Meteorological Bureau, Qinhuangdao 066000
Received 17 March 2021; received in revised from 8 May 2021; accepted 8 May 2021
Abstract: This study used eddy correlation technology to continuously observe the changes of net ecosystem CO2 exchange (NEE) during the main growing seasons of winter wheat and rice ecosystems, and evaluated the CO2 source/sink functions of the two farmland ecosystems. The results showed that: During the whole observation period, the diurnal variation curves of CO2 concentration for the two crops showed typical one-peak-one valley pattern, which was low during the day and high during the night. The diurnal variation curves of CO2 concentration for winter wheat was relatively flat, while that of rice is more severe. The mean values of 30-minute CO2 fluxes during the daytime for winter wheat and rice were -13.4 μmol·m-2·s-1 and -12.9 μmol·m-2·s-1, respectively. The highest fluxes occurred at the booting stage of winter wheat and the flowering stage of rice. In addition, CO2 fluxes of the two crops during the growing season showed a diurnal variation characteristic of a U-shaped curve. Two farmland ecosystems mainly absorbed CO2 during the daytime, and the absorption peaks of winter wheat and rice appeared at 12:00 and 11:30, respectively. The CO2 flux changes during the night were relatively stable, which manifested as the emissions of CO2 respiratory. Two farmland ecosystems were carbon sinks, and the net ecosystem carbon exchange were 188.2 g·m-2 for winter wheat and 233.8 g·m-2 for rice.
Keywords: the eddy covariance methodCO2 fluxwinter wheatricenet ecosystem carbon exchange
1 引言(Introduction)政府间气候变化专门委员会(IPCC)第五次评估报告指出全球气候变暖毋庸置疑.近几十年以来, 由于人类活动向大气中排放过量的二氧化碳(CO2)、甲烷(CH4)与氧化亚氮(N2O)等温室气体, 导致大气过度吸收地表反射的太阳辐射及其他能量, 从而引起大气保温的效应, 最终导致全球气候的逐渐变暖(Freedom et al., 2020).自工业革命以来, 由于经济的增长及社会生产力的飞速发展, 以CO2为主的温室气体含量在大气中持续增加.目前, CO2的过度排放被视为人类活动对温室效应造成影响的主要原因, 相关研究表明(Zickfeld et al., 2009), 全球平均CO2浓度从20世纪50年代的315 ppm已上升至目前的368 ppm, 其中, 由于化石燃料燃烧所排放的CO2量约占温室气体总排放量的57%.
陆地生态系统作为全球四大碳库之一, 是大气圈与生物圈之间CO2交换的桥梁, 它不仅可以利用光合作用不断地吸收与存储大气中的CO2, 以此来发挥碳汇的作用;同时也能够通过植被和土壤的呼吸作用, 向大气中输送CO2, 作为稳定的碳源(Corinne et al., 2009; Fan et al., 2019).鉴于陆地生态系统所具备的这种较为特别的双向性, 使其在全球碳平衡中扮演着极其重要的角色(于贵瑞等, 2006).森林、草地和农田生态系统是陆地生态系统的3个重要组成部分.与前两者下垫面相比, 农田下垫面是受人类活动影响最为剧烈的生态系统, 其生物类型稀少、营养结构单一、生态抵抗力薄弱且稳定性较差(Moureaux et al., 2006), 因此, 农田生态系统相比于其他陆地自然生态系统与外界进行CO2的交换也更为复杂多变, 值得去深入研究.与此同时, 现有的有关碳通量的研究主要侧重于森林与草地生态系统, 而对于农田生态系统开展的研究还较为少见(Yue et al., 2016).因此, 研究农田生态系统碳通量与碳收支状况, 对于更深入地认识陆地生态系统碳循环的规律及预测未来全球变化都具有重要的意义.
目前, 通量箱法与微气象学法是生态尺度CO2通量观测的主要方法.其中, 通量箱法操作简单且成本低廉, 但却改变了箱体内自然气流的湍流状态, 进而影响了排放通量测定的准确性(Mei et al., 2009).测定通量的微气象学方法包括梯度观测法与涡度相关法, 梯度观测法的原理是实时测量植物冠层上方不同高度的CO2浓度、风速、温度和湿度等, 以此来确定通量廓线关系, 然后再推导出近地层湍流通量, 其弊端是在应用过程中需要满足一些特定的假设条件并且达到一定的技术要求.需要指出的是, 涡度相关法是近些年发展起来的一项观测大气与生态系统之间能量与物质交换最直接的方法, 它的计算原理不基于任何假设, 而且不需要特定的经验参数, 具有较为完善的理论基础, 被认为是观测陆地生态系碳通量最为准确可靠的方法(Fares et al., 2009), 但该方法在我国的运用还不是很成熟, 值得去深入研究.
基于此, 本文利用涡度相关系统对长三角地区冬小麦和水稻生长季期间的净生态系统CO2交换(NEE)和相关气象要素进行连续观测, 研究其变化特征, 分析两种作物生长季总初级生产力(GPP)和生态系统呼吸(Reco)的动态变化及累计状况, 以期为准确估算农田生态系统的碳通量提供进一步的数据支持和理论参考.
2 材料与方法(Materials and methods)2.1 观测点与试验材料本研究的观测点位于南京信息工程大学永丰农业气象试验站(32°11′N, 118°40′E), 下垫面作物的种植类型为冬小麦与水稻, 观测时间分别为2017年3月20日—5月25日与2017年7月15日—10月8日.试验种植地开阔平坦, 长为250 m, 宽为150 m.该试验地土壤类型为黄棕壤, 其pH值约为7.0, 土层有机质含量为13.1 g·kg-1, 总氮含量为0.82 g·kg-1, 有效磷和有效钾含量分别为4.2 mg·kg-1和53.3 g·kg-1.两种作物的品种分别为扬麦13与南粳5055, 均为当地主栽品种.其中, 扬麦13是江苏省里下河地区农科所与国家小麦改良中心扬州分中心共同培育的优质高产抗病专用小麦新品种, 系属弱春性, 中早熟类型, 幼苗直立, 分蘖力中等, 成穗率高, 长势健壮;于2016年的11月初采用撒播的方式进行播种, 至下一年的6月初收割.南粳5055是由江苏省农科院粮食作物研究所培育而成的一个优良粳稻新品种, 属早熟晚粳稻品种, 具有长势旺盛、株型紧凑、分蘖力强、叶色较深和群体整齐度好等特点;于2017年的5月中旬播种, 至当年的10月成熟收割.
2.2 观测仪器温度、湿度、太阳辐射等一些常规气象要素利用WatchDog2000系列自动气象站进行观测, 该仪器由美国Spectrum公司生产.仪器的测量间隔可以自定义为1~60 min, 根据本研究的需要, 将仪器设置为30 min获取一个数据.其中, 水汽压差根据Jones等(1992)提出的公式计算得到:
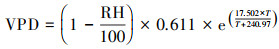 | (1) |
2.3 通量数据的处理通过涡度相关系统观测的原始数据为10 Hz, 采用EddyPro软件进行30 min通量计算与预处理, 该软件由北京天诺基业有限公司提供.其中, 预处理的主要步骤包括:野点剔除、延迟时间校正、二次坐标轴旋转、频率响应修正、水汽密度校正等(朱治林等, 2006).为了保证数据的准确性, 经上述软件输出的30 min通量数据需要进一步实施质量控制, 具体步骤如下所述:
① 首先, 剔除夜间(太阳总辐射 < 10 W·m-2)出现负值的数据;其次, 剔除观测数据中明显不符合正常数据变化范围的异常值(魏甲彬等, 2018);由于降水会干扰农田生态系统中正常的大气流动, 甚至可能引起仪器失灵, 因此, 降雨时期的数据也需要被剔除(朱治林等, 2004);最后, 剔除摩擦风速小于0.15 m·s-1时所对应的通量数据, 以消除湍流发展不充分的影响(邢庆会等, 2014).
② 整个观测期间, 恶劣天气、仪器故障、停电等因素也会导致大量数据缺失, 因此, 有必要对上述缺失的数据进行插补.插补方法主要包括:< 3 h连续缺失数据采用简单的线性内插法(Baldocchi et al., 2003);>3 h且 < 1 d的连续缺失数据采用平均日变化方法进行插补(Falge et al., 2001);此外, >1 d的连续缺失数据不进行插补, 即视为无效数据.
2.4 GPP和Reco的计算方法研究表明(Katul et al., 2001), 生态系统总呼吸Reco对温度的响应呈指数增长的趋势, 目前普遍采用Van′t Hoff经验指数模型间接描述生态系统呼吸对温度的响应特征.考虑到作物夜间不进行光合作用, 夜间生态系统总呼吸Reco(Reco, n)数值上就等同于净生态系统CO2交换(NEE), 所以众多研究通常是建立夜间NEE(NEEn)与气温(T)、5 cm(T5)、10 cm(T10)或20 cm(T20)土层温度的函数关系, 然后将该关系式应用于作物白天生态系统呼吸的估计(Guo et al., 2014).由于数据的缺乏, 本文建立了冬小麦和水稻夜间NEE(NEEn)与气温(T)的关系:
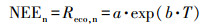 | (2) |
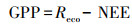 | (3) |
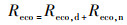 | (4) |
 |
| 图 1 温度与冬小麦和水稻夜间NEE的关系 Fig. 1The relationship between temperature and night NEE for winter wheat and rice |
3 结果与分析(Results and analysis)3.1 环境因子的变化图 2为两种作物生长季期间温度(T)、相对湿度(RH)、光合有效辐射(PAR)和水汽压差(VPD)等气象因子的逐日变化情况.从图中可以看出, 冬小麦生长季期间日均温度、相对湿度、光合有效辐射和水汽压差分别为8.6~24.9 ℃、33.8%~89.9%、29.4~549.0 μmol·m-2·s-1和0.15~1.87 kPa, 平均值分别为17.9 ℃、60.2%、295.6 μmol·m-2·s-1和0.91 kPa.水稻生长季期间日均温度、相对湿度、光合有效辐射和水汽压差的范围分别为14.1~34.7 ℃、49.1%~84.8%、128~786 μmol·m-2·s-1和0.25~2.81 kPa, 平均值分别为26.1 ℃、69.9%、387.9 μmol·m-2·s-1和1.10 kPa.
图 2(Fig. 2)
 |
| 图 2 冬小麦与水稻生长季期间气象因子的逐日变化 Fig. 2Daily changes of meteorological factors during the growing seasons of winter wheat and rice |
由图 3可知, 两种作物生长季期间CO2浓度的日变化均呈现出明显的“一峰一谷”变化特点, 且均为白天低夜间高, 这与国内外****的观测结果类似(Wang et al., 2002; 王长科等, 2003; Newman et al., 2010).冬小麦生长季期间CO2浓度的日变化较为平缓, 而水稻的CO2浓度日变化则比较剧烈, 可能与水稻的生理活动较强有关.从日出开始, 随着大气流动的活跃性与作物的光合能力等汇作用的增强, 大气中的CO2浓度便开始迅速下降, 至16:00左右达到一天中的最低值.这是因为午后边界层不稳定, 湍流交换能力较强, 有利于气体浓度的扩散, 同时午后作物的光合作用较强, 从而吸收了大量的CO2(Bajgain et al., 2018).之后, 由于作物光合作用和对流输送的减弱(即CO2汇强度降低), 土壤及生物呼吸、工业生产等释放的CO2在大气中逐渐累积增加, 浓度呈现上升趋势, 直到第2天上午达到峰值.
图 3(Fig. 3)
 |
| 图 3 冬小麦与水稻生长季期间CO2浓度的日变化 (平均值±标准误差) Fig. 3Diurnal variation of CO2 concentration during the growing seasons of winter wheat and rice |
3.2 CO2通量的变化3.2.1 CO2通量的整体变化图 4为冬小麦与水稻生长季期间白天时段CO2通量30 min平均值的时间序列变化.由于仪器故障、场地断电、天气条件等其他因素的影响, 一些时段数据缺失严重.需要指出的是, 利用涡度相关系统所观测的原始通量数据经EddyPro软件计算出的30 min的CO2通量数据指的是陆地生态系统与大气之间的净碳交换量(NEE), 因此, 本文中所描述的CO2通量均表示的是净生态系统碳交换量(NEE), 即作物对CO2的净吸收量, 负号表示通量方向向下.从图中可以看出, 30 min的CO2通量在整个冬小麦与水稻观测期间的变化范围分别为-0.4~-40.3 μmol·m-2·s-1和-0.1~-38.4 μmol·m-2·s-1, 平均值则分别为-13.4 μmol·m-2·s-1和-12.9 μmol·m-2·s-1.结合图 5可以看出, CO2通量在冬小麦的孕穗期与水稻的开花期达到最高值, 其原因可能是两种作物分别在这两个阶段生命活动力最强, 光合作用最为旺盛, 冠层导度变大, 作物对大气CO2的吸收能力也越强(Demyan et al., 2016).虽然CO2浓度在生长季后期也较高, 然而由于此时作物生命活动力相对较弱, 因此对CO2的吸收减缓.
图 4(Fig. 4)
 |
| 图 4 冬小麦与水稻生长季期间白天CO2通量的时间序列变化 Fig. 4Time series changes of daytime CO2 flux during the growing seasons of winter wheat and rice |
图 5(Fig. 5)
 |
| 图 5 冬小麦与水稻生长季期间白天CO2通量的生育期变化 (平均值±标准误差) Fig. 5Growth period changes of daytime CO2 flux during the growing seasons of winter wheat and rice |
3.2.2 CO2通量的日变化由于所观测的CO2通量反映的是土壤和植被与大气间进行CO2交换的状况, 即植被总初级生产力(GPP)扣除生态系统总呼吸(Reco)之后剩余的部分, 因此, CO2通量的大小表示这种状况的强度.若作物的光合作用强度大于土壤/作物呼吸作用强度, 则作物对CO2表现为净吸收, 且通量为负.反之, 土壤/作物向大气净释放CO2, 且通量为正.
从图 6中可以看出, 冬小麦与水稻生长季期间CO2通量具有相似的日变化特征, 均呈U形曲线变化.夜间的CO2通量为正值且变化较为稳定, 表明两种农田生态系统是大气CO2的源.白天的CO2通量为负值, 说明生态系统以吸收CO2为主.冬小麦和水稻的通量值分别在6:30—7:00和7:00—7:30由正变为负, 即生态系统由碳源转变为碳汇.此后随着太阳辐射强度的增加, 冬小麦与水稻CO2通量迅速增加, 并分别于12:00与11:30达到最大值, 之后缓慢下降, 分别于13:30与12:00时出现了一次拐点, 被称之为作物的“午休”现象(关清等, 2019).这是由于中午气温过高, VPD增大, 叶片快速失水, 从而引起气孔导度减小和光合速率降低导致的, 并且它会伴随着强光而发生或消除(张林海等, 2014).正午过后, 由于光照强度减弱, 植物的光合作用下降, CO2通量随之呈下降状态, 然而这种降低并未引起下午CO2浓度的增高, 可能受午后不稳定的大气层结、较大的湍流强度等气象扩散条件的影响(王尚明等, 2011).
图 6(Fig. 6)
 |
| 图 6 冬小麦与水稻生长季期间CO2通量的日变化 (平均值±标准误差) Fig. 6Diurnal variation of CO2 flux during the growing seasons of winter wheat and rice (mean ± standard error) |
3.3 生态系统碳交换动态变化3.3.1 生长季期间GPP和Reco的日变化图 7为冬小麦和水稻生长季期间GPP与Reco的日变化过程, 从图中可以看出, 两种作物的GPP均呈现出单峰型的变化趋势.由于作物在夜间不进行光合作用, 因此, 夜间的GPP值均为0.从早上6:00—6:30左右, GPP值开始迅速上升, 至中午11:30—12:30达到一天中的最大值后又迅速下降, 最后逐渐趋于0.温度通过影响呼吸酶的活性来影响作物呼吸作用的强度, 如图 7所示, 两种作物的呼吸强度在一定的温度范围内随温度的上升而增强, 所以白天的Reco值略高于夜间, 但这种差异并不是特别明显, 导致Reco在一天中的变化较为平缓.总的来看, 水稻的GPP与Reco值均高于冬小麦, 表明水稻具有更强的生理活动.
图 7(Fig. 7)
 |
| 图 7 冬小麦和水稻的GPP与Reco的日变化 Fig. 7Diurnal changes of GPP and Reco of winter wheat and rice |
3.3.2 生长季期间生态系统碳收支评估如图 8所示, 冬小麦生态系统生长季日NEE、Reco和GPP的变化范围分别为-8.0~2.1、1.6~6.2和1.5~13.6 g·m-2·d-1, 平均值则分别为-2.8、4.0和6.9 g·m-2·d-1.总体而言, NEE、Reco和GPP呈现生长初期与末期较低, 生长中期较高的时间变化特点.生长季初期, 太阳辐射较弱且温度较低, 使得冬小麦进行光合作用与呼吸作用的能力偏弱, 因此, GPP和Reco值较小.之后随着辐射强度的增加及冬小麦的快速生长, NEE、Reco和GPP值均逐渐升高, 表明此阶段冬小麦生态系统碳汇能力的提高.生长季末期, 随着叶片的变黄衰老, 冬小麦的光合和呼吸能力逐渐下降, NEE和GPP值偏低.植物呼吸与土壤呼吸是生态系统中碳排放的主要来源, 它们都受温度的影响.尽管生长季末期冬小麦的呼吸作用较弱, 然而此阶段温度处于最高水平, 导致土壤呼吸对Reco值有较大的贡献, 所以生长季末期的Reco值并没有大幅度降低.
图 8(Fig. 8)
 |
| 图 8 冬小麦和水稻生态系统生长季日NEE、Reco和GPP的动态变化 Fig. 8Daily changes of NEE, Reco and GPP during the growing seasons of winter wheat and rice ecosystems |
此外, 水稻生态系统生长季日NEE、Reco和GPP的变化范围分别为-6.7~3.2、1.5~10.2和1.7~14.4 g·m-2·d-1, 平均值则分别为-2.7、5.3和8.0 g·m-2·d-1.总体而言, Reco和GPP在水稻整个生长季呈下降的趋势.水稻生长季温度与光合有效辐射从前期到后期逐渐下降, 使得Reco和GPP呈现与之相同的变化状态.由于水稻生长前期Reco和GPP均较高, 因此, 生长季前期的NEE值在整个生育期并不是最高, NEE的最高值出现在生长前期与中期之间, 可能与该阶段完整的水稻冠层结构有关.同时, 9月前后的几天表现为碳排放与降水等天气有关.
从图 9中可以看出, 两种作物通过光合作用固定的CO2远高于碳排放总量, 即冬小麦和水稻生态系统均表现为碳汇.冬小麦生态系统的CO2净吸收量为188.2 g·m-2, 累计吸收CO2为459.1 g·m-2, 累计排放CO2为270.9 g·m-2, Reco约占GPP的59.0%.此外, 水稻生态系统的CO2净吸收量为233.8 g·m-2, 累计吸收CO2为689.3 g·m-2, 累计排放CO2为455.5 g·m-2, Reco约占GPP的66.1%.
图 9(Fig. 9)
 |
| 图 9 冬小麦和水稻生态系统生长季NEE、Reco和GPP的累积状况 Fig. 9Cumulative status of NEE, Reco and GPP during the growing seasons of winter wheat and rice ecosystems |
4 结论(Conclusions)1) 冬小麦和水稻生长季CO2浓度的日变化表现为“一峰一谷”的变化趋势, 且白天浓度低、晚上浓度高.此外, 冬小麦生长季CO2浓度的日变化曲线较为平缓, 而水稻生长季CO2浓度的日变化则比较剧烈.整个观测期间, 冬小麦和水稻生态系统的30 min CO2通量的变化范围分别为-0.4~-40.3 μmol·m-2·s-1和-0.1~-38.4 μmol·m-2·s-1, 平均值则分别为-13.4 μmol·m-2·s-1和-12.9 μmol·m-2·s-1.通量最高值分别出现冬小麦的孕穗期与水稻的开花期, 该阶段作物光合能力强、生命活动力旺盛, 因此对CO2的净吸收量较高.
2) 两种作物生长季CO2通量的日变化均呈“U”形曲线, 白天以吸收CO2为主, 冬小麦和水稻分别于12:00和11:30达到吸收峰值.夜间CO2通量变化较为稳定, 表现为呼吸排放CO2.此外, 观测期间两种作物均发现了光合“午休”, 这是由于中午气温过高, VPD增大, 叶片快速失水, 而引起气孔导度减小和光合速率降低的现象.
3) 为了量化作物通过呼吸作用吸收的CO2, 以及通过自身光合作用排放的CO2, 基于Van′t Hoff经验指数模型, 本文建立了夜间两种作物NEE与气温的函数关系, 以此来区分总初级生产力GPP与生态系统总呼吸Reco.研究发现, 整个观测期间冬小麦生态系统净碳交换为188.2 g·m-2, 总初级生产力为459.1 g·m-2, 碳排放为270.9 g·m-2;水稻生态系统净碳交换为233.8 g·m-2, 总初级生产力为689.3 g·m-2, 碳排放为455.5 g ·m-2.
参考文献
| Bajgain R, Xiao X, Basara J, et al. 2018. Carbon dioxide and water vapor fluxes in winter wheat and tallgrass prairie in central Oklahoma[J]. Science of the Total Environment, 644: 1511-1524. DOI:10.1016/j.scitotenv.2018.07.010 |
| Baldocchi D D. 2003. Assessing ecosystem carbon balance: problems and prospects of the eddy covariance technique[J]. Global Change Biology, 9: 478-492. |
| Corinne L Q, Raupach M R, Canadell J G, et al. 2009. Trends in the sources and sinks of carbon dioxide[J]. Nature Geoscience, 2: 831-836. DOI:10.1038/ngeo689 |
| Demyan M S, Ingwersen J, Funkuin Y N, et al. 2016. Partitioning of ecosystem respiration in winter wheat and silage maize-modeling seasonal temperature effects[J]. Agriculture, Ecosystems & Environment, 224: 131-144. |
| Falge E, Baldocchi D, Olson R, et al. 2001. Gap filling strategies for defensible annual sums of net ecosystem exchange[J]. Agricultural and Forest Meteorology, 107: 43-69. DOI:10.1016/S0168-1923(00)00225-2 |
| Fan J, McConkey B G, Liang B C, et al. 2019. Increasing crop yields and root input make Canadian farmland a large carbon sink[J]. Geoderma, 336: 49-58. DOI:10.1016/j.geoderma.2018.08.004 |
| Fares S, McKay M, Holzinger R, et al. 2010. Ozone fluxes in a Pinus ponderosa ecosystem are dominated by non-stomatal processes: Evidence from long-term continuous measurements[J]. Agricultural and Forest Meteorology, 150: 420-431. DOI:10.1016/j.agrformet.2010.01.007 |
| Freedom V B, Trucchi E, Goropashnaya A V, et al. 2020. Impact of past climate warming on genomic diversity and demographic history of collared lemmings across the Eurasian Arctic[J]. Proceedings of the National Academy of Sciences of the United States of America, 117: 3026-3033. DOI:10.1073/pnas.1913596117 |
| 关清, 郑有飞. 2019. 冬小麦田CO2通量观测及其与气象因子的关系[J]. 科学技术与工程, 19: 313-320. |
| Guo W H, Kang S Z, Li F S, et al. 2014. Variation of NEE and its affecting factors in a vineyard of arid region of northwest China[J]. Atmospheric Environment, 84: 349-354. DOI:10.1016/j.atmosenv.2013.11.057 |
| Katul G, Lai C T, Sch?fer K, et al. 2001. Multiscale analysis of vegetation surface fluxes: from seconds to years[J]. Advances in Water Resources, 24: 1119-1132. DOI:10.1016/S0309-1708(01)00029-X |
| Mei B, Zheng X, Xie B, et al. 2009. Nitric oxide emissions from conventional vegetable fields in southeastern China[J]. Atmospheric Environment, 43: 2762-2769. DOI:10.1016/j.atmosenv.2009.02.040 |
| Moureaux C, Debacq A, Bodson B, et al. 2006. Annual net ecosystem carbon exchange by a sugar beet crop[J]. Agricultural and Forest Meteorology, 139: 25-39. DOI:10.1016/j.agrformet.2006.05.009 |
| Newman S, Jeong S, Fischer M L, et al. 2013. Diurnal tracking of anthropogenic CO2 emissions in the Los Angeles basin megacity during spring 2010[J]. Atmospheric Chemistry and Physics, 13: 4359-4372. DOI:10.5194/acp-13-4359-2013 |
| 王长科, 王跃思, 刘广仁, 等. 2003. 北京城市大气CO2浓度变化特征及影响因素[J]. 环境科学, 24(4): 13-17. |
| 王尚明, 胡继超, 吴高学, 等. 2011. 亚热带稻田生态系统CO2通量特征分析[J]. 环境科学学报, 31(1): 217-224. |
| Wang Y, Wang C, Guo X, et al. 2002. Trend, seasonal and diurnal variations of atmospheric CO2 in Beijing[J]. Chinese Science Bulletin, 47: 2050-2055. DOI:10.1360/02tb9444 |
| 魏甲彬, 徐华勤, 周玲红, 等. 2018. "双季稻-冬闲田"生态系统碳交换动态变化及其影响因素[J]. 农业环境科学学报, 37: 1035-1044. |
| 邢庆会, 韩广轩, 于君宝, 等. 2014. 黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素[J]. 生态学报, 34(17): 4966-4979. |
| 于贵瑞, 孙晓敏. 2006. 陆地生态系统通量观测的原理与方法[M]. 北京: 高等教育出版社. |
| Yue Y, Ni J, Ciais P, et al. 2016. Lateral transport of soil carbon and land-atmosphere CO2 flux induced by water erosion in China[J]. Proceedings of the National Academy of Sciences of the United States of America, 113: 6617-6622. DOI:10.1073/pnas.1523358113 |
| 张林海, 仝川, 曾从盛. 2014. 河口湿地近地面大气CO2浓度日变化和季节变化[J]. 环境科学, 35(3): 879-884. |
| 朱治林, 孙晓敏, 温学发, 等. 2006. 中国通量网(ChinaFLUX)夜间CO2涡度相关通量数据处理方法研究[J]. 中国科学(D辑: 地球科学), S1: 34-44. |
| 朱治林, 孙晓敏, 袁国富, 等. 2004. 非平坦下垫面涡度相关通量的校正方法及其在ChinaFLUX中的应用[J]. 中国科学(D辑: 地球科学), S2: 37-46. |
| Zickfeld K, Eby M, Matthews H D, et al. 2009. Setting cumulative emissions targets to reduce the risk of dangerous climate change[J]. Proceedings of the National Academy of Sciences of the United States of America, 106: 16129-16134. DOI:10.1073/pnas.0805800106 |
