Correlation Analysis of Auxin Involved in the Process of Petal Abscission of Tree Peony Luoyanghong Cut Flowers by Ethylene Promoting
YE Di,1, SHI Jiang1, GAO ShuangCheng1, WANG ZhanYing2, SHI GuoAn,11College of Mudan, Henan University of Science and Technology/Luoyang Key Laboratory of Peony Biology, Luoyang 471023, Henan 2Research Institute of Peony, Luoyang Academy of Agriculture and Forestry Sciences, Luoyang 471000, Henan
Abstract 【Objective】The aim of this study was to explore the physiological mechanisms of petal wilting and abscission during the senescence of cut tree peony flowers, so as to provide the theoretical basis for better development of storage and preservation technology of cut tree peony flowers.【Method】The cut flowers were pretreated with deionized water (CK), 20 µL·L-1 ethylene antagonist Correxian (AVB), 20 µL·L-1 ethylene releasing agent ethephon (CEPA), and 20 µL·L-1 Correxian for 1 h, respectively, followed by 20 µL·L-1 ethephon 1 h (AVB+CEPA). The flowers were then placed in individual vases filled with distilled water. The morphology of the petals absciss zone cells were observed by using a microscope. The petal anti-falling ability was determined by using the tensile test during the vase cutting process. The physiological effects of different pretreatments were observed respectively using the physiological and biochemical methods to determine the ethylene and auxin metabolism, the absciss zone hydrolysis-related enzyme activities, and the relative expressions of PsETR1, PsCTR1, PsEIN3, PsERF1, PsYUCCA10, PsPIN1, PsPME1, PsPG1 and PsBG1 genes. 【Result】 The results showed that the basal petal of the cut tree peony Luoyanghong had a clear separation layer structure. AVB significantly delayed the occurrence of ethylene climacteric in cut flowers, and decreased the ethylene release peak by 34.9% (P<0.05), while the petal anti-falling ability was improved. The activities of polygalaturonase (PG), β-glucosidase (BG) and the expressions of PsPG1 and PsBG1 genes at the base of the petals absciss zone were reduced, which delayed the senescence process of cut tree peony flowers, thus significantly extended its vase life. Nevertheless, the exogenous CEPA significantly accelerated the release rate of endogenous ethylene in the petals, increased the activities of IAA oxidase (IAAO), PG and BG, accelerated the senescence of the absciss zone cells in the petals of cut flowers, and reduced the petal anti-falling ability, thus promoted its wilting and falling. Meanwhile, the vase life and the optimal viewing period of AVB+CEPA composite pretreatment were not much different from control, so CEPA could partially offset the physiological effects of AVB. 【Conclusion】 For the ethylene sensitive cut tree peony Luoyanghong, endogenous auxin was involved in the process of ethylene promoting the abscission of petal absciss zone cells. The control of ethylene was the basic way to improve the quality of Luoyanghong cut flowers. Keywords:tree peony;cut flowers;absciss zone;ethylene;auxin;abscission
PDF (1766KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 叶迪, 施江, 高双成, 王占营, 史国安. 乙烯促进牡丹‘洛阳红’切花花瓣脱落与内源生长素的关联性分析. 中国农业科学, 2021, 54(23): 5097-5109 doi:10.3864/j.issn.0578-1752.2021.23.014 YE Di, SHI Jiang, GAO ShuangCheng, WANG ZhanYing, SHI GuoAn. Correlation Analysis of Auxin Involved in the Process of Petal Abscission of Tree Peony Luoyanghong Cut Flowers by Ethylene Promoting. Scientia Acricultura Sinica, 2021, 54(23): 5097-5109 doi:10.3864/j.issn.0578-1752.2021.23.014
MAN, MAC, LIUY, SHAHID MO, WANG CP, GAO JP. Petal senescence: A hormone view Journal of Experimental Botany, 2018, 69(4):719-732. DOI:10.1093/jxb/ery009URL [本文引用: 1]
DANAEEE, MOSTOFIY, MORADIP. Effect of GA3 and BA on postharvest quality and vase life of Gerbera (Gerbera jamesonii. cv. Good Timing) cut flowers Horticulture, Environment, and Biotechnology, 2011, 52(2):140-144. DOI:10.1007/s13580-011-1581-7URL [本文引用: 1]
LIAO WB, ZHANG ML, HUANG GB, YU JH. Hydrogen peroxide in the vase solution increases vase life and keeping quality of cut Oriental×Trumpet hybrid lily ‘Manissa’ Scientia Horticulturae, 2012, 139(139):32-38. DOI:10.1016/j.scienta.2012.02.040URL [本文引用: 1]
SU XY, LI WJ. Influence of different preservatives formula on preservation effect of carnation cut flower Northern Horticulture, 2014(1):133-136.(in Chinese) [本文引用: 1]
FANGH, WANG CL, LIAO WB, ZHANGJ, HUO JQ, HUANG DJ, NIU LJ, WANGB. NO is involved in ABA-regulated senescence of cut roses by maintaining water content and increasing antioxidant enzyme activities Acta Horticulturae Sinica, 2019(5):901-909.(in Chinese) [本文引用: 1]
SHIJ, TIANZ, SHI GA. Effect of exogenous hydrogen peroxide or ascorbic acid on senescence in cut flowers of tree peony (Paeonia suffruticosa Andr.) Journal of Horticultural Science & Biotechnology, 2015, 90(6):689-694. [本文引用: 2]
ZHOUL, JIA PY, LIUJ, WANG WR, HUO ZP, DONGL. Effect of ethylene on cut flowers of tree peony ‘Luoyang Hong' opening and senescence process and endogenous ethylene biosynthesis Acta Horticulturae Sinica, 2009(2):239-244. (in Chinese) [本文引用: 1]
SHI GA, YANG ZS, WANG CZ, JIN ZW, HAN XY. Effects of temperature and chemical agents on ethylene release and storage quality of cut peony flowers Northern Horticulture, 1997, 21(6):62-63. (in Chinese) [本文引用: 2]
SHI GA, GUO XF, HAN JG, SUN XM, YANG ZS. A study on ethylene production and lipid peroxidization in florescence and flower senescence of Paeonia suffruticosa Acta Universitatis Agriculturalis Boreali-Occidentalis, 1999, 27(5):50-53. (in Chinese) [本文引用: 2]
SHI GA, GUO XF, ZHANG GH, BAO MZ. Analysis of sugar metabolism during florescence and flower senescence of tree peony petal Acta Horticulturae Sinica, 2009, 36(8):1184-1190. (in Chinese) [本文引用: 2]
SHI GA, GUO XF, LI CL, FAN BY, SHIJ, BAO MZ. Changes on ethylene release and ACC content of peony flowers at different development stages Acta Horticulturae Sinica, 2010, 37(1):77-82. (in Chinese) [本文引用: 1]
SHI GA, GUO XF, ZHANG GH, GAO SC, FAN BY, BAO MZ. Studies of vasing physiological characteristics of cut peony flowers in various development stages Acta Horticulturae Sinica, 2010, 37(3):449-456. (in Chinese) [本文引用: 1]
ZHANGC, LIUM, FU JX, WANG YJ, DONGL. Exogenous sugars involvement in senescence and ethylene production of tree peony ‘Luoyang Hong’ cut flowers Korean Journal of Horticultural Science and Technology, 2012, 30(6):718-724. DOI:10.7235/hort.2012.12089URL [本文引用: 1]
GAO SL, ZHANGC, DU DN, LIU AQ, DONGL. Effect of glucose and ethylene on flower color and anthocyanin biosynthesis in tree peony ‘Luoyang Hong’ cut flower Acta Horticulturae Sinica, 2015, 42(7):1356-1366. (in Chinese) [本文引用: 1]
WANG KX, WANGY, SHIT, DING XN, YUAN JH, SHI GA, HU YH. Physiological effects of rapamycin pretreatment on delaying the senescence of cut flower in tree peony ‘Luoyanghong’ Acta Horticulturae Sinica, 2020, 47(10):1956-1968. (in Chinese) [本文引用: 1]
PATTERSON SE, BLEECKER AB. Ethylene-dependent and -independent processes associated with floral organ abscission in Arabidopsis Plant Physiology, 2004, 134(1):194-203. DOI:10.1104/pp.103.028027URL [本文引用: 3]
ROBERTS JA, ELLIOTT KA, GONZALEZ-CARRANZAZ H. Abscission, dehiscence, and other cell separation processes Annual Review of Plant Biology, 2002, 53:131-158. DOI:10.1146/arplant.2002.53.issue-1URL [本文引用: 2]
MICHAELIR, PHILOSOPH-HADASS, RIOVJ, SHAHAKY, RATNERK, MEIRS. Chillinginduced leaf abscission of Ixora coccinea plants. III. Enhancement by high light via increased oxidative processes Physiologia Plantarum, 2010, 113(3):338-345. DOI:10.1034/j.1399-3054.2001.1130306.xURL [本文引用: 1]
PATHARKAR OR, WALKER JC. Floral organ abscission is regulated by a positive feedback loop Proceedings of the National Academy of Sciences of the USA, 2015, 112(9):2906-2911. [本文引用: 2]
CUI YS, CHEN QF, HUO DA, LI HY. Research progress on molecular mechanism of plant falling flowers and fruits Guihaia, 2018, 38(9):1234-1247. (in Chinese) [本文引用: 2]
ZHENG LY, WEIX, ZHOUK, XIANGJ, LIY, LIU BY, HE GH, LING YH, ZHAO FM. Identification of a rice chromosome segment substitution line Z481 carrying a major gene for seed shattering and mapping of SH6(t) Chinese Science Bulletin, 2016, 61(7):748-758. (in Chinese) [本文引用: 1]
GAO YR, LIUY, LIANGY, LU JY, JIANG CY, FEI ZJ, JIANG CZ, MAC, GAO JP. Rosa hybrida RhERF1 and RhERF4 mediate ethylene- and auxin-regulated petal abscission by influencing pectin degradation The Plant Journal, 2019, 99(6):1159-1171. DOI:10.1111/tpj.v99.6URL [本文引用: 2]
MENGJ, ZHOUQ, ZHOUX, FANG HX, JI SJ. Ethylene and 1-MCP treatments affect leaf abscission and associated metabolism of Chinese cabbage Postharvest Biology and Technology, 2019, 157:110963. DOI:10.1016/j.postharvbio.2019.110963URL [本文引用: 3]
MEIRS, PHILOSOPH-HADASS, SUNDARESANS, SELVARAJK S V, BURDS, OPHIRR, KOCHANEKB, REIDM S, JIANGC Z, LERSA. Microarray analysis of the abscission-related transcriptome in the tomato flower abscission zone in response to auxin depletion Plant Physiology, 2010, 154(4):1929-1956. DOI:10.1104/pp.110.160697URL [本文引用: 1]
ZHAO YD. Essential roles of local auxin biosynthesis in plant development and in adaptation to environmental changes Annual Review of Plant Biology, 2018, 69:417-435. DOI:10.1146/arplant.2018.69.issue-1URL [本文引用: 1]
PARRA-LOBATO MC, GOMEZ-JIMENEZ MC. Polyamine- induced modulation of genes involved in ethylene biosynthesis and signalling pathways and nitric oxide production during olive mature fruit abscission Journal of Experimental Botany, 2011, 62(13):4447-4465. DOI:10.1093/jxb/err124URL [本文引用: 1]
BUNYA-ATICHARTK, KETSAS, VAN DOORNW. Ethylene- sensitive and ethylene-insensitive abscission in Dendrobium: Correlation with polygalacturonase activity Postharvest Biology and Technology, 2011, 60(1):71-74. DOI:10.1016/j.postharvbio.2010.11.006URL [本文引用: 1]
SINGH AP, PANDEY SP, RAJLUXMI, PANDEYS, NATHP, SANE AP. Transcriptional activation of a pectate lyase gene, RbPel1, during petal abscission in rose Postharvest Biology and Technology, 2011, 60(2):143-148. DOI:10.1016/j.postharvbio.2010.12.014URL [本文引用: 1]
WANGZ, SHI GA, MA XQ, FAN BY. The ethylene metabolism in flowers of Chinese peony ‘Taohua feixue’ during opening and senescence Acta Horticulturae Sinica, 2014, 41(11):2268-2274. (in Chinese) [本文引用: 1]
LIZADA MC, YANG SF. A simple and sensitive assay for 1-aminocyclopropane-1-carboxylic acid Analytical Biochemistry, 1979, 100(1):140-145. DOI:10.1016/0003-2697(79)90123-4URL [本文引用: 1]
FERNÁNDEZ-MACULET JC, YANG SF. Extraction and partial characterization of the ethylene-forming enzyme from apple fruit Plant Physiology, 1992, 99(2):751-754. DOI:10.1104/pp.99.2.751URL [本文引用: 1]
KATOM, HYODOH. Purification and characterization of ACC oxidase and increase in its activity during ripening of pear fruit Journal of the Japanese Society for Horticultural Science, 1999, 68(3):551-557. [本文引用: 1]
YANG JC, ZHANG JH, WANG ZQ, ZHU QS, WANGW. Hormonal changes in the grains of rice subjected to water stress during grain filling Plant Physiology, 2001, 127(1):315-323. DOI:10.1104/pp.127.1.315URL [本文引用: 1]
ZHANG ZL. Experimental Guidance of Plant Physiology. 2nd ed. Beijing: Higher Education Press, 1991. (in Chinese) [本文引用: 1]
LOHANIS, TRIVEDI PK, NATHP. Changes in activities of cell wall hydrolases during ethylene-induced ripening in banana: Effect of 1-MCP, ABA and IAA Postharvest Biology and Technology, 2004, 31(2):119-126. DOI:10.1016/j.postharvbio.2003.08.001URL [本文引用: 1]
CAO JK, JIANG WB, ZHAO YM. Physiological and Biochemical Experiment Guidance for Fruits and Vegetables After Harvest. Beijing: China Light Industry Press, 2007. (in Chinese) [本文引用: 2]
TIEMAN DM, CIARDI JA, TAYLOR MG, KLEE HJ. Members of the tomato LeEIL (EIN3-like) gene family are functionally redundant and regulate ethylene responses throughout plant development The Plant Journal, 2001, 26(1):47-58. DOI:10.1046/j.1365-313x.2001.01006.xURL [本文引用: 1]
KE MY, GAOZ, CHEN JQ, QIU YT, ZHANG LS, CHENX. Auxin controls circadian flower opening and closure in the waterlily BMC Plant Biology, 2018, 18(1):143. DOI:10.1186/s12870-018-1357-7URL [本文引用: 1]
REN XY, LIU GC, LI GP, YE CH, FENGF, WANG JN. Effects of ethephon and 1-MCP on the expression of AheAAT gene and AheERF transcription factors in jackfruit fruit Scientia Agricultura Sinica, 2019, 52(21):3890-3902. (in Chinese) [本文引用: 1]
WANG YC, LI TL, HOU JP. Effect of ethylene on abscission of tomato pedicel in vitro Acta Horticulturae Sinica, 2003, 30(5):554-558.(in Chinese) [本文引用: 1]
SONG LP, LIU JH, ZHENG DF, FENG NJ. Effects of different plant growth regulators on abscission rate and polygalacturonase activities of soybean flowers and pods by spraying in different stages Plant Physiology Communications, 2011, 47(4):356-362. (in Chinese) [本文引用: 1]
HU ZH, WANG YJ, CHEN CY. Effects of spraying cytokinin on abscission rate and enzymic activity of flowers and pods of cowpea Plant Science Journal, 2016, 34(3):439-445. (in Chinese) [本文引用: 1]
 ,1, 施江1, 高双成1, 王占营2, 史国安
,1, 施江1, 高双成1, 王占营2, 史国安

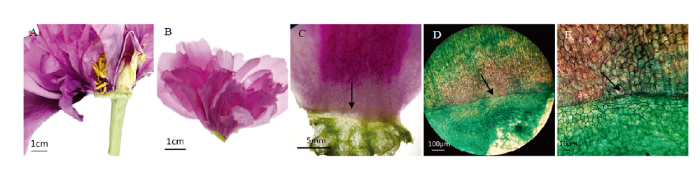 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT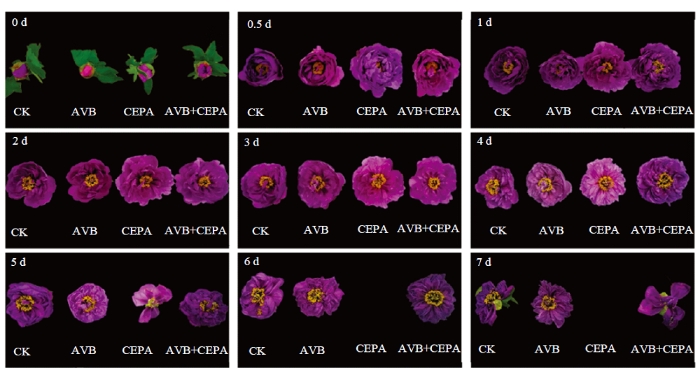 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT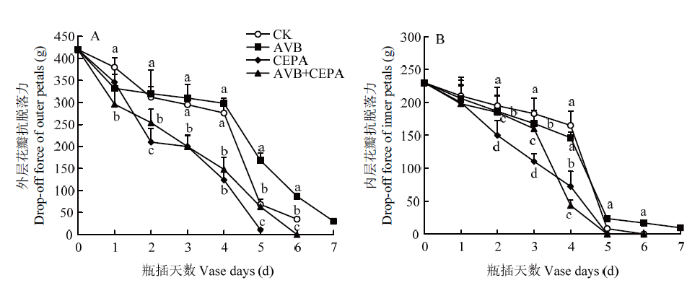 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT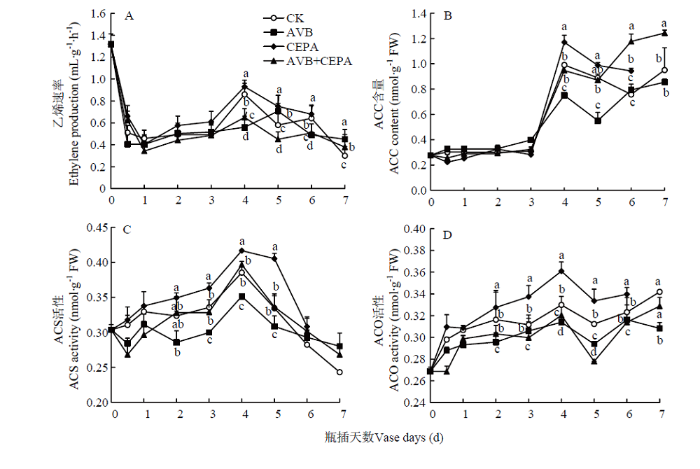 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT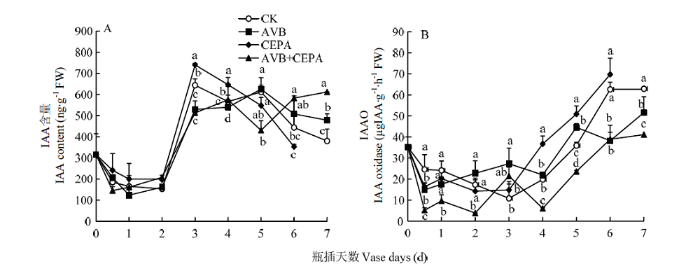 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT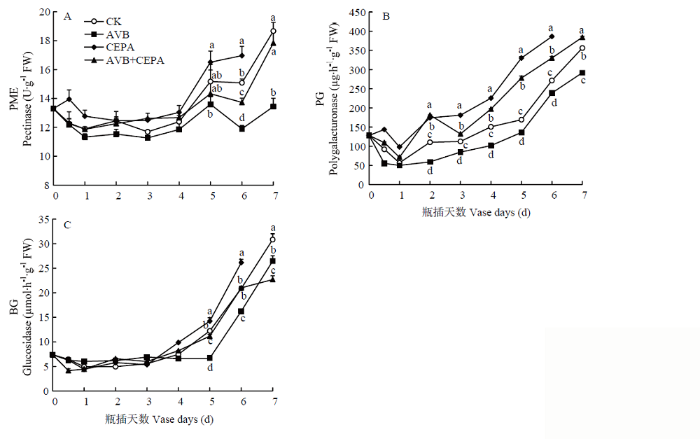 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT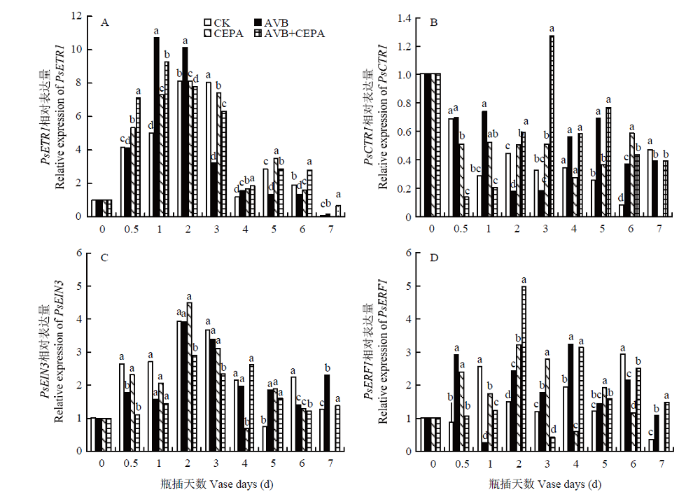 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT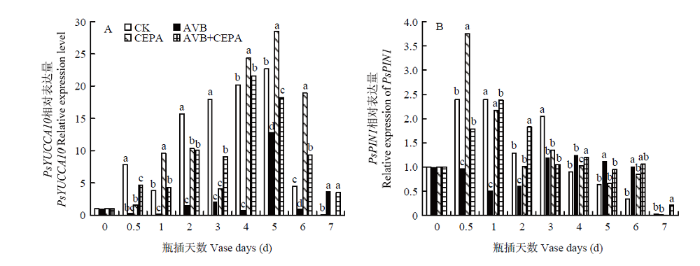 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT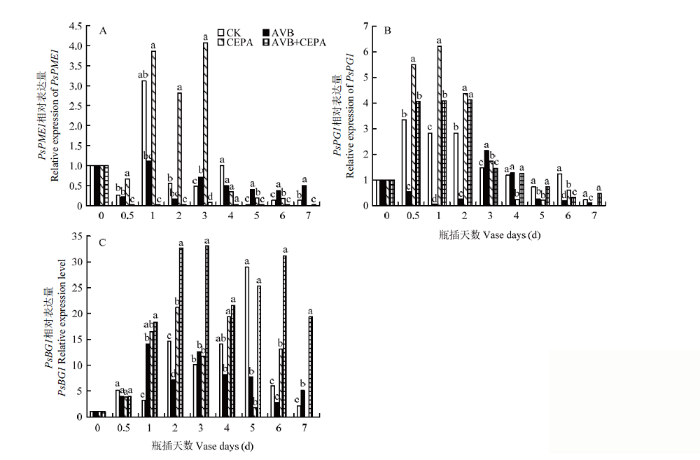 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}