0 引言
【研究意义】冷害是热带亚热带果蔬贮运保鲜中存在的主要问题之一[1]。热处理是提高果蔬抗冷性的最有效采后处理之一[2],但热处理诱导的抗冷机制特别是有关的信号传导途径目前还不太清楚。模式植物上的研究表明,低温转录因子CBF/DREB调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径[3-5],但是有关CBF冷应答途径在热处理诱导抗冷性中的作用目前很少报道。对CBF冷应答途径在热处理诱导香蕉抗冷性中的作用展开深入研究,将为了解香蕉果实中CBF冷应答途径的信号转导过程提供参考。【前人研究进展】模式植物上的研究表明,低温转录因子CBF调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径,即ICE-CBF-COR通路。CBF是该途径中基因表达的“主开关”,受ICE调控的CBF转录因子能够特异结合启动子中含有CRT/DRE(C-repeat/ dehydration responsive element)的顺式元件,可以激活下游一系列冷调节基因COR的表达,从而提高拟南芥等植物的抗寒性[6-8]。高温和低温均能诱导CBF冷应答途径相关基因的表达。4℃低温诱导了蝴蝶兰PaCBF1在4 h开始表达,24 h达到最高峰。在拟南芥中过表达PaCBF1,诱导了AtCOR6.6和RD29a表达的增强[9]。桃果实经过0℃低温处理6 h时,PpCBF5表达水平增加约4倍,然后迅速增加至72 h的4 000多倍。PpCBF1和PpCBF6的表达,在0℃下72 h可分别被诱导575倍和6倍[10]。38℃热处理1 h诱导了黄瓜幼苗CBF1的表达,同时增加了其抗热性[11];高温胁迫诱导了拟南芥中DREB2A蛋白的产生,此蛋白作为一个调控因子参与拟南芥的热激胁迫反应[12]。高温和低温都能诱导新疆野苹果MsDREB2C [13]、玉米ZmDREB2A[14]的表达增强。【本研究切入点】笔者前期研究结果表明,采用52℃热水处理香蕉果实3 min,能有效减轻香蕉果实的冷害症状[15-17]。然而热水处理是否通过CBF冷应答途径提高香蕉果实的抗冷性,目前尚不清楚。【拟解决的关键问题】研究香蕉果实在低温下是否存在CBF冷应答途径,以及CBF冷应答途径中的相关基因是否参与了热水处理诱导的香蕉抗冷性。1 材料与方法
试验于2015年在华南农业大学园艺学院进行。1.1 材料及处理
香蕉品种为‘巴西’(Musa spp. AAA group cv. Brazil),采自广州市番禺香蕉园。挑选饱满度约7成的绿熟香蕉,运回广东省果蔬保鲜重点实验室。挑选大小均匀、无病虫害及机械损伤的单个香蕉,先后用0.1%漂白粉和0.05%施保功各浸泡5 min,晾干备用。热水及低温贮藏处理:将香蕉果实一次性浸入热水处理机(体积为400 L)中,温度为52℃,时间为3 min,热水处理前后温度差异在0.5℃以内。热水处理(hot water dipping,HWD)后将香蕉果实放入20℃恒温箱中贮藏3 h(前期研究表明热水处理后于20℃恢复3 h抗冷效果最好[16]),然后置于7℃下贮藏5 d,对照为未经过热水处理,其他贮藏条件一样。热处理香蕉100个,对照处理100个,每处理3个重复。取样时间点为:20℃下0、0.5、1.5、3 h;7℃下1、4、8、24、48、120 h。每个取样点10个香蕉,取香蕉中部果皮用液氮冷冻置于-80℃冰箱保存。
1.2 测定方法
1.2.1 实时定量引物设计 从香蕉基因组数据库(http://banana-genome.cirad. fr/)中搜索并挑选CBF冷信号途径中的7个相关基因序列,设计实时定量引物,验证后备用。引物序列为:MaICE:5-TTGAGGTCA GGGTAAGAG和3-CTTCCTTGGATTGCTCAG;MaDREB1D:5-CATGGATTTG GGCTTCGGTTAC和3-GTCCACGTCGTCCCAGTTACA;MaDREB1E:5-CGATGAATGAGGAGGACGAGAT和3-CACC AAACTGATGCTGGAGGA;MaDREB1G:5-AACT TCGGAATGCAAGGGTA和3-CCGGTCTCAGATGG AATAGC;MaDREB2:5-GACGAGCACCAGCGA GTCAAG和3-AGAGCGGAATGTCACTGAAAGC;MaDREB3:5-CGAGTTCCTGTACGAAGACCCG和3-AGATGGTGGTGGGAACAACAGTG;MaCOR413:5-CTGACTGGTTGGAGATGC和3-TACCAAGCGCC AGATAGG。1.2.2 实时荧光定量PCR(qRT-PCR) 实时荧光定量PCR使用仪器为BIO-RAD CFX9600,采用TOYOBO THUNDERBIRD SYBR® qPCR Mix 进行Real-time PCR扩增。qRT-PCR反应体系(20 μL)包括10 μL SYBR-Green PCR Master Mix,0.25 μL上游(10 μmol·L-1)和0.25 μL下游引物(10 μmol·L-1),2 μL稀释的cDNA以及7.5 μL ddH2O。反应条件为:95℃ 3 min,95℃ 10 s,59℃ 10 s,72℃ 20 s,循环数为40。为了确认产物的质量和引物的特异性,在溶解曲线中分析了产物的Tm(分析溶解温度为65—95℃)。利用Actin1[18]作为内参基因。所有的qRT-PCR反应根据内参基因Actin1进行Ct值校准,目的基因的相对表达水平利用公式2-ΔΔCt进行计算,重复3次。
1.2.3 数据处理 用MEGA5.1软件对氨基酸序列构建系统进化树,采用Sigmaplot12.5计算标准误并作图。
2 结果
2.1 香蕉DREB/CBF基因的系统进化树分析
从香蕉基因组数据库搜索到5个DREB类基因:MaDREB1D、MaDREB1E、MaDREB1G、MaDREB2C 和MaDREB3。参考SAKUMA[19]、MIZOI[20]等的研究结果,从拟南芥、水稻等物种选取已克隆的DREB/CBF转录因子的氨基酸序列,用MEGA5.1软件构建系统进化树。从图1可知,MaDREB1E和MaDREB1G属于DREB/CBF 转录因子A-1亚类,MaDREB1D属于A-6亚类、MaDREB2C属于A-3亚类、MaDREB3属于A-4亚类。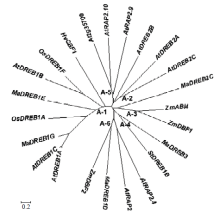 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1植物中DREB类基因的系统进化树
-->Fig. 1Phylogenetic tree of the DREB subfamily transcription factors in plants
-->
2.2 热激处理及热激后低温贮藏对香蕉MaICE表达的影响
前期研究表明,7℃(冷害温度)下,香蕉果实在第5天已经出现明显的冷害症状,利用52℃热水处理(HWD)3 min能有效提高香蕉果实在7℃低温贮藏中的抗冷性[16-17]。香蕉用52℃热水处理(HWD)3 min ,然后在20℃下放置3 h后置于7℃下贮藏5 d。从图2可知,未经热处理的香蕉(对照)MaICE表达在7℃处理1 h时迅速升高达到最大值,之后又迅速降低为低温前的水平。香蕉经热处理后的0.5 h,其MaICE表达迅速升高,之后又下降到对照水平,7℃低温贮藏下8 h有一个高峰。说明香蕉MaICE在20℃下表达量很低,7℃低温能在1 h内快速诱导MaICE表达的升高。热处理能在0.5 h内快速诱导MaICE表达的升高,而7℃低温下出现高峰的时间从1 h推迟到8 h。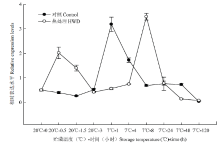 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2热处理和低温贮藏对香蕉果实MaICE表达的影响
-->Fig. 2Effect of hot water dipping (HWD) and cold storage on the expression of MaICE in banana fruits
-->
2.3 热激处理及热激后低温贮藏对香蕉DREB类基因表达的影响
从图3—图5可知,未经热处理的香蕉MaDREB1D、MaDREB1E和MaDREB1G这3个基因表达均在7℃1 h有一个高峰。说明7℃低温能在1 h内快速诱导这3个基因表达的升高。MaDREB1D、MaDREB1E和MaDREB1G这3个基因表达均在热处理后0.5 h有一个高峰。在7℃低温贮藏过程中,HWD处理的MaDREB1D表达一直高于对照,MaDREB1E表达在4—24 h高于对照,而MaDREB1G表达与对照差异不大。从图6可知,对照的MaDREB2C表达在7℃下4 h后逐渐增强,直至24 h有一个高峰,之后逐渐下降至较低的水平。而HWD处理后的0.5 h,其MaDREB2C表达有一个短暂而迅速的升高,低温下其MaDREB2C表达变化趋势与对照一致,并且均高于对照。
从图7可知,对照的MaDREB3表达在7℃下1 h有一个高峰,而HWD处理后MaDREB3表达在HWD后0.5 h有一个小高峰,表达比对照稍高。在7℃低温贮藏过程中,MaDREB3表达在前期低于对照,后期则高于对照。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3热处理和低温贮藏对香蕉果实MaDREB1D表达的影响
-->Fig. 3Effect of HWD and cold storage on the expression of MaDREB1D in banana fruits
-->
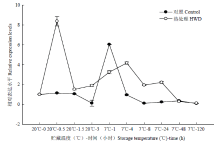 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4热处理和低温贮藏对香蕉果实MaDREB1E表达的影响
-->Fig. 4Effect of HWD and cold storage on the expression of MaDREB1E in banana fruits
-->
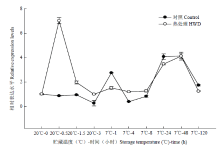 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5热处理和低温贮藏对香蕉果实MaDREB1G表达的影响
-->Fig. 5Effect of HWD and cold storage on the expression of MaDREB1G in banana fruits
-->
2.4 热激处理及热激处理后低温贮藏对香蕉MaCOR413表达的影响
从图8可知,对照的MaCOR413表达在7℃下4 h有一个高峰,其他时间的表达量均较低。而HWD的MaCOR413表达在热处理后0.5 h有一个高峰,在7℃下4 h有一个高峰,整个过程,HWD的MaCOR413表达量均高于对照。3 讨论
多个植物物种中的研究表明,低温转录因子CBF调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径。DREB/CBF转录因子是AP2/EREBP转录因子家族中的1个亚家族,含有1个保守的AP2/EREBP结构域。拟南芥基因组DREB/CBF亚族又分为6个亚类(A1—A6)[19]。DREB1/CBF类主要受到低温诱导,DREB2类主要受到干旱和高盐诱导,而对于A3—A6目前研究较少。从图1的系统进化树发现香蕉的5个DREB基因属于不同的亚类。众多研究证实,CBF/DREB类基因在多种高等植物低温下的表达模式均是在低温下短时间内快速诱导增强,最终通过诱导COR表达增强从而提高植物的抗冷性。例如,拟南芥2.5℃低温15 min即可诱导AtCBF1、AtCBF2和AtCBF3的表达,2 h后达到表达高峰,随后下降,同时在低温2 h后COR开始被诱导表达[21-22]。过量表达CBF1或CBF3可促进 COR高水平表达,使转基因拟南芥植株的抗冷性提高[23]。宽叶独行菜LICBF在4℃低温胁迫下1 h的表达迅速增强,之后4—48 h逐渐降低至低温处理前的水平[24]。本研究结果表明,香蕉果实(对照)的MaICE表达在7℃处理1 h时迅速升高达到最大值,DREB类基因(MaDREB1D、MaDREB1E、MaDREB1G、和MaDREB3)表达在7℃处理1 h均存在一个高峰,随后MaCOR413表达在7℃下4 h出现一个高峰。其中MaDREB1D的表达模式与其他DREB有所不同,除了在1 h有一个高峰,MaDREB1D在120 h(冷害症状明显)时还出现一个表达最高峰。采后香蕉果实在低温胁迫下的CBF冷应答基因表达模式与拟南芥类等植物类似,进一步验证了CBF冷应答途径在高等植物中保守性很高。将CBF冷应答途径基因转入拟南芥中,均提高了拟南芥的抗冷性。例如,ZHUANG等[25]将具有抗冷性草地早熟禾中的PpCBF3转化至拟南芥幼苗中,通过瞬时表达,发现4℃下0.5 h,PpCBF3表达开始增强,在1 h迅速最高峰,之后3—24 h迅速下降至较低的水平。进一步研究发现,4℃低温能诱导野生型和转基因拟南芥COR15a、COR6.6、COR47和COR78的表达,且转基因拟南芥中这些COR的表达高于野生型拟南芥。类似地,将中国野生葡萄中的VaICE1/2在拟南芥中过表达,增强了下游基因CBF1、COR15A和COR47的表达,提高了拟南芥的抗冷性[26]。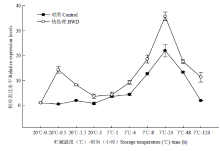 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6热处理和低温贮藏对香蕉果实MaDREB2C表达的影响
-->Fig. 6Effect of HWD and cold storage on the expression of MaDREB2C in banana fruit
-->
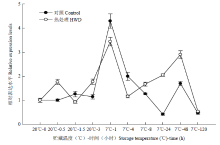 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7热处理和低温贮藏对香蕉果实MaDREB3表达的影响
-->Fig. 7Effect of HWD and cold storage on the expression of MaDREB3 in banana fruit
-->
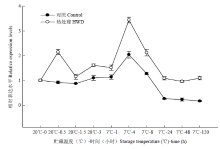 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8热处理和低温贮藏对香蕉果实MaCOR413表达的影响
-->Fig. 8Effect of HWD and cold storage on the expression of MaCOR413 in banana fruit
-->
香蕉果实MaDREB2C在低温下的表达模式与其他DREB类基因有所不同。香蕉果实(对照)在7℃低温胁迫下24 h才出现表达高峰,这与其他DREB类基因均在7℃低温1 h出现表达高峰不同。说明MaDREB2C在香蕉果实低温贮藏后期产生响应。以上结果表明,7℃低温胁迫下,MaICE、DREB类基因、MaCOR413表达依次增强,说明低温下香蕉果实中存在CBF冷应答途径,即ICE-CBF-COR通路。
热处理是提高采后果实抗冷性的有效手段,笔者前期研究结果表明,采用52℃热水处理香蕉果实3 min,能有效减轻香蕉果实的冷害症状[15-17]。同时,热处理也能够诱导CBF冷应答途径相关基因的表达。例如,40℃高温处理能诱导马铃薯植株中的AtCBF3表达增强,并且发现在40℃胁迫l h时,叶中AtCBF3表达量最大[27]。将军菊苣幼苗高温37℃处理1 h后,叶片和根中CiDREB2A表达量即上升,并一直维持高表达,24 h表达量达到峰值[28]。CAO等[29]利用低温、高温和干旱对巨桉处理,对17个EgrDREB1/CBF和6个EgrDREB2研究,发现高温能诱导所有基因的表达增强,低温能诱导16个EgrDREB1/CBF的表达,认为EgrDREB1/CBF亚家族主要是受低温诱导,而EgrDREB2主要是对干旱敏感。从本研究中的图2至图8可知,香蕉果实经过热处理后,CBF冷应答途径相关的7个基因表达均在热处理后的0.5 h有一个短暂而迅速的升高,在3 h恢复到处理前较低的水平。香蕉果实CBF冷应答途径的7个基因均在热处理后0.5 h的迅速表达增强,说明香蕉果实CBF冷应答途径的相关基因能受到热处理诱导快速表达增强。
进一步研究发现,热激处理后在低温贮藏时,CBF冷应答途径相关基因也会受到诱导增强。猕猴桃果实AcCBF在冷藏期间的相对表达量呈现先升高后下降的趋势,表达高峰在贮藏20 d时出现。而利用35℃和45℃热处理与对照相比显著提高了AcCBF的表达峰值[30]。香蕉果实经热处理后于7℃低温贮藏5 d,发现MaDREB1D、MaDREB1E、MaDREB2C、MaDREB3和MaCOR413等基因的表达均强于对照,MaICE表达出现峰值的时间从1 h(对照)推迟到8 h,而MaDREB1G表达与对照差异不大。以上研究表明,香蕉果实经过热处理后,7个CBF冷应答途径基因在热处理后0.5 h的迅速表达增强,可能是作为一个早期信号,诱导了后期CBF冷应答途径相关基因的进一步表达增强,导致香蕉果实抗冷性增强。这说明热处理诱导采后香蕉果实的耐冷性可能与CBF冷应答途径密切相关。目前有关热处理诱导CBF冷应答途径相关基因表达方面的研究报道很少,热处理诱导的CBF冷应答途径相关基因表达提高与植物抗冷性之间的关系需要进一步研究。
4 结论
7℃低温胁迫下,MaICE、DREB类基因、MaCOR413表达依次增强,说明低温下香蕉果实中可能存在CBF冷应答途径,即ICE-CBF-COR通路。热激处理能快速诱导CBF冷应答途径相关基因的表达,并且DREB类基因和MaCOR413在7℃低温胁迫下表达强于对照。推测CBF冷应答途径相关基因表达增强可能参与了热处理诱导采后香蕉果实的抗冷性。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}