Circadian rhythm of root pressure in popular and its driving factors
GUOJian-Rong, WANXian-Chong*, Institute of New Forestry Technology, Chinese Academy of Forestry, Beijing 100091, China 通讯作者:* 通信作者Author for correspondence (E-mail:wxc@caf.ac.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31270648)
关键词:根压;离体根系;日周期;影响因素 Abstract Aims Our main purposes were to investigate root pressure and its circadian rhythm of excised roots in ‘84K’ popular (Populus alba × P. glandulosa) cultured in soil and solution, to explore the influencing factors and their relationships with root pressure systematically and to understand the generation and rhythm regulation of root pressure. Methods We investigated the root pressure of excised roots in ‘84K’ popular using the method of digital pressure transducer. The diurnal rhythm of excised roots was conducted through different experimental treatments including sampling in different time, defoliation and girdling, together with ambient condition like soil temperature, differential or consistant temperature during day and night. Then we discussed the effects of root respiration and hydraulic conductivity on root pressure by further using chemical inhibitor. Furthermore, diurnal variation of osmotic potential and ions content as well as soluble sugar content of exudation was determined in order to explore their relationships with root pressure rhythm. Important findings Root pressure of excised roots in popular had diurnal rhythm which was higher during daytime and lower overnight. It reached its peak value in the morning to noon and valley value at 20:00. Root pressure of excised roots sampled at different time and cultured in different medium had influence on the rhythm of root pressure to some degrees, but did not the general rhythm of high in daytime and low overnight. Defoliation, girdling and the inhibitors for root respiration or cytomembrane hydraulic conductivity could affect the maximum value of root pressure while have no significant influence on the daily rhythm. Defoliation, girdling and respiration inhibitor reduced the maximum value of root pressure, whereas the hydraulic conductivity inhibitor had little influence on root pressure. The maximum value of root pressure declined with the decrease in soil temperature which could change the rhythm of root pressure. The synchronous change in the maximum value of root pressure and root respiration rate with temperature indicated that root respiration contributed to the change of root pressure along with temperature. Osmotic potential of root exudation was higher during the daytime and lower at night. Diurnal variations of ions and soluble sugar content of exudation were consistant with that of osmotic potential. The peak of root pressure measured under the condition of differential temperature during day and night was significant higher than that measured under constant temperature. In conclusion, root pressure of the poplar ‘84K’ showed significant diurnal rhythm, i.e. higher during the daytime and lower at night. The maximum value of root pressure was mainly regulated by root respiration metabolism. The factors such as respiration inhibitor, respiration substrate and temperature influence the value of the maximum root pressure of poplar ‘84K’. Root hydraulic conductivity had no significant influence on root pressure.
显示原图|下载原图ZIP|生成PPT 图3不同温度条件下测定的根压最大值和呼吸速率(平均值±标准偏差, n = 6)。 -->Fig. 3The maximum of root pressure and respiration rate measured under different soil temperatures (mean ± SD, n = 6). -->
2.3 昼夜温差对根压的影响
昼夜恒温和昼夜温差条件下, 根压均表现出昼高夜低的日周期, 且二者的根压峰值和根压值谷值出现时间基本一致。二者相比, 昼夜温差情况下, 根压峰值显著的高于昼夜恒温条件下, 根压及其节律维持时间更长(图4)。 显示原图|下载原图ZIP|生成PPT 图4昼夜恒温(25 ℃/25 ℃)和昼夜温差(25 ℃/19 ℃)条件下的根压。 -->Fig. 4The root pressure under constant temperature during day and night (25 °C/25 °C) and different temperature during day and night (25 °C /19 °C). -->
2.4 环割和摘叶对根压的影响
提前24 h进行环割、摘叶处理, 之后和对照组苗木一起进行根压测定。处理组和对照组根压的昼夜节律表现一致, 但环割或摘叶处理大大地降低了根压的峰值(图5)。表现出昼高夜低的日节律, 从20:00开始逐渐上升, 到5:00后, 开始快速上升, 直至9:00-11:00, 之后开始迅速下降。对照组根压的峰值随测定时间的延长迅速降低, 处理组根压峰值随时间变化较小。 显示原图|下载原图ZIP|生成PPT 图5摘叶和环割对根压的影响。 -->Fig. 5Effects of defoliation and girdling on root pressure. -->
2.5 呼吸速率抑制剂和根系导水率抑制剂对根压的影响
为了便于化学药品的处理, 该试验用了水培的苗木, 从图6可见其根压和土生苗木相似也呈现昼高夜低的节律性。但由于抑制剂的存在, 大大降低了根压的峰值。其中呼吸抑制剂NaN3单独施用或和HgCl2一起施用都严重降低了根压峰值。而细胞膜水分导度抑制剂HgCl2也一定程度上降低了根压峰值。 显示原图|下载原图ZIP|生成PPT 图6抑制剂处理对根压的影响。 -->Fig. 6Effects of inhibitors on root pressure. -->
CaoKF, YangSJ, ZhangYJ, BrodribbTJ (2012). The maximum height of grasses is determined by roots. Ecology Letters, 15, 666-672. [本文引用: 2]
[2]
ClarksonDT, CarvajalM, HenzlerT, WaterhouseRN, SmythAJ, CookeDT, SteudleE (2000). Root hydraulic condu¬ctance: Diurnal aquaporin expression and the effects of nutrient stress. Journal of Experimental Botany, 51, 61-70. [本文引用: 1]
[3]
CochardH, EwersF, TyreeM (1994). Water relations of a tropical vine-like bamboo (Rhipidocladum racemiflorum): Root pressures, vulnerability to cavitation and seasonal changes in embolism. Journal of Experimental Botany, 45, 1085-1089. [本文引用: 1]
[4]
EnnsL, CannyM, McCullyM (2000). An investigation of the role of solutes in the xylem sap and in the xylem paren¬chyma as the source of root pressure. Protoplasma, 211, 183-197. [本文引用: 3]
[5]
EwersFW, AméglioT, CochardH, BeaujardF, MartignacM, VandameM, BodetC, CruiziatP (2001). Seasonal variation in xylem pressure of walnut trees: Root and stem pressures. Tree Physiology, 21, 1123-1132. [本文引用: 3]
[6]
EwersFW, CochardH, TyreeMT (1997). A survey of root pressures in vines of a tropical lowland forest. Oecologia, 110, 191-196. [本文引用: 1]
[7]
FanJF, LiLing, HanYF, LiJR (2002). Establishment of leaf-explant regeneration system of popular 84K. Journal of Northwest Forestry University, 17(2), 33-36. (in Chinese with English abstract)[樊军锋, 李玲, 韩一凡, 李嘉瑞 (2002). 84K杨叶片外植体再生系统的建立 . 西北林学院学报, 17(2), 33-36.]
[8]
FisherJB, GuillermoA, EwersFW, López-PortilloJ (1997). Survey of root pressure in tropical vines and woody spe-cies. International Journal of Plant Sciences, 158, 44-50. [本文引用: 1]
[9]
HenzlerT, WaterhouseRN, SmythAJ, CarvajalM, CookeDT, SchäffnerAR, SteudleE, ClarksonDT (1999). Diurnal variations in hydraulic conductivity and root pressure can be correlated with the expression of putative aquaporins in the roots of Lotus japonicus. Planta, 210, 50-60. [本文引用: 2]
[10]
HolbrookNM, AhrensET, BurnsMJ, ZwienieckiMA (2001). In vivo observation of cavitation and embolism repair using magnetic resonance imaging. Plant Physiology, 126, 27-31. [本文引用: 4]
[11]
KamaluddinM, ZwiazekJJ (2001). Metabolic inhibition of root water flow in red-osier dogwood (Cornus stolonifera) seedlings. Journal of Experimental Botany, 52, 739-745. [本文引用: 1]
[12]
KramerPJ, BoyerJS (1995).Water Relations of Plants and Soils. Academic Press, Kaedinya, Australia. [本文引用: 4]
[13]
KusakinaJ, GouldPD, HallA (2014). A fast circadian clock at high temperatures is a conserved feature across Arabidopsis accessions and likely to be important for vegetative yield. Plant, Cell & Environment, 37, 327-340.
[14]
LiKY, FanJF, ZhaoZ, ZhouYX, GaoJS (2007). Prioritization of the system of regeneration and genetictransformation of Populus 84K. Journal of Northwest A & F University (Natural Science Edition), 35(7), 90-96. (in Chinese with English abstract)[李科友, 樊军锋, 赵忠, 周永学, 高建社(2007). 84K杨再生和遗传转化体系的优化 . 西北农林科技大学学报(自然科学版), 35(7), 90-96.] [本文引用: 1]
[15]
SecchiF, ZwienieckiMA (2012). Analysis of xylem sap from functional (nonembolized) and nonfunctional (embolized) vessels of Populus nigra: Chemistry of refilling. Plant Physiology, 160, 955-964.
[16]
SperryJS, HolbrookNM, ZimmermannMH, TyreeMT (1987). Spring filling of xylem vessels in wild grapevine. Plant Physiology, 83, 414-417. [本文引用: 2]
[17]
SteudleE, MurrmannM, PetersonCA (1993). Transport of water and solutes across maize roots modified by punctur¬ing the endodermis (further evidence for the composite tran¬¬¬sport model of the root). Plant Physiology, 103, 335-349.
[18]
TyreeMT, SperryJS (1989). Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Biology, 40, 19-38. [本文引用: 1]
[19]
TyreeMT, YangS, CruiziatP, SinclairB (1994). Novel methods of measuring hydraulic conductivity of tree root systems and interpretation using AMAIZED (A maize-root dyn¬¬amic model for water and solute transport). Plant Physiology, 104, 189-199. [本文引用: 1]
[20]
TyreeMT, ZimmermannMH (2002). Xylem Structure and the Ascent of Sap. Springer, Berlin , Germany. [本文引用: 1]
[21]
WanX, ZwiazekJJ (1999). Mercuric chloride effects on root water transport in aspen seedlings. Plant Physiology, 121, 939-946. [本文引用: 1]
[22]
WangF, TianX, DingY, WanX, TyreeMT (2011). A survey of root pressure in 53 Asian species of bamboo. Annals of Forest Science, 68, 783-791. [本文引用: 1]
[23]
WangSY, ChenQJ, WangWL, WangXC, LuMZ (2005). Cultivation of Populus 84K transferred salt-tolerant genes OsNHX1. Chinese Science Bulletin, 50(2), 140-144. (in Chinese)[王树耀, 陈其军, 王文龙, 王学臣, 卢孟柱 (2005). 转OsNHX1基因耐盐84K杨的培育 . 科学通报, 50(2), 140-144.] [本文引用: 4]
[24]
WegnerLH (2014). Root pressure and beyond: Energetically uphill water transport into xylem vessels? Journal of Experimental Botany, 65, 381-393. [本文引用: 1]
[25]
WilsonC, KramerP (1949). Relation between root respiration and absorption. Plant Physiology, 24, 55-59.
[26]
WismerWV, MarangoniAG, YadaRY (1995). Low¬tem¬perature sweetening in roots and tubers. Horticultural Reviews, 17, 203-231. [本文引用: 1]
[27]
YangSJ, ZhangYJ, SunM, GoldsteinG, CaoKF (2012). Rec¬overy of diurnal depression of leaf hydraulic conductance in a subtropical woody bamboo species: Embolism refilling by nocturnal root pressure. Tree Physiology, 32, 414-422. [本文引用: 1]
The maximum height of grasses is determined by roots. 2 2012
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
Root hydraulic condu¬ctance: Diurnal aquaporin expression and the effects of nutrient stress. 1 2000
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
Water relations of a tropical vine-like bamboo (Rhipidocladum racemiflorum): Root pressures, vulnerability to cavitation and seasonal changes in embolism. 1 1994
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
An investigation of the role of solutes in the xylem sap and in the xylem paren¬chyma as the source of root pressure. 3 2000
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... ), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
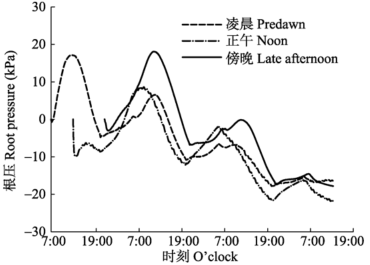
... 本研究中, 无论土壤盆栽还是溶液培养的杨树苗木, 其根压都呈现出明显的昼夜节律, 昼高夜低.一天当中不同的采样时间对根压周期性有一定的影响, 但总体昼高夜低没有改变.到达谷值的时间差别不大, 但与凌晨和傍晚采样相比, 正午采样测得的根压峰值显著前移, 而且峰值最小, 说明测定时间对根压的节律及峰值大小均有影响.正午时间取样测定根压, 此时的植株处在水分亏缺状态下, 根系吸收的水分一部分进入到水势较低的根系薄壁细胞中, 使导管中水分减少, 根压值减小.由此可知, 根压峰值的大小及出现时间与根系水分状况和渗透活动有关, 这与其他****的发现 (Enns et al., 2000; Yang et al., 2012) 一致. ...
Seasonal variation in xylem pressure of walnut trees: Root and stem pressures. 3 2001
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
A survey of root pressures in vines of a tropical lowland forest. 1 1997
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
84K杨叶片外植体再生系统的建立 2002
Survey of root pressure in tropical vines and woody spe-cies. 1 1997
... ‘84K’ (Populus alba × P. glandulosa)由杂交获得, 随着该树种再生和遗传转化体系的建立(樊军锋等, 2002; 李科友等, 2007), 基于这个树种开展了许多基因转化及抗性机理研究(王树耀等, 2005).我们利用‘84K杨’这一模式植物, 通过对其离体根系的根压测定研究, 探索根压昼夜节律及其对取样时间、温度等外界环境的响应; 与呼吸代谢、根系导水率、光合产物向下运输及伤流液中离子及可溶性糖含量的关系, 以便为进一步研究木本植物根压节律及其调控机制提供科学依据. ...
Diurnal variations in hydraulic conductivity and root pressure can be correlated with the expression of putative aquaporins in the roots of Lotus japonicus. 2 1999
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
In vivo observation of cavitation and embolism repair using magnetic resonance imaging. 4 2001
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... ), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... 的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
Metabolic inhibition of root water flow in red-osier dogwood (Cornus stolonifera) seedlings. 1 2001
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... ).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
A fast circadian clock at high temperatures is a conserved feature across Arabidopsis accessions and likely to be important for vegetative yield. Plant, 2014
Transport of water and solutes across maize roots modified by punctur¬ing the endodermis (further evidence for the composite tran¬¬¬sport model of the root). 1993
Vulnerability of xylem to cavitation and embolism. 1 1989
Novel methods of measuring hydraulic conductivity of tree root systems and interpretation using AMAIZED (A maize-root dyn¬¬amic model for water and solute transport). 1 1994
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
Xylem Structure and the Ascent of Sap. Springer, Berlin 1 2002
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
Root pressure and beyond: Energetically uphill water transport into xylem vessels? 1 2014
... 研究中我们借助呼吸代谢抑制剂NaN3和细胞水分导度抑制剂HgCl2, 探索根系呼吸速率和水力学导度对离体根系根压的影响.NaN3能降低根系的呼吸速率和水力学导度(Kamaluddin & Zwiazek, 2001), HgCl2通过对细胞膜水孔堵塞来抑制根系水导率(Wan & Zwiazek, 1999).目前, 常用于根压测定的方法有3种, 最简单的方法是气泡压力计法(Ewers et al., 1997; Fisher et al., 1997; Wang et al., 2011), 许多****用这个方法调查研究植物凌晨的根压, 这个方法的局限性是只能测定特定时间的根压.另一种是根压力探针法, 这个方法不仅能测定实时根压, 还能对根压进行连续测定(Steudle et al., 1993; Henzler et al., 1999).而压力转换器在根压测定中的应用给根压的连续测定带来了极大方便(Cochard et al., 1994; Ewers et al., 2001; Cao et al., 2012), 使得我们可以在各种条件下进行根压的连续监测. ...
Relation between root respiration and absorption. 1949
Low¬tem¬perature sweetening in roots and tubers. 1 1995
... ‘84K’ (Populus alba × P. glandulosa)由杂交获得, 随着该树种再生和遗传转化体系的建立(樊军锋等, 2002; 李科友等, 2007), 基于这个树种开展了许多基因转化及抗性机理研究(王树耀等, 2005).我们利用‘84K杨’这一模式植物, 通过对其离体根系的根压测定研究, 探索根压昼夜节律及其对取样时间、温度等外界环境的响应; 与呼吸代谢、根系导水率、光合产物向下运输及伤流液中离子及可溶性糖含量的关系, 以便为进一步研究木本植物根压节律及其调控机制提供科学依据. ...
Rec¬overy of diurnal depression of leaf hydraulic conductance in a subtropical woody bamboo species: Embolism refilling by nocturnal root pressure. 1 2012
... 根压产生于木质部溶质渗透势(Henzler et al., 1999; Enns et al., 2000), 与根系的生理活动及内皮层内外的水势差有关, 根压取决于对矿质元素的吸收和向木质部的运输(Tyree et al., 1994; Kramer & Boyer, 1995), 这一主动过程需要腺苷三磷酸(ATP) (Enns et al., 2000; Ewers et al., 2001).中柱薄壁细胞和木质部管道中渗透势的升高和降低是一个连续的过程, 因此对ATP的需求也是持续的.水分通过木质部薄壁组织中细胞膜是由水和溶质的联合转运来驱动的, 而溶质需要代谢能量来补充, 以持续维持水分分泌所需的溶质梯度(Wegner, 2014).呼吸代谢是植物ATP的来源, 因此呼吸作用在根系水分吸收中起着关键作用(Wilson & Kramer, 1949).另一类渗透物质是可溶性的有机物质(Kramer & Boyer, 1995), 它们除直接调节渗透势之外, 还通过影响呼吸代谢来影响矿质元素的吸收和积累.另外, 离体根系的水力学导度存在昼夜周期性(Henzler et al., 1999; Clarkson et al., 2000), 研究发现Lotus japonicus的水力学导度和根压的昼夜节律相吻合(Henzler et al., 1999). ...

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT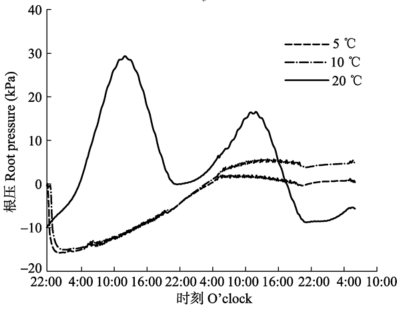 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT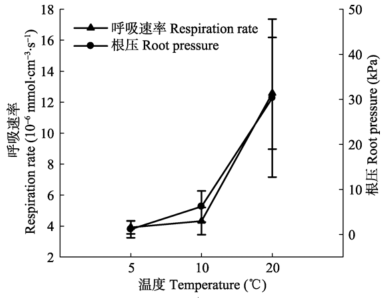 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT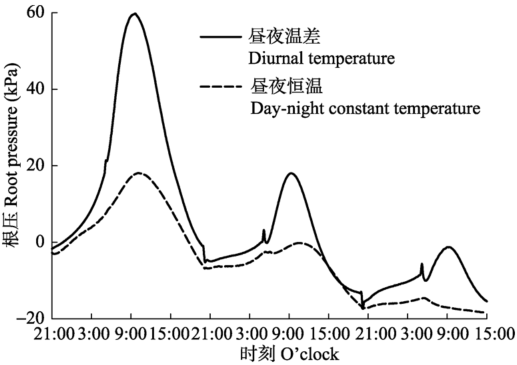 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT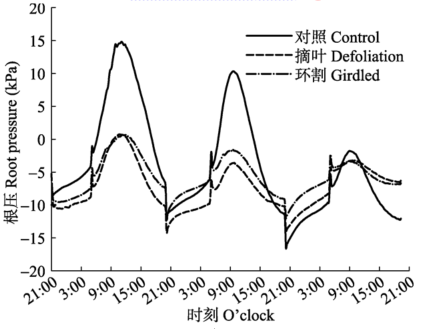 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT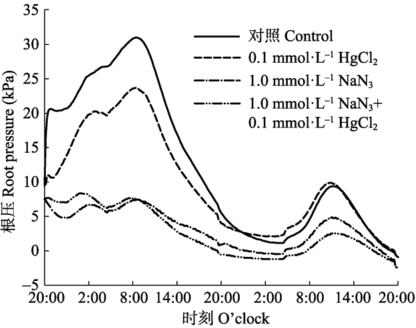 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT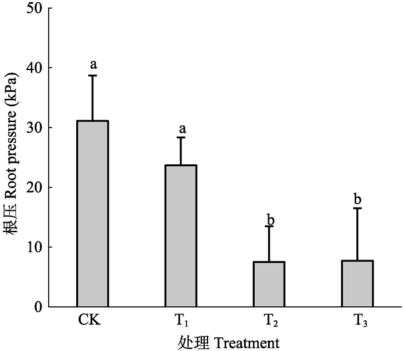 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT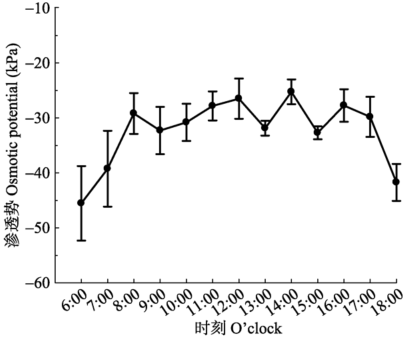 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT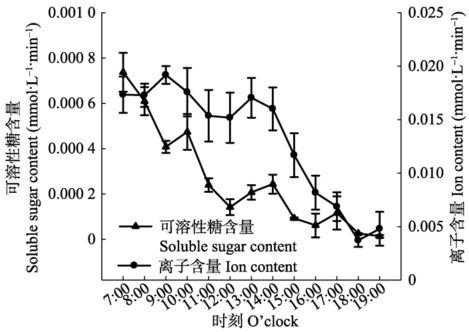 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}