Effects of miR-31-5p on the Proliferation and Apoptosis of Hair Follicle Stem Cells in Goat
FENG YunKui,, WANG Jian, MA JinLiang, ZHANG LiuMing, LI YongJun,College of Animal Science and Technology, Yangzhou University, Yangzhou 225009, Jiangsu
Abstract 【Objective】 The Yangtze River Delta White Goat is the only goat breed that can produce superior-quality brush hair in China and the world. The transcriptome sequencing results of the research team showed that there were significant differences in the expression level of MAP3K1 in the individual skin tissues of superior-quality brush hair and normal-quality brush hair. This study aimed to explore the key miRNAs that interacted with MAP3K1 during the formation of superior-quality brush hair and their effects on the proliferation and apoptosis of goat hair follicle stem cells. 【Method】 The bioinformatics websites (StrBase, miRDB, TargetScan, miRWalk, DAVID, KEGG, and RNAhybrid) were used to predict and select the miRNAs, with targeted relationship with MAP3K1, and use the online website Venny 2.1 to draw a Venn diagram. Through the construction of miR-31-5p overexpression vector, wild-MAP3K1, wild/Mut-RASA1 Luciferase Reporter assay vector, the relationship between miR-31-5p and MAP3K1, RASA1 was verified, and the effects of miR31-5p on MAP3K1, RASA1 mRNA and protein expression were detected by qPCR and Western Blot. In order to explore the effect of overexpression of miR-31-5p on cell proliferation and apoptosis, the mRNA and protein expression levels of proliferation marker gene (PCNA,CDK1,CCND2), anti-apoptotic gene (Bcl-2) and pro-apoptotic gene (Bax) in hair follicle stem cells transfected with miR-31-5p were detected, and the effects of overexpression of miR-31-5p on the viability, cell cycle and apoptosis of hair follicle stem cells were verified by CCK-8, EdU, flow cytometry. 【Result】 Through the database, the final score of the three miRNAs were predicted, which might relatively highly interact with MAP3K1. Then, combined with the known miRNAs studies in skin and hair follicle cells, miR-31-5p with highest score was selected as research object. After transfection of miR-31-5p, the relative expression of miR-31-5p in cells was detected, and it was found that the expression of miR-31-5p was significantly higher than that in the control group and blank vector group (P<0.01). The results of double luciferase reporter genes showed that overexpression of miR-31-5p could increase the activity of MAP3K1. Combined with Target Scan and KEGG database, it was predicted that miR-31-5p could target RASA1, the upstream inhibitor of MAP3K1 in MAPK signal pathway. In order to verify the relationship between miR-31-5p and RASA1, it was found that overexpression of miR31-5p inhibited the activity of RASA1 (P<0.01); qPCR and Western Blot assays showed that overexpression of miR-31-5p significantly inhibited the expression of mRNA and protein of RASA1 and promoted the expression of MAP3K1 (P<0.01). CCK-8 assays showed that overexpression of miR-31-5p increased the ability of cell proliferation. EdU staining showed that the rate of positive cells overexpressing miR-31-5p was significantly higher than that in the blank group (P<0.01), and promoted cell proliferation. Cell cycle data showed that after overexpression of miR-31-5p, the proportion of cells in G1/G0 phase was 52.23%, which was significantly lower than that in Control group (56.81% P<0.01). It slowed down the cell arrest in G1/G0 phase, but there was no significant difference between S phase and G2/M phase, however there was still an upward trend. Through apoptosis experiment, it was found that the survival rate of miR-31-5p group was 93.8%, and the total apoptosis rate was 4.9%; while that of control group was only 90.1%, and the total apoptosis rate was 8.41%, which indicated that the apoptosis rate decreased significantly after overexpression of miR-31-5p. Finally, the effects of miR-31-5p on proliferation and apoptosis-related genes were detected. It was found that overexpression of miR-31-5p significantly increased the mRNA and protein expression levels of proliferation marker genes and anti-apoptosis genes (Bcl-2), and decreased the mRNA and protein expression levels of pro-apoptosis gene (Bax). According to the results of the research, the molecular mechanism of miR-31-5p in hair follicle stem cells was revealed. 【Conclusion】 miR-31-5p targeted RASA1 and up-regulated the expression level of MAP3K1, thereby promoting the proliferation of hair follicle stem cells and inhibiting their apoptosis. It provided a theoretical basis for further investigating the molecular mechanism that regulates the characteristics of superior-quality brush hair of the Yangtze River Delta White Goats. Keywords:Yangtze River Delta White Goat;superior-quality brush hair trait;hair follicle stem cells;MAP3K1;miR-31-5p;RASA1;proliferation;apoptosis
PDF (1939KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 冯云奎, 王健, 马金亮, 张柳明, 李拥军. miR-31-5p对山羊毛囊干细胞增殖和凋亡的影响. 中国农业科学, 2021, 54(23): 5132-5143 doi:10.3864/j.issn.0578-1752.2021.23.017 FENG YunKui, WANG Jian, MA JinLiang, ZHANG LiuMing, LI YongJun. Effects of miR-31-5p on the Proliferation and Apoptosis of Hair Follicle Stem Cells in Goat. Scientia Acricultura Sinica, 2021, 54(23): 5132-5143 doi:10.3864/j.issn.0578-1752.2021.23.017
开放科学(资源服务)标识码(OSID):
0 引言
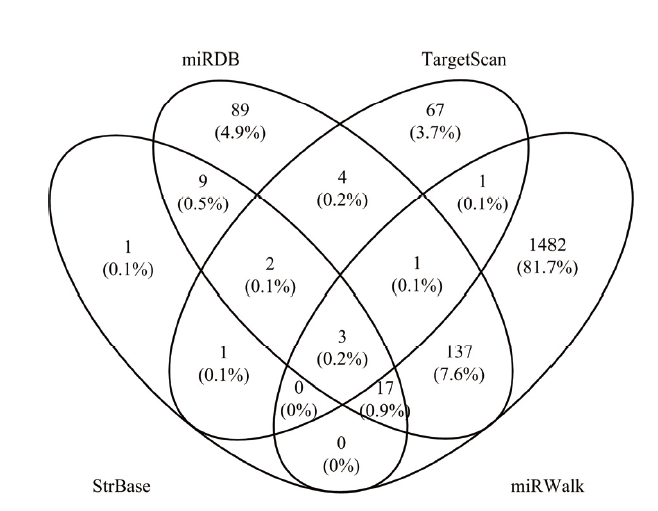
【研究意义】长江三角洲白山羊,是国内外唯一能生产优质笔料毛的山羊品种,其颈脊部产出的毛发具有“毛色洁白、挺直有峰、弹性好”等优良性状,为制作高档毛笔的独特原料[1,2]。【前人研究进展】毛发生长与毛囊发育之间存在着密切联系,毛囊生长发育不仅仅影响着绒毛的生长,而且也影响着绒毛的质量[3]。毛囊发育为一个循环的生物系统,包括生长期,退行期和休止期三个阶段,并涉及基因调控的动态变化[4]。课题组前期转录组测序筛选出促分裂原活化蛋白激酶的激酶1(mitogen-activated protein kinase kinase kinase 1, MAP3K1),它是与优质笔料毛性状相关的候选基因之一,在优质笔料毛中RPKM(reads per kilobase per million reads)值为8.7731,在非优质笔料毛个体中RPKM值仅为0.0345,log2Fold Change值为7.49[5]。MAP3K1,又称MEKK1,是MAPK信号通路中的重要调控因子。已有的研究发现,MAP3K1除了参与免疫机制,损伤修复,肿瘤进展和骨骼肌生长发育的调控外,还在毛发形成方面起着至关重要的作用[6]。microRNA是一种短链(20—24 nt)非编码RNA,通过与靶基因的mRNA 3′UTR结合[7,8]进而降解或抑制靶基因表达[9]。miRNAs参与多种生理进程,包括组织发育、器官形成、细胞生长和凋亡等过程[10,11,12]。迄今为止,在毛囊发育过程中鉴定出许多miRNAs,如miR-203[13]、miR-let-7a[14]和miR-196a[15]等对于调节毛囊的发育和再生至关重要。已有的研究指出,miR- 31-5p是与肿瘤相关的miRNA,miR-31-5p在大肠癌、口腔癌、结肠腺癌中通过抑制靶基因的转录或翻译,促进癌细胞的生长、迁移和侵袭等过程[7, 16-17],也有相关文献证实miR-31参与皮肤组织修复,如系统性红斑狼疮、伤口愈合等[18]。最近研究表明,miR-31-5p可与RAS p21蛋白激活剂1(RAS p21 protein activator 1, RASA1)的3′UTR结合,RASA1是RAS / MAPK信号通路的负调节因子,然后激活RAS / MAPK(小鼠皮肤组织中的ERK1 / 2)通路,促进细胞存活[19]。【本研究切入点】关于miR-31-5p在长江三角洲白山羊优质笔料毛形成过程中的作用和调控机制以及在毛囊干细胞中的功能知之甚少。【拟解决的关键问题】本研究通过验证miR-31-5p是否直接靶向RASA1影响MAP3K1,进而激活MAPK信号通路,然后调节长江三角白山羊优质笔料毛的形成,为长江三角洲白山羊的分子选育提供理论依据。
A:miR-31-5p过表达效率;B:MAP3K1双荧光结果;C:RASA1双荧光结果;D:过表达miR-31-5p对MAP3K1和RASA1基因mRNA的影响;E、F:过表达miR-31-5p对MAP3K1和RASA1基因蛋白的影响 Fig. 2The effects of overexpression of miR-31-5p on MAP3K1 and RASA1
A: Overexpression efficiency of miR-31-5p; B: The result of Double-Luciferase Reporter of MAP3K1; C: The result of Double-Luciferase Reporter of RASA1; D: Effects of MAP3K1 and RASA1 mRNA after overexpression of miR-31-5p; E, F: Effects of MAP3K1 and RASA1 protein after overexpression of miR-31-5p
A:CCK-8结果;B:EdU试验染色结果;C:EdU阳性细胞率;D:Control组周期分布图;E:pre-miR-31-5p组周期分布图;F:Control组和pre-miR-31-5p组 G0/G1、S、G2/M期细胞数量对比图 Fig. 3The effects of overexpression of miR-31-5p on the proliferation and cell cycle of hair follicle stem cells
A: Result of CCK-8结果; B: Result of EdU assay; C: Percentage of EdU-positive Cells; D: Periodic distribution diagram of Control; E: Periodic distribution diagram of pre-miR-31-5p; F: Comparison of cell number in G0/G1, S and G2/M phase between the Control and the pre-miR-31-5p
A:Control组细胞凋亡分布图;B:pre-miR-31-5p组细胞凋亡分布图;C:Control组和pre-miR-31-5p组细胞凋亡率对比图 Fig. 4The effect of overexpression of miR-31-5p on the apoptosis of hair follicle stem cells
A: Apoptosis profile of Control; B: Apoptosis profile of pre-miR-31-5p; C: Comparison of apoptosis rate between the Control and the pre-miR-31-5p
A:过表达miR-31-5p对增殖和凋亡相关基因mRNA的影响;B:过表达miR-31-5p对增殖和凋亡相关基因蛋白的影响 Fig. 5The effect of overexpression of miR-31-5p on proliferation and apoptosis related genes
A: Effect of overexpression of miR-31-5p on the mRNA of proliferation and apoptosis related genes; B: Effect of overexpression of miR-31-5p on the proteins of proliferation and apoptosis related genes
LI YJ, HUANG YH. The status of the goat and its wool production for writing brush in China China Herbivores, 2005, 25(1):44-45. doi: 10.3969/j.issn.2095-3887.2005.01.026. (in Chinese) [本文引用: 1]
GUOH, CHENGG, LIY, ZHANGH, QINK. A screen for key genes and pathways involved in high-quality brush hair in the Yangtze River Delta white goat PLoS ONE, 2017, 12(1):e0169820. doi: 10.1371/journal.pone.0169820. URL [本文引用: 1]
MENGY, JIANG HZ. Research on the development of skin and hair follicles of Liaoning cashmere goats at lamb stage China Herbivore Science, 2020(5):70-73. (in Chinese) [本文引用: 1]
MARDARYEV AN, AHMED MI, VLAHOV NV, FESSING MY, GILL JH, SHAROV AA, BOTCHKAREVA NV. Micro-RNA-31 controls hair cycle-associated changes in gene expression programs of the skin and hair follicle BMC Dermatology, 2010, 24(10):3869-3881. doi: 10.1096/fj.10-160663. [本文引用: 1]
JID, YANGB, LIY, CAIM, ZHANGW, CHENGG, GUOH. Transcriptomic inspection revealed a possible pathway regulating the formation of the high-quality brush hair in Chinese Haimen goat (Capra hircus) Royal Society Open Science, 2018, 5(1):170907. doi: 10.1098/rsos.170907. URL [本文引用: 1]
ECKERT RL, EFIMOVAT, DASHTI SR, BALASUBRAMANIANS, DEUCHERA, CRISH JF, STURNIOLOM, BONEF. Keratinocyte survival, differentiation, and death: Many roads lead to mitogen- activated protein kinase The Journal of Investigative Dermatology Symposium Proceedings, 2002, 7(1):36-40. doi: 10.1046/j.1523-1747.2002.19634.x. [本文引用: 1]
PENGH, WANGL, SUQ, YIK, DUJ, WANGZ. miR-31-5p promotes the cell growth, migration and invasion of colorectal cancer cells by targeting NUMB Biomedicine & Pharmacotherapy, 2019, 109:208-216. doi: 10.1016/j.biopha.2018.10.048. URL [本文引用: 2]
LEWIS BP, BURGE CB, BARTEL DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets Cell, 2005, 120(1):15-20. doi: 10.1016/j.cell.2004.12.035. URL [本文引用: 1]
YIR, O'CARROLLD, PASOLLIH A, ZHANGZ, DIETRICHF S, TARAKHOVSKYA, FUCHSE. Morphogenesis in skin is governed by discrete sets of differentially expressed microRNAs Nature Genetics, 2006, 38(3):356-362. doi: 10.1038/ng1744. URL [本文引用: 1]
YIR, POY MN, STOFFELM, FUCHSE. A skin microRNA promotes differentiation by repressing ‘stemness’ Nature, 2008, 452(7184):225-229. DOI:10.1038/nature06642URL [本文引用: 1]
PAL AS, KASINSKI AL. Animal models to study microRNA function Advances in Cancer Research, 2017, 135:53-118. doi: 10.1016/bs.acr.2017.06.006. [本文引用: 1]
LENA AM, SHALOM-FEUERSTEINR, RIVETTIDI VAL CERVO P, ABERDAMD, KNIGHTR A, MELINOG, CANDIE. miR-203 represses ‘stemness’ by repressing DeltaNp63 Cell Death and Differentiation, 2008, 15(7):1187-1195. doi: 10.1038/cdd.2008.69. URL [本文引用: 1]
MAT, LI JP, JIANGQ, WU SF, JIANG HZ, ZHANG QL. Differential expression of miR-let7a in hair follicle cycle of Liaoning cashmere goats and identification of its targets Functional & Integrative Genomics, 2018, 18(6):701-707. doi: 10.1007/s10142-018-0616-x. [本文引用: 1]
WANGZ, JINNINM, KUDOH, INOUEK, NAKAYAMAW, HONDAN, MAKINOK, KAJIHARAI, FUKUSHIMAS, INOUEY, IHNH. Detection of hair-microRNAs as the novel potent biomarker: evaluation of the usefulness for the diagnosis of Scleroderma Journal of Dermatological Science, 2013, 72(2):134-141. doi: 10.1016/j.jdermsci.2013.06.018. URL [本文引用: 1]
KAO YY, CHOU CH, YEH LY, CHEN YF, CHANG KW, LIU CJ, FANCHIANG C Y, LINS C. microRNA miR-31 targets SIRT3 to disrupt mitochondrial activity and increase oxidative stress in oral carcinoma Cancer Letters, 2019, 456:40-48. doi: 10.1016/j.canlet.2019.04.028. URL [本文引用: 1]
ZHANGY, MA DD, ZHANG ZL, ZHANGY, CAOJ. The effect and mechanism of miR-31 on the metastasis and invasion of colon cancer Journal of Clinical Surgery, 2018(10):747-750. (in Chinese) [本文引用: 1]
SHI JY. MiR-31 mediates inflammatory signaling to promote re-epithelialization during skin wound healing [D]. Beijing: China Agricultural University, 2018. (in Chinese) [本文引用: 3]
SHIJ, MAX, SUY, SONGY, TIANY, YUANS, ZHANGX, YANGD, ZHANGH, SHUAIJ, CUIW, RENF, PLIKUS MV, CHENY, LUOJ, YUZ. miR-31 mediates inflammatory signaling to promote Re-epithelialization during skin wound healing The Journal of Investigative Dermatology, 2018, 138(10):2253-2263. doi: 10.1016/j.jid.2018.03.1521. URL [本文引用: 1]
WANGQ, QUJ, LIY, JID, ZHANGH, YINX, WANGJ, NIUH. Hair follicle stem cells isolated from newborn Yangtze River Delta White Goats Gene, 2019, 698:19-26. doi: 10.1016/j.gene.2019.02.052. URL [本文引用: 1]
DWEEPH, GRETZN. miRWalk2.0: A comprehensive atlas of microRNA-target interactions Nature Methods, 2015, 12(8):697. doi: 10.1038/nmeth.3485. URL [本文引用: 1]
ZHANG LH, MA YY, LIU WY, LAN WT, ZHUZ W. microRNA- 96-5p targets MITF gene in alpaca melanocytes Acta Veterinaria et Zootechnica Sinica, 2020, 51(6):1229-1237. doi: 10.11843/j.issn.0366-6964.2020.06.007. (in Chinese) [本文引用: 1]
SLATTERY ML, LUNDGREENA, WOLFF RK. MAP kinase genes and colon and rectal cancer Carcinogenesis, 2012, 33(12):2398-2408. DOI:10.1093/carcin/bgs305URL [本文引用: 1]
KLINGE CM, BLANKENSHIP KA, RISINGER KE, BHATNAGARS, NOISIN EL, SUMANASEKERA WK, ZHAOL, BREY DM, KEYNTON RS. Resveratrol and estradiol rapidly activate MAPK signaling through estrogen receptors alpha and beta in endothelial cells The Journal of Biological Chemistry, 2005, 280(9):7460-7468. doi: 10.1074/jbc.m411565200. URL [本文引用: 1]
REBBECK TR, DEMICHELEA, TRAN TV, PANOSSIANS, BUNIN GR, TROXEL AB, STROM BL. Hormone-dependent effects of FGFR2 and MAP3K1 in breast cancer susceptibility in a population-based sample of post-menopausal African-American and European-American women Carcinogenesis, 2009, 30(2):269-274. doi: 10.1093/carcin/bgn247. URL [本文引用: 1]
YANGB. Study on high quality brush hair gens based on RNA-Seq of Yangtze River Delta White Goat and observation on follicles structures [D]. Yangzhou: Yangzhou University, 2015. (in Chinese) [本文引用: 1]
CORREIA DE SOUSAM, GJORGJIEVAM, DOLICKAD, SOBOLEWSKIC, FOTIM. Deciphering miRNAs’ action through miRNA editing International Journal of Molecular Sciences, 2019, 20(24):6249. DOI:10.3390/ijms20246249URL [本文引用: 1]
WANGJ, WANGW, LIJ, WUL, SONGM, MENGQ. miR182 activates the Ras-MEK-ERK pathway in human oral cavity squamous cell carcinoma by suppressing RASA1 and SPRED1 OncoTargets and Therapy, 2017, 10:667-679. doi: 10.2147/ott.s121864. URL [本文引用: 1]
STEPICHEVA NA, SONG JL. Function and regulation of microRNA-31 in development and disease Molecular Reproduction and Development, 2016, 83(8):654-674. doi: 10.1002/mrd.22678. URL [本文引用: 1]
HEJ, JINS, ZHANGW, WUD, LIJ, XUJ, GAOW. Long non-coding RNA LOC554202 promotes acquired gefitinib resistance in non-small cell lung cancer through upregulating miR-31 expression Journal of Cancer, 2019, 10(24):6003-6013. doi: 10.7150/jca.35097. URL [本文引用: 1]
HUC, HUANGF, DENGG, NIEW, HUANGW, ZENGX. miR-31 promotes oncogenesis in intrahepatic cholangiocarcinoma cells via the direct suppression of RASA1 Experimental and Therapeutic Medicine, 2013, 6(5):1265-1270. doi: 10.3892/etm.2013.1311. URL [本文引用: 1]
ZHU YQ, WANGH, SHI XY, WANG JJ, WANG WG, XUEL X. miR-31 promotes the proliferation of colorectal cancer cells through activating NF-B signal pathway Chinese Journal of Biochemistry and Molecular Biology, 2017, 33(9):908-916.(in Chinese) [本文引用: 1]
MA JL, WANGJ, FENG YK, WANGQ, ZHANG LM, LI YJ. Effect of interference with MAP3K1 gene on proliferation and apoptosis of goat hair follicle stem cells Journal of Northeast Agricultural University, 2020(10):56-62. (in Chinese) [本文引用: 1]
 ,, 王健, 马金亮, 张柳明, 李拥军
,, 王健, 马金亮, 张柳明, 李拥军

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT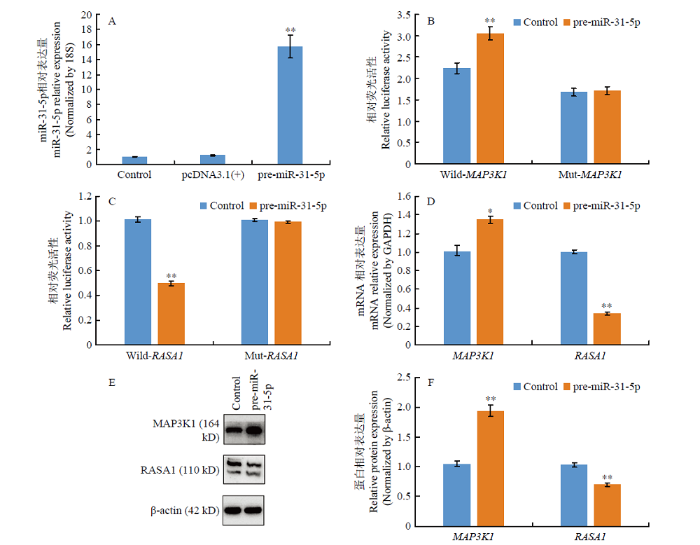 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT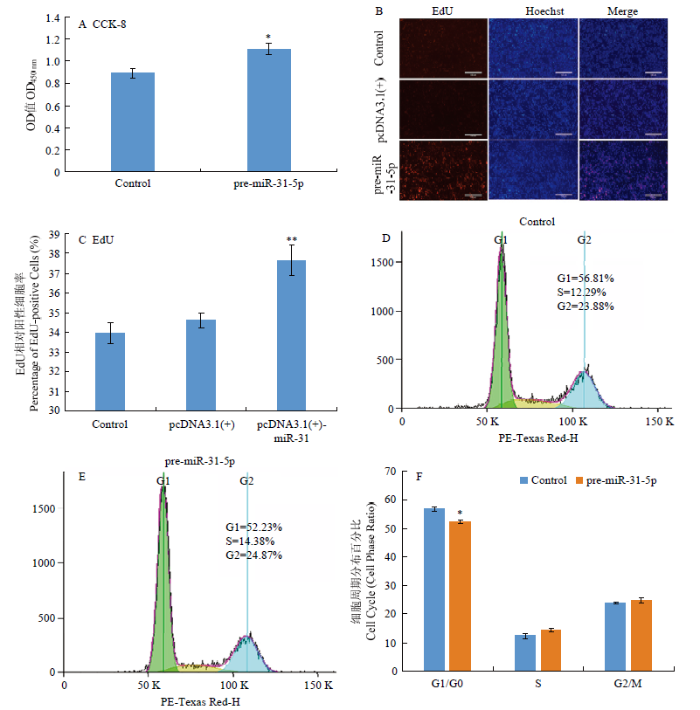 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT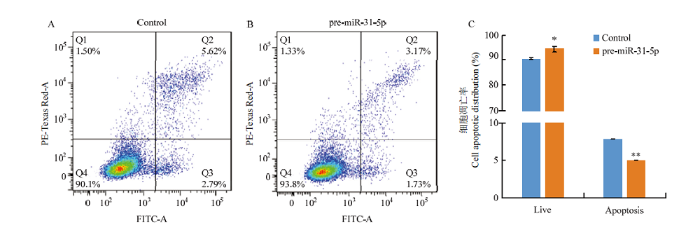 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT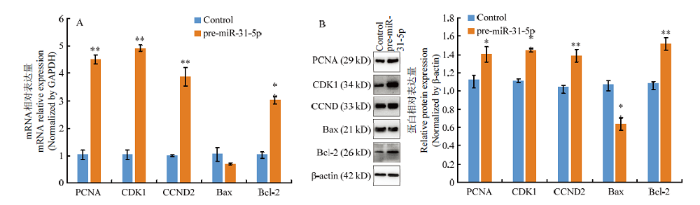 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT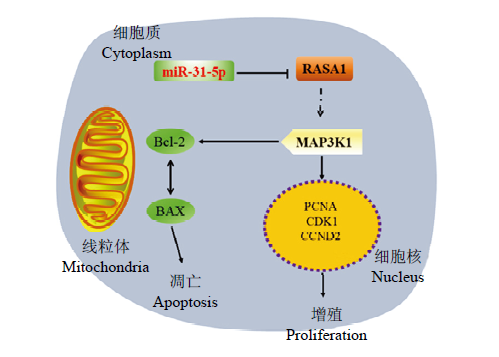 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}