Effect of Bacillus subtilis NCD-2 on the Growth of Tomato and the Microbial Community Structure of Rhizosphere Soil Under Salt Stress
SHAO MeiQi,1,2, ZHAO WeiSong2, SU ZhenHe2, DONG LiHong2, GUO QingGang,2,*, MA Ping21College of Plant Protection, Hebei Agricultural University, Baoding 071001, Hebei 2Plant Protection Institute of Hebei Academy of Agricultural and Forestry Sciences/IPM Centre of Hebei Province/Key Laboratory of IPM on Crops in Northern Region of North China, Ministry of Agriculture and Rural Affairs, Baoding 071000, Hebei
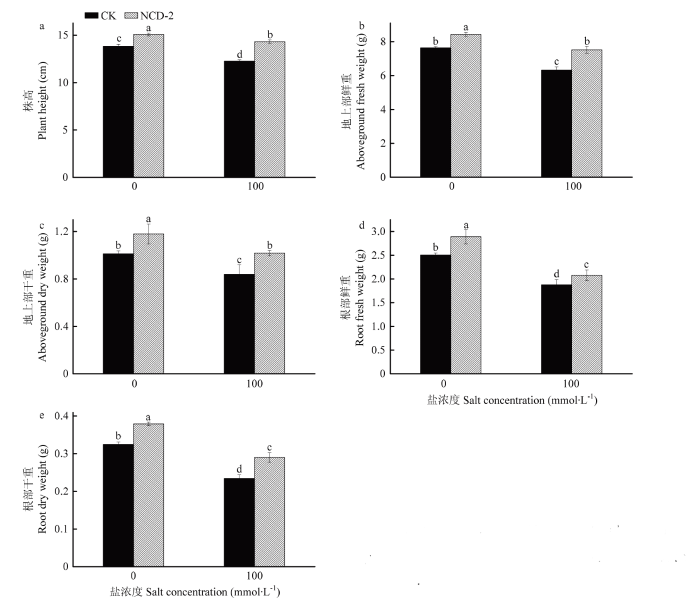
Abstract 【Objective】The objective of this study is to evaluate the growth promotion effect of Bacillus subtilis strain NCD-2 on tomato seedlings under salt stress, as well as the effect of strain NCD-2 on soil microbial community diversity. The results will be useful for expanding the application of strain NCD-2 in agricultural system.【Method】The pot experiments were conducted to evaluate the effects of strain NCD-2 treatment on stem length, aboveground fresh weight, aboveground dry weight, root fresh weight and root dry weight. The activity of resistance-related enzymes such as peroxidase (POD), superoxide dismutase (SOD), catalase (CAT) and the content of abscisic acid (ABA) were measured. High-throughput sequencing (Illumina MiSeq) technique was used to determine the bacterial and fungal community structures in the rhizosphere soil. The four treatments were set as NCD-2 strain suspension treatment (NCD0), 100 mmol·L -1 NaCl treatment (CK100), NCD-2 strain suspension + 100 mmol·L-1 NaCl treatment (NCD100) and water treatment as control (CK0).【Result】Under normal conditions, strain NCD-2 treatment significantly increased the biomass of tomato. The plant height, aboveground fresh weight, aboveground dry weight, root fresh weight and root dry weight were increased by 9.08%, 10.37%, 16.64%, 15.42% and 16.78%, respectively, when compared with the control. Under salt stress, compared to the control, the plant height, aboveground fresh weight, aboveground dry weight, root fresh weight and root dry weight were increased by 16.86%, 18.96%, 21.32%, 10.50% and 23.99% after treated with strain NCD-2, respectively. The activity of resistance-related enzymes SOD, POD, CAT and the content of ABA were increased by 50.45%, 56.18%, 29.55% and 34.60% after treated with strain NCD-2, respectively, when compared with the control. For bacteria community composition analysis, compared to CK0, the relative abundance of bacteria phylum Actinobacteria, Acidobacteria and Chloroflexi was increased by 7.28%, 15.14% and 23.03% after treated with strain NCD-2 without salt stress, respectively. The relative abundance of bacteria genus Arthrobacter, Sphingomonas, Microvirga and Streptomyces was increased by 50.88%, 15.31%, 11.32% and 16.41% after treated with strain NCD-2, respectively. Under 100 mmol·L -1 NaCl stress, compared to CK100, the relative abundance of bacteria phylum Proteobacteria, Actinobacteria, Firmicutes and Gemmatimonadetes was increased by 6.08%, 8.19%, 14.11% and 4.70% after treated with strain NCD-2, respectively. The relative abundance of bacteria genus Arthrobacter, Bacillus, Sphingomonas and Microvirga was increased by 5.54%, 31.80%, 23.39% and 23.08% after treated with strain NCD-2, respectively. For fungal community composition analysis, compared to CK0, the relative abundance of fungal phylum Mortierellomycota, Glomeromycota and Chytridiomycota was increased to 186%, 477% and 1 650% of CK0, respectively. The relative abundance of fungal genus Mortierella, Trichoderma and Preussia was increased to 186%, 108%, and 120% of CK0 after treated with strain NCD-2 without salt stress, respectively. Under 100 mmol·L -1 NaCl stress, compared to CK100, the relative abundance of fungal phylum Mortierellomycota, Glomeromycota and Chytridiomycota was increased to 345%, 154%, 921% of CK100 after treated with strain NCD-2, respectively. The relative abundance of fungal genus Mortierella was increased by 246% after treated with strain NCD-2.【Conclusion】After the treatment of strain NCD-2 under salt stress, the activity of stress-resistant enzymes and the content of ABA in tomato were increased, and the population of beneficial microorganisms in tomato rhizosphere was increased, thus improving the tolerance of tomato to salt stress and significantly increasing the growth and development of tomato. Keywords:Bacillus subtilis;salt stress;activity of antioxidant enzymes;high-throughput sequencing;soil microbial community structure
PDF (1124KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 邵美琪, 赵卫松, 苏振贺, 董丽红, 郭庆港, 马平. 盐胁迫下枯草芽孢杆菌NCD-2对番茄促生作用及对土壤微生物群落结构的影响. 中国农业科学, 2021, 54(21): 4573-4584 doi:10.3864/j.issn.0578-1752.2021.21.008 SHAO MeiQi, ZHAO WeiSong, SU ZhenHe, DONG LiHong, GUO QingGang, MA Ping. Effect of Bacillus subtilis NCD-2 on the Growth of Tomato and the Microbial Community Structure of Rhizosphere Soil Under Salt Stress. Scientia Agricultura Sinica, 2021, 54(21): 4573-4584 doi:10.3864/j.issn.0578-1752.2021.21.008
a:门水平细菌群落结构PCOA分析 PCOA analysis of bacterial community structure at phylum level;b:属水平细菌群落结构PCOA分析 PCOA analysis of bacterial community structure at genus level Fig. 3PCOA analysis of bacterial community structure
a:门水平真菌群落结构PCOA分析PCOA analysis of fungus community structure at phylum level;b:属水平真菌群落结构PCOA分析PCOA analysis of fungus community structure at genus level Fig. 6PCOA analysis of fungus community structure
MANGAT PK, GANNABAN RB, SINGLETON JJ, ANGELES- SHIM R B. Development of a PCR-based, genetic marker resource for the tomato-like nightshade relative, Solanum lycopersicoides using whole genome sequence analysis PLoS ONE, 2020, 15(11):e0242882. DOI:10.1371/journal.pone.0242882URL [本文引用: 1]
MAYAKS, TIROSHT, GLICK BR. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress Plant Physiology and Biochemistry, 2004, 42(6):565-572. DOI:10.1016/j.plaphy.2004.05.009URL [本文引用: 1]
GAMALEROE, BERTAG, MASSAN, GLICK BR, LINGUAG. Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and their consequences for the growth of cucumber under salt -stress conditions Journal of Applied Microbiology, 2010, 108(1):236-245. DOI:10.1111/jam.2009.108.issue-1URL [本文引用: 1]
NUMANM, BASHIRS, KHANY, MUMTAZR, SHINWARI ZK, KHAN AL, KHANA, AL-HARRASIA. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review Microbiological Research, 2018, 209:21-32. DOI:10.1016/j.micres.2018.02.003URL [本文引用: 1]
SHARMAS, KULKARNIJ, JHAB. Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut Frontiers in Microbiology, 2016, 7:1600. [本文引用: 1]
PAULD, LADEH. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review Agronomy for Sustainable Development, 2014, 34(4):737-752. DOI:10.1007/s13593-014-0233-6URL [本文引用: 1]
SARKARA, GHOSH PK, PRAMANIKK, MITRAS, SORENT, PANDEYS, MONDAL MH, MAITI TK. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress Research in Microbiology, 2018, 169(1):20-32. DOI:10.1016/j.resmic.2017.08.005URL [本文引用: 1]
NIU SQ. Physiological research on growth promoting rhizobacteria from Haloxylon ammodendron regulating perennial ryegrass growth and stress tolerance [D]. Lanzhou: Lanzhou University, 2017. (in Chinese) [本文引用: 1]
EL-ESAWIM, ALARAIDHI, ALSAHLIA, ALZAHRANIS, ALIH, ALAYAFIA, AHMADM. Serratia liquefaciens KM4 improves salt stress tolerance in maize by regulating redox potential, ion homeostasis, leaf gas exchange and stress-related gene expression International Journal of Molecular Sciences, 2018, 19(11):3310. DOI:10.3390/ijms19113310URL [本文引用: 1]
ZHOUC, MAZ, ZHUL, XIAOX, XIEY, ZHUJ, WANGJ. Rhizobacterial strain Bacillus megaterium BOFC15 induces cellular polyamine changes that improve plant growth and drought resistance International Journal of Molecular Sciences, 2016, 17(6):976. DOI:10.3390/ijms17060976URL [本文引用: 1]
YASMEENT, AHMADA, ARIF MS, MUBINM, REHMANK, SHAHZAD SM, IQBALS, RIZWANM, ALIS, ALYEMENI MN, WIJAYAL. Biofilm forming rhizobacteria enhance growth and salt tolerance in sunflower plants by stimulating antioxidant enzymes activity Plant Physiology and Biochemistry, 2020, 156:242-256. DOI:10.1016/j.plaphy.2020.09.016URL [本文引用: 2]
LATEF AA, OMER AM, BADAWY AA, OSMAN MS, RAGAEY MM. Strategy of salt tolerance and interactive impact of Azotobacter chroococcum and/or Alcaligenes faecalis inoculation on canola (Brassica napus L.) plants grown in saline soil Plants, 2021, 10(1):110. DOI:10.3390/plants10010110URL [本文引用: 1]
ZHAO DD. Effects of different concentrations of microbial agents on physical and chemical properties and growth of tomato soil [D]. Yangling: Northwest A&F University, 2020. (in Chinese) [本文引用: 1]
BUDDRUS-SCHIEMANNK, SCHMIDM, SCHREINERK, WELZLG, HARTMANNA. Root colonization by Pseudomonas sp. DSMZ 13134 and impact on the indigenous rhizosphere bacterial community of barley Microbial Ecology, 2010, 60(2):381-393. DOI:10.1007/s00248-010-9720-8URL [本文引用: 1]
GAMALEROE, LINGUAG, TOMBOLINIR, AVIDANOL, PIVATOB, BERTAG. Colonization of tomato root seedling by Pseudomonas fluorescens 92rkG5: Spatio-temporal dynamics, localization, organization, viability, and culturability Microbial Ecology, 2005, 50(2):289-297. DOI:10.1007/s00248-004-0149-9URL [本文引用: 1]
WANG XH, ZHANG GY, LIR, LU YL, RANW, SHEN QR. Control of watermelon fusarium wilt by using antagonist-enhanced biological organic fertilizers Journal of Plant Nutrition and Fertilizer, 2013, 19(1):223-231. (in Chinese) [本文引用: 1]
HUANG YL, ZHENG LW, HUANG YY, JIA ZH, SONG SS, LI ZX. Effects of different application methods of Bacillus subtilis agent on soil microbial diversity and growth of muskmelon Chinese Journal of Biotechnology, 2020, 36(12):2644-2656. (in Chinese) [本文引用: 1]
QIUQ, ZHANGL, HANG, SHIJ, HU ZF. Improving fertility of newly reclaimed soil by PGPR inoculums in combination with alfalfa growing Journal of Southwest University (Natural Science Edition), 2011, 33(5):109-115. (in Chinese) [本文引用: 1]
GIANNOPOLITIS CN, RIES SK. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings Plant Physiology, 1977, 59(2):315-318. DOI:10.1104/pp.59.2.315URL [本文引用: 1]
ZENG SX, WANG YR. Comparison of the changes of membrane protection system in rice seedlings during enhancement of chilling resistance by different stress pretreatments Acta Botanica Sinica, 1997, 39(4):308-314. (in Chinese) [本文引用: 1]
DHINDSA RS, PLUMB-DHINDSAP, THORPE TA. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase Journal of Experimental Botany, 1981, 32(126):93-101. DOI:10.1093/jxb/32.1.93URL [本文引用: 1]
ARKHIPOVA TN, PRINSENE, VESELOV SU, MARTINENKO EV, MELENTIEV AI, KUDOYAROVA GR. Cytokinin producing bacteria enhance plant growth in drying soil Plant and Soil, 2007, 292:305-315. DOI:10.1007/s11104-007-9233-5URL [本文引用: 1]
XUN, TANG, WANGH, GAIX. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure European Journal of Soil Biology, 2016, 74:1-8. DOI:10.1016/j.ejsobi.2016.02.004URL [本文引用: 1]
ADAMS RI, MILETTOM, TAYLOR JW, BRUNS TD. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances The ISME Journal, 2013, 7(7):1262-1273. DOI:10.1038/ismej.2013.28URL [本文引用: 2]
MELONI DA, OLIVA MA, MARTINEZ CA, CAMBRAIAJ. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress Environmental and Experimental Botany, 2003, 49(1):69-76. DOI:10.1016/S0098-8472(02)00058-8URL [本文引用: 1]
KIMK, JANG YJ, LEE SM, OH BT, CHAE JC, LEE KJ. Alleviation of salt stress by Enterobacter sp. EJ01 in tomato and Arabidopsis is accompanied by up-regulation of conserved salinity responsive factors in plants Molecules and Cells, 2014, 37(2):109-117. DOI:10.14348/molcells.2014.2239URL [本文引用: 1]
SAMADDARS, CHATTERJEEP, CHOUDHURY AR, AHMEDS, SAT. Interactions between Pseudomonas spp. and their role in improving the red pepper plant growth under salinity stress Microbiological Research, 2019, 219:66-73. DOI:10.1016/j.micres.2018.11.005URL [本文引用: 1]
KHAN MH, PANDA SK. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress Acta Physiologiae Plantarum, 2008, 30:81-89. DOI:10.1007/s11738-007-0093-7URL [本文引用: 1]
BHARTIN, PANDEY SS, BARNAWALD, PATEL VK, KALRAA. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress Scientific Reports, 2016, 6:34768. DOI:10.1038/srep34768URL [本文引用: 1]
MAUCH-MANIB, FLORSV. The ATAF1 transcription factor: At the convergence point of ABA-dependent plant defense against biotic and abiotic stresses Cell Research, 2009, 19(12):1322-1323. DOI:10.1038/cr.2009.135URL [本文引用: 1]
TAOZ, KOUY, LIUH, LIX, XIAOJ, WANGS. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice Journal of Experimental Botany, 2011, 62(14):4863-4874. DOI:10.1093/jxb/err144URL [本文引用: 1]
ZHENGN, KE LF, YANG JY, WANG XF, HUANGD, CHENG WL, LI JH, ZHENG LY, YU ZN, ZHANG JB. Growth improvement and salt tolerance mechanisms of tomato seedlings mediated by plant growth-promoting rhizobacteria from contaminated soils Chinese Journal of Applied and Environmental Biology, 2018, 24(1):47-52. (in Chinese) [本文引用: 1]
HE ZQ, HE CX, ZHANG ZB, ZOU ZR, WANG HS. Changes of antioxidative enzymes and cell membrane osmosis in tomato colonized by arbuscular mycorrhizae under NaCl stress Colloids and Surfaces B: Biointerfaces, 2007, 59(2):128-133. DOI:10.1016/j.colsurfb.2007.04.023URL [本文引用: 2]
BAKKER MG, CHAPARRO JM, MANTER DK, VIVANCO JM. Impacts of bulk soil microbial community structure on rhizosphere microbiomes of Zea mays Plant and Soil, 2015, 392:115-126. DOI:10.1007/s11104-015-2446-0URL [本文引用: 1]
WALTERS WA, JINZ, YOUNGBLUTN, WALLACE JG, SUTTERJ, ZHANGW, GONZALEZ-PENAA, PEIFFERJ, KORENO, SHIQ, et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(28):7368-7373. [本文引用: 1]
BERENDSEN RL, PIETERSEC, BAKKERP. The rhizosphere microbiome and plant health Trends in Plant Science, 2012, 17(8):478-486. DOI:10.1016/j.tplants.2012.04.001URL [本文引用: 1]
FAN ZQ, LU AQ, WANG HT, WANGY, DUAN HQ, LI HL, QIUG. Effects of Bacillus amyloliquefaciens on the growth of tobacco and microflora in rhizosphere soil Journal of Henan Agricultural Science, 2019, 48(4):33-40. (in Chinese) [本文引用: 1]
QIAOJ, YUX, LIANGX, LIUY, BORRISSR, LIUY. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome BMC Microbiology, 2017, 17(1):131. DOI:10.1186/s12866-017-1039-xURL [本文引用: 1]
GUOQ, LIS, LUX, LIB, MAP. PhoR/PhoP two component regulatory system affects biocontrol capability of Bacillus subtilis NCD-2 Genetics and Molecular Biology, 2010, 33(2):333-340. DOI:10.1590/S1415-47572010005000032URL [本文引用: 1]
ZHU WJ, WANGN, YU XP, WANGW. Effects of the biocontrol agent Pseudomonas fluorescens 2P24 on microbial community diversity in the melon rhizosphere Scientia Agricultura Sinica, 2010, 43(7):1389-1396. (in Chinese) [本文引用: 1]
ALIA, MOHANTA TK, ASAFS, REHMANN, AL-HOUSNIS, AL-HARRASIA, KHAN AL, AL-RAWAHIA. Biotransformation of benzoin by Sphingomonas sp. LK11 and ameliorative effects on growth of Cucumis sativus Archives of Microbiology, 2019, 201:591-601. DOI:10.1007/s00203-019-01623-1URL [本文引用: 2]
EL-LAITHY NA, BADAWY EA, YOUNESS ER, IBRAHIM AM, EL-NEMRM, EL-SHAMY KA. Antioxidant defense system as a protector against oxidative stress induced by thyroid dysfunction Der Pharmacia Lettre, 2016, 8(6):113-118. [本文引用: 1]
 ,1,2, 赵卫松2, 苏振贺2, 董丽红2, 郭庆港
,1,2, 赵卫松2, 苏振贺2, 董丽红2, 郭庆港

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT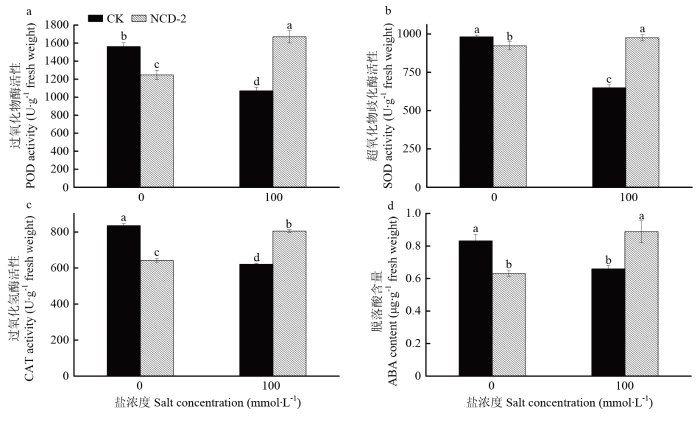 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT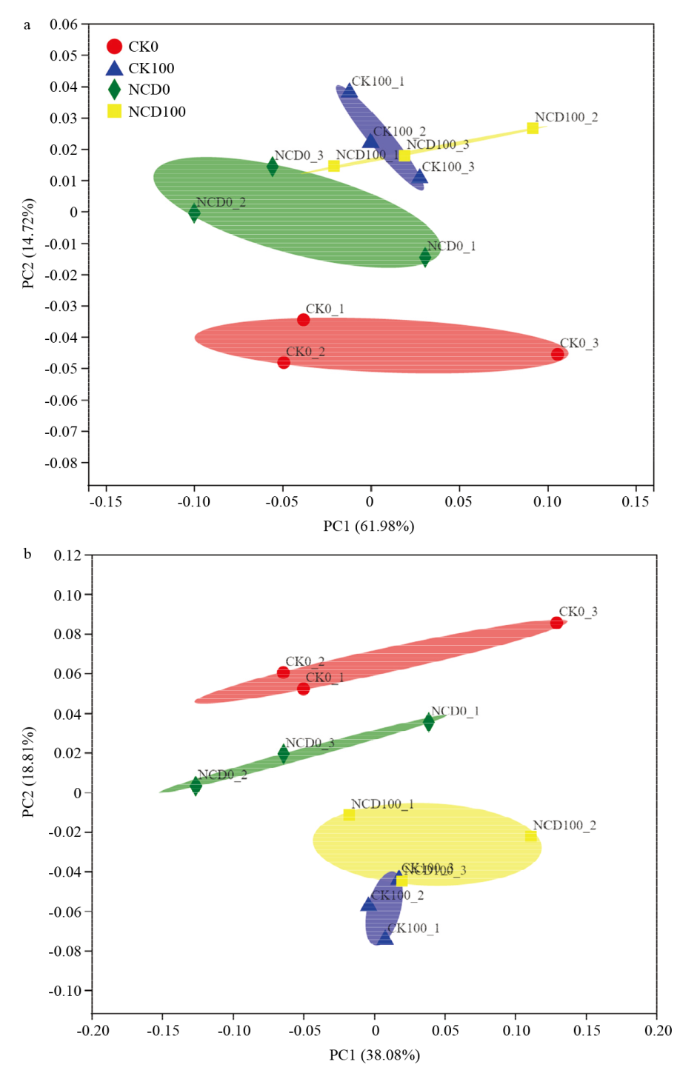 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT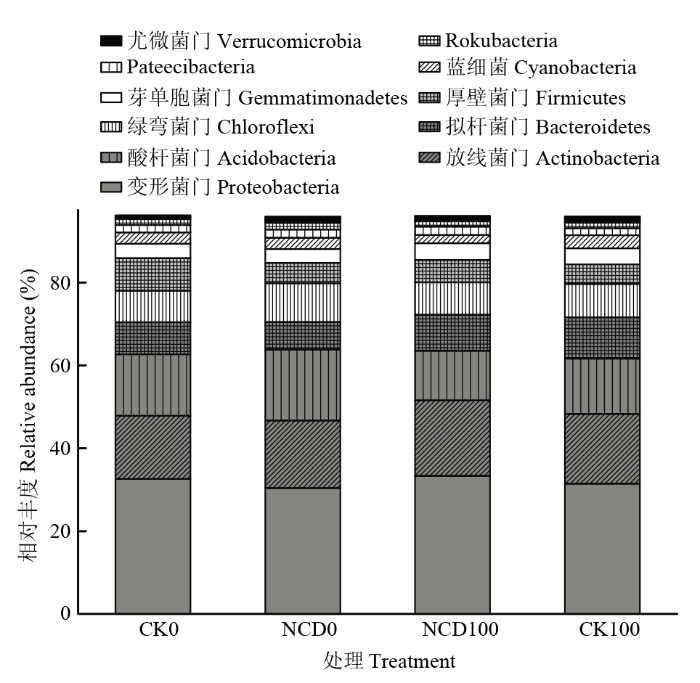 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT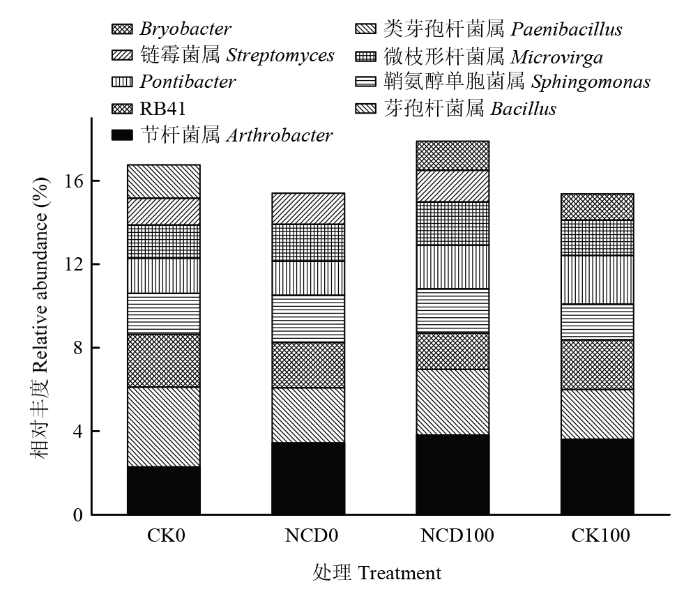 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT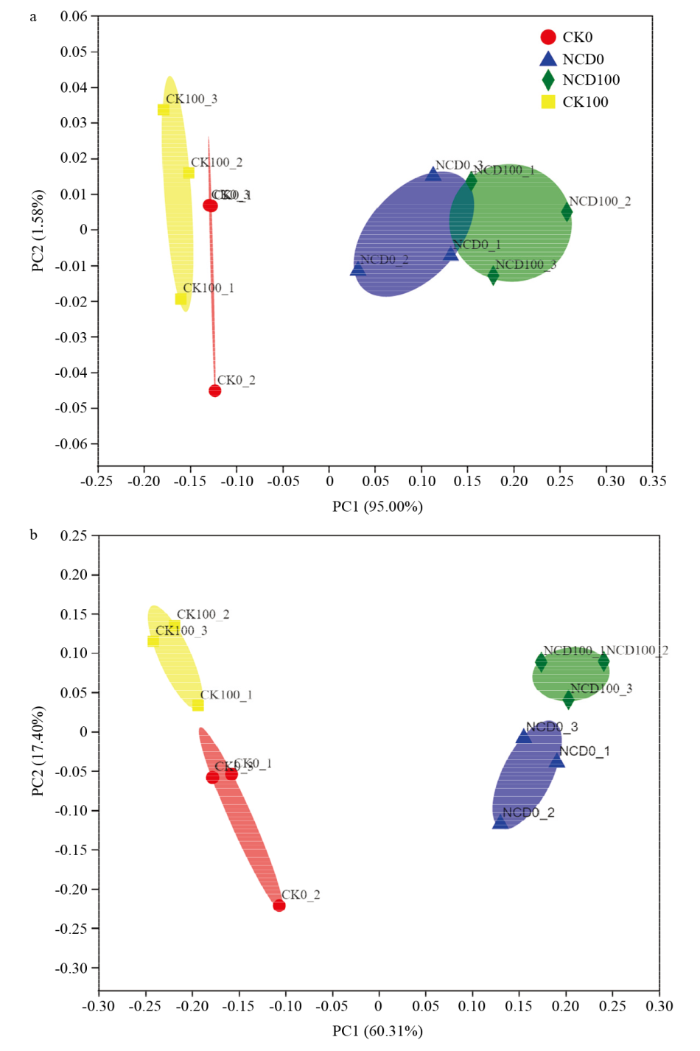 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}