0 引言
【研究意义】大豆(Glycine max)是世界上最重要的作物之一,在我国有悠久的种植历史,是我国重要的油料作物。全球大豆种植面积1.02亿公顷,年产量2.54亿吨[1]。影响大豆生产的病害很多,而大豆孢囊线虫病是制约我国大豆生产的主要病害之一。大豆孢囊线虫病在我国东北、华北及黄淮海等大豆主产区广泛分布,一般造成大豆减产5%—10%,严重地块减产30%—50%,甚至绝收[2],其致病的病原线虫主要为大豆孢囊线虫(Heterodera glycines)。2015年7月至2016年7月,作者从江西婺源大豆根际土样中及根系上发现一个孢囊线虫群体,其形态特征和大豆孢囊线虫存在较大差异,明确该孢囊线虫群体的种类及其对豆科植物寄生性,对病害防控具有重要意义。【前人研究进展】孢囊线虫系一类定居性的植物内寄生线虫,很多具经济重要性的种类都属于孢囊线虫属(Heterodera)。目前国内外报道的孢囊线虫属描述的种类有80多个,其中在全球范围内给农业生产造成严重损失的有大豆孢囊线虫、甜菜孢囊线虫(H. schachtii)、禾谷孢囊线虫(H. avenae)等[3,4,5]。在我国,引起农作物病害的重要孢囊线虫种类主要有大豆孢囊线虫、禾谷孢囊线虫和菲利普孢囊线虫(H. filipjevi)[6,7,8]等。近年来国内报道的孢囊属新种有中华孢囊线虫(H. sinensis)[9]、冯氏孢囊线虫(H. fengi)[10]、海南孢囊线虫(H. hainanensis)[11]和广东孢囊线虫[12](H. guangdongensis)等;描述的新记录种有旱稻孢囊线虫(H. elachista)[13]、朝鲜孢囊线虫(H. koreana)[14,15]等。2016年,KANG等[16]描述了一种寄生于大豆的孢囊线虫新种——野生豆孢囊线虫(H. sojae),该孢囊线虫目前仅在韩国被报道,对于该孢囊的致病性和寄主范围也还未见相关研究报道。【本研究切入点】2015—2016年,在对江西省主要农作物孢囊线虫病害调查中,于上饶市婺源县大豆根际土样及根系上发现一个孢囊线虫群体,该孢囊线虫的形态特征与大豆孢囊线虫有较大差异,有必要明确其种类、寄主范围及致病性等。【拟解决的关键问题】结合形态学和分子生物学方法对采集自江西婺源大豆上的孢囊线虫群体进行鉴定,明确其寄主范围及致病性特点,为今后进一步的研究和防控打下基础。1 材料与方法
1.1 孢囊线虫的采集、分离与扩繁
大豆根系及根际土壤于2015年7月至2016年7月采自江西省上饶市婺源县万岭村。将带回的大豆根系洗净后在解剖镜下观察,直接解剖法获取根系上的孢囊,土样采用淘洗过筛法分离土样中的孢囊[5]。采用浅盘法分离土壤中的2龄幼虫(J2)和雄虫。在盛满灭菌沙土(土壤:沙=3:1)的塑料钵(直径12 cm、高15 cm)中种植和样品采集地所种植的相同大豆品种(婺源罗汉豆),每盆定苗一株,待大豆长至一片真叶时参照KONG等的方法[17],配制和接种孢囊卵悬浮液,用1 mL的枪头在大豆苗的周围均匀插3个小孔,将卵悬浮液分别从小孔注入后,覆好灭菌土。接种后5 d内不大量浇水,置于25—28℃温室条件下培养。
1.2 形态学观察及测量
选择收集饱满的孢囊,参照冯志新[18]的方法制作阴门锥切片。参照谢辉[19]的方法挑取J2和雄虫制作玻片。65℃水浴加热杀死线虫,用FAA(福尔马林-冰醋酸-乙醇固定液)固定,制作成临时玻片,在Leica光学显微镜(DM2500)下观察、拍照、测量。形态学观察特征及测量值包括J2的体长、最大体宽、口针长、食道长、背食道腺开口到口针基球的距离、尾长及透明尾长;孢囊的形态、孢囊下桥的有无、膜孔及类型、泡状囊泡的有无及阴门裂长度等[5]。1.3 分子生物学鉴定
1.3.1 DNA提取 参考亓晓莉等[20]的方法,挑取田间采集和温室单孢囊扩繁群体中大豆根际土样及根系上的单孢囊,置于含有10 μL双蒸水、3 μL的蛋白酶K(600 mg·mL-1,Roche,德国)和7 μL PCR-buffer缓冲液(含mg2+)(TaKaRa,大连)的离心管中,在液氮中速冻1 min后用灭菌玻璃棒研磨,待冰完全融化后,重复在液氮中速度研磨3次后,置于-80℃条件下至少保存2 h,将离心管转入PCR仪中,进行DNA提取,反应过程如下:65℃下温育90 min,85℃条件下保存10 min,最后4℃保存。DNA提取的产物在12 000 r/min离心1 min,取上清DNA悬浮液于-20℃中保存备用。1.3.2 序列扩增 以上述单孢囊DNA为模板,参考CUI等[21]的方法,以引物D2A(5′-ACA AGT ACC GTG AGG GAA AGT TG-3′)和D3B(5′-TCG GAA GGA ACC AGC TAC TA-3′)扩增核糖体DNA 28S D2-D3区域,以引物AB28(5′-ATA TGC TTA AGT TCA GCG GT-3′)和TW81(5′-GTT TCC GTA GGT GAA CCT GC-3′)[22]PCR扩增内转录间隔区(internal transcribed spacer,ITS),PCR扩增体系:模板DNA 2 μL,2×Taq PCR StarMix buffer(TaKaRa,大连)12.5 µL,10 μmol·L-1的引物各0.5 µL,无菌双蒸水补充至25 μL。扩增条件为94℃预变性,5 min,94℃变性30 s,58℃退火45 s,72℃延伸1.5 min,共35个循环,72℃终延伸10 min,4℃保存。扩增产物用1.5%的琼脂糖凝胶分离后,观察照相。扩增产物采用琼脂糖凝胶纯化试剂盒(北京天根生物科技公司)纯化;纯化产物连接到pEASY-T1 Simple载体上(北京全式金生物技术有限公司),热激转化大肠杆菌感受态细胞,经过PCR扩增鉴定得到的阳性克隆送生工生物工程(上海)股份有限公司测序。
1.3.3 序列分析及系统进化树构建 测序结果采用DNAman和DNAstar 7.01进行去载体和序列拼接后,在NCBI网站进行blastn比对。同时从GenBank下载孢囊线虫其他近缘种的ITS序列和D2-D3区序列,用MEGA 5.0软件,邻接法(neighbor-joining method,NJ)构建该线虫与其他孢囊线虫属ITS序列和D2-D3区序列系统进化树[23,24]。
1.4 寄主范围测定
11种豆科作物21个品种(种子购自兴农种业公司)分别播种到PVC管(直径5 cm、高25 cm)中,播种和人工接种方法如1.1所述,接种量为每株接种1 mL 1 200粒/mL的卵悬浮液,每个处理4次重复,设感病大豆(婺源罗汉豆)为对照,温室条件下常规栽培管理。在接种后7 d参照BYBD等[25]的方法取其中两株进行根系品红染色,显微镜下观察线虫侵染情况。在接种后60 d取剩余两株,采用过筛法收集根系和土壤中孢囊于滤纸上统计孢囊数量,计算每个品种剩余两株的平均孢囊数。1.5 抗病性测定
同1.4中的方法种植40个大豆栽培品种(来自黑龙江、辽宁、河北、河南、江西等省份)并参照1.1接种野生豆孢囊线虫卵粒,每株接种1 mL 1 200粒/mL的卵悬浮液,每个品种设置4次重复,设置感病品种(婺源罗汉豆)为对照,在日光温室25—28℃条件下常规栽培管理。在接种后第60天取PVC管采用过筛法收集根系和土壤中孢囊于滤纸上统计孢囊数量,计算每个品种各重复的平均孢囊数和孢囊指数(公式1)并对其进行抗性评价。孢囊指数=(供试品种的平均孢囊量/感病对照品种的平均孢囊量)×100% (1)
按 SCHMITT等[26]提出的鉴定大豆抗病性的IP标准进行分级:高抗0—9%(VR),中抗 10%—30%(MR),中感31%—60%(MS),高感>60%(VS)。
2 结果
2.1 形态学观察与测量值
显微镜下对孢囊、2龄幼虫(J2)和雄虫的形态学进行观察和测量,形态学测量和文献记述比较见表1。中国江西大豆根上孢囊线虫群体的描述如下。Table 1
表1
表1野生豆孢囊线虫中国江西群体形态特征测量值
Table 1Morphometrics of H. sojae from Jingxi, China (μm)
| 测量特征 Population character | 样本量 Number | 江西群体 Jiangxi population | 样本量 Number | 韩国群体[15] Korea population[15] | |
|---|---|---|---|---|---|
| 孢囊 Cyst | n | n | |||
| 体长(不含颈长) Body length (excluding neck) (L) | 20 | 495.02±32.0 (447.6-571.5) | 25 | 513.4±70.4 (434.7-663.8) | |
| 体宽Body width (W) | 20 | 416.7±34.6 (345.3-457.0) | 25 | 448.8±56.2 (343.4-663.8) | |
| 阴门裂长Vulva slit length | 10 | 19.8±2.3 (16.1-22.8) | 10 | 18.1±4.5 (9.0-24.4) | |
| 膜孔长Fenestra length | 20 | 33.5±3.9 (28.1-43.2) | 10 | 42.0±9.3 (23.5-54.2) | |
| 膜孔宽Fenestra width | 20 | 26.9±4.5 (18.9-36.4) | 10 | 37.9±9.1 (22.4-51.0) | |
| 肛阴距Vulval-anal distance | 20 | 43.2±7.3 (30.5-58.8) | 10 | 40.9±11.0 (29.5-65.2) | |
| 半膜孔长Semifenestra length | 20 | 13.3±2.0 (10.6-19.6) | 10 | 15.2±1.9 (12.3-18.4) | |
| 2龄幼虫 J2 | |||||
| 体长Body length (L) | 20 | 425.7±35.5 (393.5-467.9) | 15 | 451.3±20.2 (411.7-477.8) | |
| 口针长Stylet length | 20 | 24.6±0.9 (23.0-26.2) | 15 | 24.5±0.4 (23.8-25.3) | |
| 唇高Lip region height | 20 | 5.3±0.6 (4.1-6.2) | |||
| 唇宽Lip region diameter | 20 | 10.5±0.7 (9.2-12.4) | |||
| 体前端到中食道球距离 Anterior end to median bulb | 20 | 74.7±5.0 (69.1-86.7) | |||
| 尾长 Tail length | 20 | 68.1±3.7 (61.8-74.7) | 15 | 65.8±5.9 (54.3-74.8) | |
| 透明尾长Tail hyaline region | 20 | 37.7±1.0 (36.0-39.4) | 15 | 38.7±3.9 (32.6-46.3) | |
| 雄虫 Male | |||||
| 体长Body length (L) | 5 | 1028.5±30.4 (974.8-1045.5) | 15 | 870.0±81.5 (668.3-967.5) | |
| 最大体宽Maximum body width (W) | 5 | 28.1±1.8 (26.7-31.0) | 15 | 23.0±1.9 (20.0-26.0) | |
| 体长/最大体宽L/W (a) | 5 | 39.0±1.5 (37.1-40.4) | 15 | 38.0±2.9 (32.8-42.0) | |
| 体前端到中食道球距离 Anterior end to median bulb | 5 | 82.8±8.9 (70.8-95.4) | |||
| 口针长Stylet length | 5 | 26.7±1.4 (24.5-28.1) | 15 | 26.8±0.8 (25.4-28.7) | |
| 交合刺长Spicule length | 5 | 28.1±1.8 (26.7-31.0) | 15 | 27.1± 2.1 (23.2-30.2) |
新窗口打开
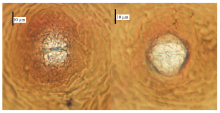
孢囊:虫体柠檬形或近圆形,与大豆孢囊线虫相比更圆,阴门锥突出较不明显;雌虫颜色白色,成熟以后孢囊呈深褐色或亮黑色,有明显的颈(图1-C、1-D);阴门区双半膜孔,无下桥和囊泡,极少存在卵囊团(图2)。J2:杀死固定时,虫体向腹部微弯,呈线形(图3-F);口针粗壮,口针基部球卵圆形,头部缢缩不明显,头架明显(图3-D);侧区4条侧线;尾圆锥形,尾端尖,透明尾长占尾长的53%—66%(图3-E、3-F)。雄虫:杀死固定时,虫体向腹部弯曲,轻微的呈弓形(图3-C)口针粗壮,口针基部球卵圆形,食道腺发育良好,有发达的椭圆形中食道球(图3-A)。交合刺明显,弯曲呈弓形。尾部很短,末端呈圆形(图3-B)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1大豆根系上的孢囊及白雌虫 A:根系上的孢囊Cyst on the roots;B:根系上的白雌虫Female on the roots;C、D:孢囊Cyst
-->Fig. 1Cyst and female on soybean roots
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2孢囊阴门锥特征
-->Fig. 2Characteristics of cyst vulva cone
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3大豆样品中2龄幼虫及雄虫在光学显微镜下主要形态特征 A:雄虫体前端 Anterior body portion of male;B:雄虫尾部 Tail of male;C:雄虫 Entire of male;D:2龄幼虫体前部 Anterior body portion of J2;E:2龄幼虫尾部 Tail of J2;F:2龄幼虫 Entire of J2
-->Fig. 3Photomicrographs of J2 and male from soybean samples
-->
2.2 分子生物学鉴定
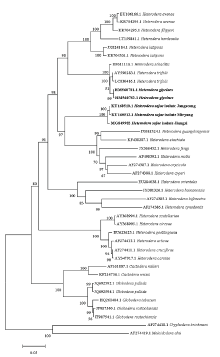
测序结果表明,孢囊线虫婺源群体的ITS的扩增片段长度为1 033 bp(GenBank登录号MG859982),28S D2-D3区片段长度755 bp(MG859981),Blast分析结果表明,该群体的ITS序列与GenBank收录野生豆孢囊线虫韩国群体的ITS序列(KU160510和KU160512)相似度分别为99%和98%,其次为麦类孢囊线虫(H. latipons)(HM560790),相似度为83%,与大豆孢囊线虫( KY794762.1)相似度只有81%。扩增得到D2-D3序列与野生豆孢囊线虫韩国群体的(KU160511)相似度最高为99%,与旱稻孢囊线虫(H. elachista)(HM560842),相似度为95%,与大豆孢囊线虫(LC208677)相似度为91%。从田间采集样本和温室扩繁的样本中获得的孢囊线虫相应序列完全一致,无任何单碱基差异。基于ITS序列和28S D2-D3序列构建的系统进化树分析结果表明(图4、图5),以ITS序列和D2-D3序列构建的发育树结果相一致,采集自江西大豆上的孢囊群体与来自韩国的野生豆孢囊线虫聚在同一支,置信度为100%,与旱稻孢囊线虫、广东孢囊线虫等莎草孢囊线虫群组(Cyperi)[27]聚在一个大的分支中,而大豆孢囊线虫与甜菜孢囊线虫、三叶草孢囊线虫(H. trifolii)等甜菜群组(Schachtii)[28]聚在另外一个分支。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4利用邻接法构建的江西孢囊线虫种群与其他孢囊线虫种的ITS序列系统进化树
-->Fig. 4Phylogenetic tree of Jiangxi population and other cyst nematodes based on ITS sequences using the neighbor-joining (NJ) method
-->
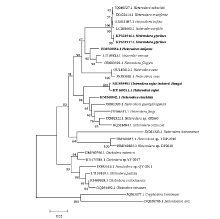 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5利用邻接法构建的江西孢囊线虫种群与其他孢囊线虫种的LUS D2-D3序列系统进化树
-->Fig. 5Phylogenetic tree of Jiangxi population and other cyst nematodes based on LUS D2-D3 sequences using the neighbor-joining (NJ) method
-->
2.3 寄主范围测定
11种豆科作物21个品种在人工接种野生豆孢囊线虫后7 d,经染色观察发现J2能够侵染大豆、相思豆、豇豆、豌豆、扁豆、绿豆、赤小豆、刀豆、四季豆和苜蓿10种豆科作物,但是只有在大豆和相思豆(红豆)的根系上能够发现豆荚形的3龄幼虫(J3),而在其他豆科作物上发现J2的侵染点都产生了较为严重的褐变(图6)。在接种后60 d经过检测,仅在大豆和相思豆(红豆)上能够产生孢囊,完成生活史(表2)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6不同豆科作物在接种后7 d根中2龄幼虫的侵染情况 a:大豆Glycine max;b:相思豆Abrus precatorius;c:豇豆Vigna unguiculata;d:豌豆Pisum sativum;e:扁豆Lablab purpureus;f:绿豆Vigna radiata;g:赤豆Vigna angularis;h:刀豆Canavalia gladiata;i:菜豆Phaseolus vulgaris;j:苜蓿Medicago sativa
-->Fig. 6J2 infections in different legume crop roots after 7 days of inoculation
-->
Table 2
表2
表2野生豆孢囊线虫江西群体对豆科作物的寄主范围测定
Table 2Parasitism test of H. sojae from Jiangxi on legume crops
| 植物种类 Species | 品种 Cultivar | 接种7 d侵染情况 Infections after inoculation 7 days | 接种60 d孢囊的产生情况 Cysts produced after inoculation 60 days |
|---|---|---|---|
| 大豆 Glycine max | 婺源罗汉豆Wuyuanluohan bean | 有2龄幼虫侵染Infection with J2 | 产生26个孢囊Produce 26 cysts |
| 黑豆Black bean | 有2龄幼虫侵染Infection with J2 | 产生14个孢囊Produce 14 cysts | |
| 豇豆 Vigna unguiculata | 红条无柴Hongtiaowucai cowpea | 有2龄幼虫侵染Infection with J2 | 无 None |
| 江南豇豆Jiangnan cowpea | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 五月红豇豆Wuyuehong cowpea | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 高产4号豇豆Gaochan4 cowpea | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 刀豆Canavalia gladiata | 武皇精选刀豆Wuhuang selection sword bean | 有2龄幼虫侵染Infection with J2 | 无 None |
| 豌豆Pisum sativum | 韩育荷兰豆Hanyu pea | 有2龄幼虫侵染Infection with J2 | 无 None |
| 白荷兰豆 Sweet broad pea | 未侵染Uninfected | 无 None | |
| 甜脆豌豆Tiancui pea | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 豌豆Pea | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 扁豆Lablab purpureus | 紫扁豆Purple lentils | 有2龄幼虫侵染Infection with J2 | 无 None |
| 绿眉豆Green brown bean | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 蚕豆Vicia faba | 蚕豆Broad bean | 未侵染Uninfected | 无 None |
| 大志蚕豆Dazhi broad bean | 未侵染Uninfected | 无 None | |
| 绿豆Vigna radiata | 黑绿豆Black green bean | 有2龄幼虫侵染Infection with J2 | 无 None |
| 早熟绿豆Early-maturing green bean | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 菜豆Phaseolus vulgaris | 架四季豆Kidney bean | 有2龄幼虫侵染Infection with J2 | 无 None |
| 无架地豆Wujia kidney bean | 有2龄幼虫侵染Infection with J2 | 无 None | |
| 赤豆Vigna angularis | 赤豆Red bean | 有2龄幼虫侵染Infection with J2 | 无 None |
| 相思豆 Abrus precatorius | 相思豆(红豆)Jequirity | 有2龄幼虫侵染Infection with J2 | 产生6个孢囊Produce 6 cysts |
| 紫苜蓿Medicago sativa | 紫苜蓿 Alfalfa | 有2龄幼虫侵染Infection with J2 | 无 None |
新窗口打开
2.4 抗病性测定
40份大豆栽培品种中对野生豆孢囊线虫表现高抗的品种有5个,中抗的品种有5个,中感品种有11个,高感品种有19个。结果表明参鉴品种中75%的大豆栽培品种对来自江西的野生豆孢囊线虫表现感病(表3)。Table 3
表3
表3大豆栽培品种对野生豆孢囊线虫江西种群抗性鉴定
Table 3Soybean cultivars resistant identification to H. sojae from Jiangxi
| 品种名称 Cultivar | 鉴定结果 Identification result | ||
|---|---|---|---|
| 平均单株孢囊个数Number of cysts per plant | 孢囊指数Cyst index | 抗性等级Resistance level | |
| 中黄12 Zhonghuang 12 | 1±0.58 | 1 | 高抗VR |
| 中黄27 Zhonghuang 27 | 3±0.58 | 2 | 高抗VR |
| 辽豆13 Liaodou 13 | 8±8.62 | 4 | 高抗VR |
| PI437654 | 12±15.62 | 7 | 高抗VR |
| 丰豆8号 Fengdou 8 | 14±10.66 | 8 | 高抗VR |
| 中黄13 Zhonghuang 13 | 18±10.21 | 10 | 中抗MR |
| 中黄38 Zhonghuang 38 | 30±31.34 | 16 | 中抗MR |
| 十月青 Shiyueqing | 36±8.54 | 20 | 中抗MR |
| 辽豆14 Liaodou 14 | 40±32.60 | 22 | 中抗MR |
| 赣豆 Gandou | 46±18.34 | 25 | 中抗MR |
| 垦丰8 Kenfeng 8 | 70±50.80 | 38 | 中感MS |
| 垦豆23 Kendou 23 | 71±64.49 | 39 | 中感MS |
| 中作RNO83 Zhongzuo RNO83 | 72±33.50 | 40 | 中感MS |
| 沧豆5号 Cangdou 5 | 76±28.28 | 42 | 中感MS |
| 辽豆34 Liaodou 34 | 85±25.17 | 47 | 中感MS |
| 垦豆26 Kendou 26 | 86±38.54 | 47 | 中感MS |
| 中豆43 Zhongdou 43 | 89±59.56 | 49 | 中感MS |
| 垦农26 Kennong 26 | 97±13.69 | 53 | 中感MS |
| 菏豆19 Hedou 19 | 98±23.26 | 54 | 中感MS |
| 垦豆41 Kendou 41 | 106±96.23 | 58 | 中感MS |
| 沧豆21 Cangdou 21 | 109±65.43 | 60 | 中感MS |
| 辽豆32 Liaodou 32 | 113±91.08 | 62 | 高感VS |
| 开育12 Kaiyu 12 | 119±30.92 | 65 | 高感VS |
| 垦豆38 Kendou 38 | 120±54.01 | 66 | 高感VS |
| 中黄42 Zhonghuang 42 | 126±50.71 | 69 | 高感VS |
| 翠豆43 Cuidou 43 | 135±51.42 | 74 | 高感VS |
| 中黄35 Zhonghuang 35 | 138±51.23 | 76 | 高感VS |
| 辽豆33 Liaodou 33 | 143±40.50 | 79 | 高感VS |
| 邯豆3号Handou 3 | 149±70.44 | 82 | 高感VS |
| 早熟1号 Zaoshu 1 | 162±41.32 | 89 | 高感VS |
| 美F 88-60 Mei F 88-60 | 165±98.78 | 91 | 高感VS |
| 邯豆10号 Handou 10 | 165±54.66 | 91 | 高感VS |
| 徐豆9号 Xudou 9 | 184±10.61 | 101 | 高感VS |
| 中黄51 Zhonghuang 51 | 190±19.66 | 104 | 高感VS |
| 中黄26 Zhonghuang 26 | 205±88.84 | 113 | 高感VS |
| 垦农30 Kennong 30 | 213±177.21 | 117 | 高感VS |
| 黑农51 Heinong 51 | 228±53.93 | 125 | 高感VS |
| 中黄43 Zhonghuang 43 | 231±24.68 | 127 | 高感VS |
| 翠扇Cuishan | 240±94.98 | 132 | 高感VS |
| 垦丰9 Kenfeng 9 | 268±56.43 | 147 | 高感VS |
| 婺源罗汉豆 CK | 182±41.93 | — | — |
新窗口打开
3 讨论
形态学观察和测量结果表明,采集自中国江西大豆根上的孢囊线虫群体与韩国的野生豆孢囊线虫[16]形态学特征基本一致。江西群体的孢囊个体较韩国群体孢囊略小(中国江西种群/韩国种群:体长495.0 μm/513.0 μm,体宽448.0 μm/416.0 μm)。江西群体与韩国群体就阴门锥特征相比膜孔较小,中国江西群体/韩国群体:膜孔长33.0 μm/41.0 μm,膜孔宽26.0 μm/37.0 μm)。可能是生态环境、地域差异造成,大豆品种的不同也可能对群体分化存在一定的影响[29];中国江西群体2龄幼虫(J2)、雄虫与韩国群体J2、雄虫相比形态学特征测量结果较为一致(表1)。孢囊形态观察韩国群体没有能够观察到卵囊团[16],而中国江西群体极少但能观察到卵囊团。形态学鉴定结果认为江西大豆根上孢囊线虫群体与韩国野生豆孢囊线虫一致,而与大豆孢囊线虫相比有较大差别,野生豆孢囊线虫的孢囊形态更圆,表皮有光泽,无下桥和囊泡。野生豆孢囊线虫于2016年由KANG等[16]在韩国大豆上发现并首次报道,目前还未在其他国家和地区被发现报道,也未见关于其致病性与寄生性方面的报道。在研究野生豆孢囊线虫对豆科作物寄主范围测定中,通过根系染色观察到在J2侵染时期,在非寄主或不良寄主的根系内J2侵染点附近会产生较为严重的褐化现象(图6),可能是因为线虫侵染导致的免疫反应,使得孢囊线虫不能建立取食位点,最终造成其不能完成生活史。栽培大豆品种的抗性鉴定结果表明,在参与鉴定的40个大豆栽培品种中感病品种所占比例较高,目前国内多数大豆栽培品种对该孢囊线虫没有抗性。国内外的生产实践表明在大豆孢囊线虫病的防治中,培育抗病品种是最经济有效的方法[30],因此通过筛选对野大豆孢囊线虫具有抗性的大豆种质资源具有重要意义。
大豆孢囊线虫病在黑龙江、吉林、辽宁、内蒙古、山西、山东、河南、河北、安徽、江苏、浙江、广西和江西等省(自治区)均有不同程度发生[31]。此次报道的野生豆孢囊线虫系我国寄生于大豆上的孢囊线虫新记录种,目前仅在韩国和中国江西被发现。我国其他省份是否有该孢囊线虫发生分布还有待于进一步调查明确。鉴于韩国和中国江西两地纬度跨度较大,涵盖了我国东北、华北和黄淮海等主要的大豆种植区,与大豆孢囊线虫的主要发生分布区域一致[31],并且本研究在种群扩繁时,在北京夏天室外盆栽种植大豆接种后该孢囊线虫也能够较好的寄生和完成生活史,综合以上情形推测该孢囊线虫对于环境有较好的适应性,在我国大豆主产区能够生存繁殖,因此有必要引起重视。
4 结论
依据形态学和分子生物学特征,将采自中国江西大豆上的孢囊线虫群体鉴定为野生豆孢囊线虫,系中国一新记录种;寄主范围测定结果表明大豆和相思豆(红豆)是其适宜寄主;对野生豆孢囊线虫的抗性鉴定表明,40个国内大豆栽培品种中有19个高感、11个中感、5个中抗、5个高抗。(责任编辑 岳梅)
The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}