Molecular Evolution and Function Analysis of bZIP Family in Nymphaea colorata
YE FangTing,, PAN XinFeng, MAO ZhiJun, LI ZhaoWei, FAN Kai,*College of Agriculture, FuJian Agriculture and Forestry University/Key Laboratory of Ministry of Education for Genetics, Breeding and Multiple Utilization of Crops, Fuzhou 350002
Abstract 【Objective】The genome-wide analysis of the bZIP family in Nymphaea colorata was used to identify the bZIP (basic leucine zipper) family members in waterlily, and then which were further analyzed. This study revealed molecular evolution and function of the bZIP family in waterlily.【Method】The genome sequence of N. colorata was downloaded from Waterlily Pond database. The bZIP members in waterlily were identified by using HMMER 3.0 program, and the conserved bZIP domain was verified by using CDD program. The phylogenetic tree was constructed by the IQ-tree software. The ExPASy and SOPMA online website were performed to analyze protein structure characters. The conserved motifs were identified by using MEME program. The gene duplication events were found and visualized by the MCScan and Circos software. Transcriptome data of NcbZIP members were obtained from the NCBI website (SRA Study: SRP222853). The Pearson Correlation Coefficient (PCC) about the expression levels of the NcbZIP family members was calculated by using R software, and the network of the expression levels in the NcbZIP family was analyzed by using Cytoscape software.【Result】46 bZIP members were identified in N. colorata, and were named from NcbZIP01 to NcbZIP46 according to their chromosome distributions. The A subfamily contained the most NcbZIP members (11 NcbZIPs). There were 10 subfamilies (A, B, C, D, E, G, H, I, J and S) according to the phylogenetic analysis. The protein length in the NcbZIP family was from 101 aa to 1 898 aa, and the molecular weight ranged from 12.04 kD to 214.64 kD. The NcbZIP members from same subfamily had the similar distributions of the conserved motifs and gene structures. Waterlily had 14 chromosomes, and 46 NcbZIP members were unevenly distributed across 10 chromosomes. Chromosome 1 had the highest number of NcbZIP members. There were 10 gene duplication events in the NcbZIP family, including nine segmental duplication events and one tandem duplication event. The A subfamily had the most number of the gene duplication events (three). Based on the expression patterns in different tissues, the NcbZIP family could be divided into three groups (I, II and III). The NcbZIP members in Group I were highly expressed in all tissues, while the NcbZIP members in group II were not expressed in almost all tissues. The NcbZIP members in group III had tissue-specific expression profiles, and most of NcbZIP members in C, D and E subfamilies belonged to group III. The PPC analysis about the expression levels of NcbZIP members indicated NcbZIP45 had the highest connection with other members.【Conclusion】46 NcbZIP members were identified in Nymphaea colorata, and were unevenly distributed in 14 chromosomes. The NcbZIP family could be divided into 10 subfamilies with conserved motifs distributions and diverse expression levels. The current study could lay the foundation on the functional analysis of the bZIP family in N. colorata. Keywords:Nymphaea colorata;bZIP family;molecular evolution;expression profile;function analysis
PDF (786KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 叶方婷, 潘鑫峰, 毛志君, 李兆伟, 范凯. 睡莲转录因子bZIP家族的分子进化以及功能分析. 中国农业科学, 2021, 54(21): 4694-4708 doi:10.3864/j.issn.0578-1752.2021.21.018 YE FangTing, PAN XinFeng, MAO ZhiJun, LI ZhaoWei, FAN Kai. Molecular Evolution and Function Analysis of bZIP Family in Nymphaea colorata. Scientia Agricultura Sinica, 2021, 54(21): 4694-4708 doi:10.3864/j.issn.0578-1752.2021.21.018
A:睡莲bZIP复制基因的共线性分析;B:睡莲bZIP复制基因的亚家族分布;C:睡莲bZIP复制基因的Ks分布,箭头指向Ks峰值 Fig. 4Syntenic analysis and Ks distribution of the duplicated bZIP members in Nymphaea colorata
A: Syntenic analysis of the duplicated bZIP members in Nymphaea colorata; B: Subfamily distribution of the duplicated bZIP members in waterlily; C: Ks distribution of the duplicated bZIPs in waterlily, the arrow points topeak value of Ks
Table 2 表2 表2睡莲复制的NcbZIP成员的Ka和Ks分析 Table 2Ka and Ks analysis for the duplicated NcbZIP in N. colorata
ZHANG LS, CHENF, ZHANG XT, LIZ, ZHAO YY, LOHAUSR, CHANG XJ, DONGW, HOS Y W, LIUX, SONGA X, CHENJ H, GUOW L, WANGZ J, ZHUANGY Y, WANGH F, CHENX Q, HUJ A, LIUY H, QINYet al. The water lily genome and the early evolution of flowering plants Nature, 2020, 577(7788):79-84. DOI:10.1038/s41586-019-1852-5URL [本文引用: 2]
DRÖGE-LASERW, SNOEK BL, SNELB, WEISTEC. The Arabidopsis bZIP transcription factor family-an update Current Opinion in Plant Biology, 2018, 45(Pt A):36-49. DOI:10.1016/j.pbi.2018.05.001URL [本文引用: 1]
LIU HJ, XUH, QIU WY, LI XF, ZHANGH, ZHUY, LI CS, WANG LC. Roles of bZIP transcription factors in plant growth and development and abiotic stress response Acta Agriculturae Zhejiangensis, 2019, 31(7):1205-1214. (in Chinese) [本文引用: 1]
WANG JY, DINGF, PAN JC, ZHANG SW, YANG YH, HUANGX, FAN ZY, LIL, WANGY. Research progress of bZIP lineage transcription factors in plant Chinese Journal of Tropical Agriculture, 2019, 39(6):39-45. (in Chinese) [本文引用: 1]
SORNARAJP, LUANGS, LOPATOS, HRMOVAM. Basic leucine zipper (bZIP) transcription factors involved in abiotic stresses: A molecular model of a wheat bZIP factor and implications of its structure in function Biochimica et Biophysica Acta, 2016, 1860(1 Pt A):46-56. [本文引用: 1]
CUI RX, ZHANG YW, CHEN XQ, GU CH, ZHANGQ. The Latest Research Progress on the Stress Responses of bZIP Involved in Plants Biotechnology Bulletin, 2019, 35(2):143-155. (in Chinese) [本文引用: 1]
DASP, LAKRAN, NUTAN KK, SINGLA-PAREEKS L, PAREEKA. A unique bZIP transcription factor imparting multiple stress tolerance in Rice Rice (New York, NY), 2019, 12(1):58. [本文引用: 1]
RONG SY, WU ZY, CHENG ZZ, ZHANGS, LIUH, HUANG QM. Genome-wide identification, evolutionary patterns, and expression analysis of bZIP gene family in olive (Olea europaea L.) Genes (Basel), 2020, 11(5):510. DOI:10.3390/genes11050510URL [本文引用: 1]
WANG ZH, YAN LY, WAN LY, HUAI DX, KANG YP, SHIL, JIANG HF, LEIY, LIAO BS. Genome-wide systematic characterization of bZIP transcription factors and their expression profiles during seed development and in response to salt stress in peanut BMC Genomics, 2019, 20(1):51. DOI:10.1186/s12864-019-5434-6URL [本文引用: 1]
LIU YH, CHAI MN, ZHANGM, HEQ, SU ZX, PRIYADARSHANIS V G N, LIUL P, DONGG X, QINY A. Genome-wide analysis, characterization, and expression profile of the basic leucine zipper transcription factor family in pineapple International Journal of Genomics, 2020, 2020:3165958. [本文引用: 1]
AZEEMF, TAHIRH, IJAZU, SHAHEENT. A genome-wide comparative analysis of bZIP transcription factors in G. arboreum and G. raimondii (Diploid ancestors of present-day cotton) Physiology and Molecular Biology of Plants, 2020, 26(3):433-444. DOI:10.1007/s12298-020-00771-9URL [本文引用: 1]
WANG WW, WANG YF, ZHANG SM, XIE KL, ZHANGC, XI YJ, SUN FL. Genome-wide analysis of the abiotic stress-related bZIP family in switchgrass Molecular Biology Reports, 2020, 47(6):4439-4454. DOI:10.1007/s11033-020-05561-wURL [本文引用: 2]
FANK, WANGM, MIAOY, NIM, BIBIN, YUAN SN, LIF, WANG XD. Molecular evolution and expansion analysis of the NAC transcription factor in Zea mays PLoS One, 2014, 9(11):e111837. DOI:10.1371/journal.pone.0111837URL [本文引用: 2]
PERTEAM, KIMD, PERTEA GM, LEEK JT, SALZBERG SL. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown Nature Protocols, 2016, 11(9):1650-1667. DOI:10.1038/nprot.2016.095URL [本文引用: 1]
CHAI WB, SI WN, JIW, QIN QQ, ZHAO ML, JIANG HY. Genome-wide investigation and expression profiling of HD-zip transcription factors in foxtail millet (Setaria italica L.) BioMed Research International, 2018, 2018:8457614. [本文引用: 1]
YANGY, YU TF, MAJ, CHENJ, ZHOU YB, CHENM, MA YZ, WEI WL, XU ZS. The soybean bZIP transcription factor gene GmbZIP2 confers drought and salt resistances in transgenic plants International Journal of Molecular Sciences, 2020, 21(2):670. DOI:10.3390/ijms21020670URL [本文引用: 1]
ZHU YY, XUEB, WANG AQ, WANG WJ, ZHOUA, HUANG SX, LIU YS. Comprehensive bioinformatic analysis of bZIP transcription factors in Solanum lycopersicum Chinese Journal of Applied & Environmental Biology, 2014, 20(5):767-774. (in Chinese) [本文引用: 1]
WANG SJ, SUNH, DANGH. Genome-wide analysis of the bZIP transcription factors in Populus in response to salt stress Journal of Shanxi Agricultural University (Natural Science Edition), 2018, 38(8):1-7, 14. (in Chinese) [本文引用: 1]
GAOB, CHEN JJ, CUI SL, HOU MY, MU GJ, CHEN HY, YANG XL, LIU LF. Genome-wide identification and expression analysis of bZIP gene family under drought stress in peanut Journal of Plant Genetic Resources, 2020, 21(1):174-191. (in Chinese) [本文引用: 1]
BAILLO EH, KIMOTHO RN, ZHANG ZB, XUP. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement Genes (Basel), 2019, 10:771. DOI:10.3390/genes10100771URL [本文引用: 1]
E ZG, ZHANGY P, ZHOUJ H, WANGL. Mini review roles of the bZIP gene family in rice Genetics and Molecular Research, 2014, 13(2):3025-3036. DOI:10.4238/2014.April.16.11PMID:24782137 [本文引用: 1] The basic leucine zipper (bZIP) genes encode transcription factors involved in the regulation of various biological processes. Similar to WRKY, basic helix-loop-helix, and several other groups of proteins, the bZIP proteins form a superfamily of transcription factors that mediate plant stress responses. In this review, we present the roles of bZIP proteins in multiple biological processes that include pathogen defense; responses to abiotic stresses; seed development and germination; senescence; and responses to salicylic, jasmonic, and abscisic acids in rice. We also examined the characteristics of the bZIP proteins and their genetic composition. To ascertain the evolutionary changes in and functions of this supergene family, we performed an exhaustive comparison among the 89 rice bZIP genes that were previously described and those more recently listed in the MSU Rice Genome Annotation Project Database using a Hidden Markov Model. We excluded 3 genes from the list, resulting in a total of 86 bZIP genes in japonica rice.
PANF, WUM, HU WF, LIUR, YAN HW, XIANGY. Genome-Wide Identification and Expression Analyses of the bZIP Transcription Factor Genes in moso bamboo (Phyllostachys edulis) International Journal of Molecular Sciences, 2019, 20(9):2203. DOI:10.3390/ijms20092203URL [本文引用: 1]
ZHANGM, LIU YH, SHIH, GUO ML, CHAI MN, HEQ, YAN MK, CAOD, ZHAO LH, CAI HY, QIN YA. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family BMC Genomics, 2018, 19(1):159. DOI:10.1186/s12864-018-4511-6URL [本文引用: 1]
YUJ, HU SN, WANGJ, KA-SHUG, LI SG, LIUB, DENG YJ, DAIL, ZHOUY, ZHANG XQ, CAO ML, LIUJ, SUN JD, TANG JB, CHEN YJ, HUANG XB, LINW, YEC, TONGW, CONG LJ, et al. A Draft Sequence of the Rice Genome (Oryza sativa L. ssp. indica) Science, 2002, 296(5565):79-92. DOI:10.1126/science.1068037URL [本文引用: 1]
SCHMUTZJ, CANNON SB, SCHLUETERJ, MA JX, MITROST, NELSONW, HYTEN DL, SONG QJ, THELEN JJ, CHENG JL, XUD, HELLSTENU, MAY GD, YUY, SAKURAIT, UMEZAWAT, BHATTACHARYYA MK, SANDHUD, VALLIYODANB, LINDQUISTE, et al. Genome sequence of the palaeopolyploid soybean Nature, 2010, 463(7278):178-183. DOI:10.1038/nature08670URL [本文引用: 1]
WEI KF, CHENJ, WANG YM, CHEN YH, CHEN SX, LIN YN, PANS, ZHONG XJ, XIE DX. Genome-wide analysis of bZIP-encoding genes in maize DNA Research, 2012, 19(6):463-476. DOI:10.1093/dnares/dss026URL [本文引用: 1]
LIU MY, WEN YD, SUN WJ, MA ZT, HUANGL, WUQ, TANG ZZ, BU TL, LI CL, CHENH. Genome-wide identification, phylogeny, evolutionary expansion and expression analyses of bZIP transcription factor family in tartaty buckwheat BMC Genomics, 2019, 20(1):483. DOI:10.1186/s12864-019-5882-zURL [本文引用: 1]
LI DY, FU FY, ZHANG HJ, SONG FM. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato (Solanum lycopersicum L.) BMC Genomics, 2015, 16:771. DOI:10.1186/s12864-015-1990-6URL [本文引用: 1]
ZHAOP, YE MH, WANG RQ, WANG DD, CHENQ. Systematic identification and functional analysis of potato (Solanum tuberosum L.) bZIP transcription factors and overexpression of potato bZIP transcription factor StbZIP-65 enhances salt tolerance International Journal of Biological Macromolecules, 2020, 161:155-167. DOI:10.1016/j.ijbiomac.2020.06.032URL [本文引用: 1]
FANK, CHEN YR, MAO ZJ, FANGY, LI ZW, LIN WW, ZHANG YQ, LIU JP, HUANG JW, LIN WX. Pervasive duplication, biased molecular evolution and comprehensive functional analysis of the PP2C family in Glycine max BMC Genomics, 2020, 21(1):465. DOI:10.1186/s12864-020-06877-4URL [本文引用: 1]
FANK, MAO ZJ, ZHENG JX, CHEN YR, LI ZW, LIN WW, ZHANG YQ, HUANG JW, LIN WX. Molecular evolution and expansion of the KUP family in the allopolyploid cotton species Gossypium hirsutum and Gossypium barbadense Frontiers in Plant Science, 2020, 11:545042. DOI:10.3389/fpls.2020.545042URL [本文引用: 1]
LIANG CZ, MENG ZH, MENG ZG, MALIKW, YANR, LWIN KM, LIN FZ, WANG YA, SUN GQ, ZHOUT, ZHUT, LI JY, JIN SX, GUO SD, ZHANGR. GhABF2, a bZIP transcription factor, confers drought and salinity tolerance in cotton (Gossypium hirsutum L.) Scientific Reports, 2016, 6:35040. DOI:10.1038/srep35040URL [本文引用: 1]
LIM CW, BAEKW, JUNGJ, KIM JH, LEE SC. Function of ABA in stomatal defense against biotic and drought stresses International Journal of Molecular Sciences, 2015, 16(7):15251-15270. DOI:10.3390/ijms160715251URL [本文引用: 1]
NAKASHIMAK, YAMAGUCHI-SHINOZAKIK. ABA signaling in stress-response and seed development Plant Cell Reports, 2013, 32(7):959-970. DOI:10.1007/s00299-013-1418-1URL [本文引用: 1]
GUO GH, LIU HY, LI GH, LIUM, LIY, WANG SH, LIU ZH, TANGS, DING YF. Analysis of physiological characteristics about ABA alleviating rice booting stage drought stress Scientia Agricultura Sinica, 2014, 47(22):4380-4391. (in Chinese) [本文引用: 1]
SHAN YS, DAI HH, HEX, XIN ZQ, WU NB. Effects of exogenous methyl jasmonate and salicylic acid on physiological characteristics and secondary metabolism of Atropa belladonna under NaCl stress Plant Physiology Communications, 2019, 55(9):1335-1346. (in Chinese) [本文引用: 1]
YU XX, ZHANG WJ, ZHANGY, ZHANG XJ, LANG DY, ZHANG XH. The roles of methyl jasmonate to stress in plants Functional Plant Biology, 2019, 46(3):197-212. DOI:10.1071/FP18106URL [本文引用: 1]
HO TT, MURTHY HN, PARK SY. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures International Journal of Molecular Sciences, 2020, 21(3):716. DOI:10.3390/ijms21030716URL [本文引用: 1]
SCHLÖGL PS, NOGUEIRA F TS, DRUMMONDR, FELIX JM, DE ROSA VE, VICENTINIR, LEITEA, ULIAN EC, MENOSSIM. Identification of new ABA- and MEJA-activated sugarcane bZIP genes by data mining in the SUCEST database Plant Cell Reports, 2008, 27(2):335-345. DOI:10.1007/s00299-007-0468-7URL [本文引用: 1]
YANG ZM, SUNJ, CHENY, ZHU PP, ZHANGL, WU SY, MA DF, CAO QH, LI ZY, XUT. Genome-wide identification, structural and gene expression analysis of the bZIP transcription factor family in sweet potato wild relative Ipomoea trifida BMC Genetics, 2019, 20(1):41. DOI:10.1186/s12863-019-0743-yURL [本文引用: 1]
MURMUJ, BUSH MJ, DELONGC, LI ST, XU ML, KHANM, MALCOLMSONC, FOBERT PR, ZACHGOS, HEPWORTH SR. Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development Plant Physiology, 2010, 154(3):1492-1504. DOI:10.1104/pp.110.159111URL [本文引用: 1]
XU DB, CHENM, MA YN, XU ZS, LI LC, CHEN YF, MA YZ. A G-protein β subunit, AGB1, negatively regulates the ABA response and drought tolerance by down-regulating AtMPK6-related pathway in Arabidopsis PLoS ONE, 2015, 10(1):e0116385. DOI:10.1371/journal.pone.0116385URL [本文引用: 1]
LIU DF, SHI SP, HAO ZJ, XIONG WT, LUO MZ. A homologue of Arabidopsis VIP1, may positively regulate JA levels by directly targetting the genes in JA signaling and metabolism pathway in rice International Journal of Molecular Sciences, 2019, 20(9):2360. DOI:10.3390/ijms20092360URL [本文引用: 1]
 ,, 潘鑫峰, 毛志君, 李兆伟, 范凯
,, 潘鑫峰, 毛志君, 李兆伟, 范凯

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT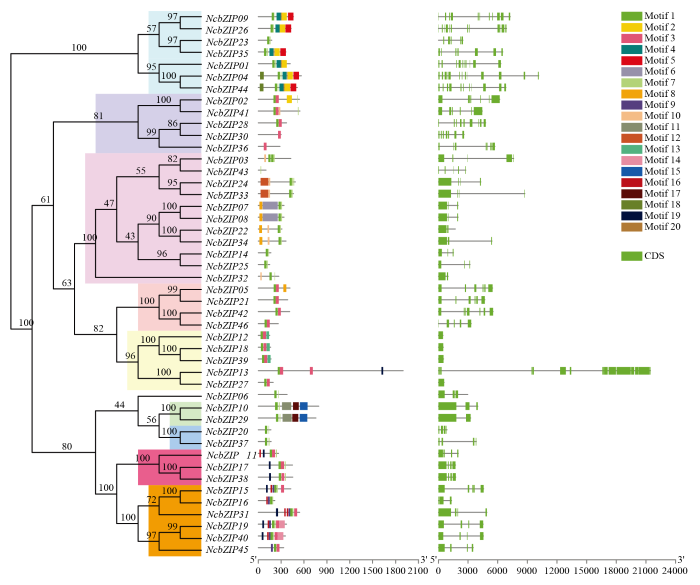 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT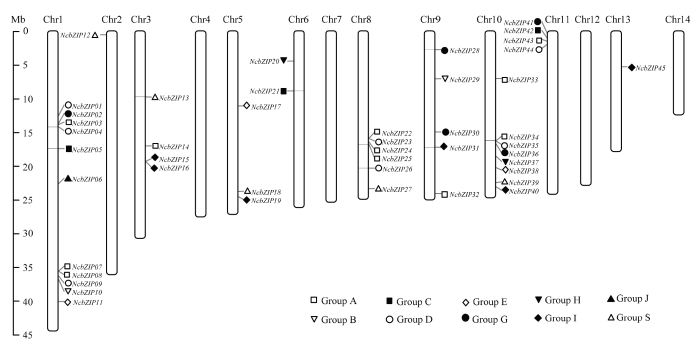 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT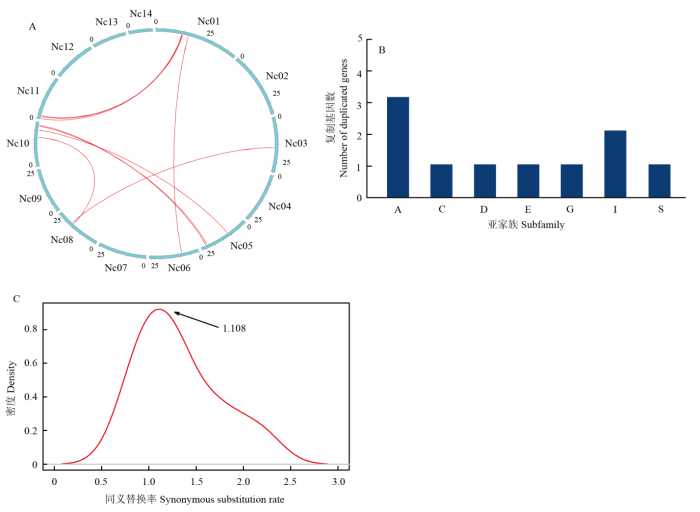 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT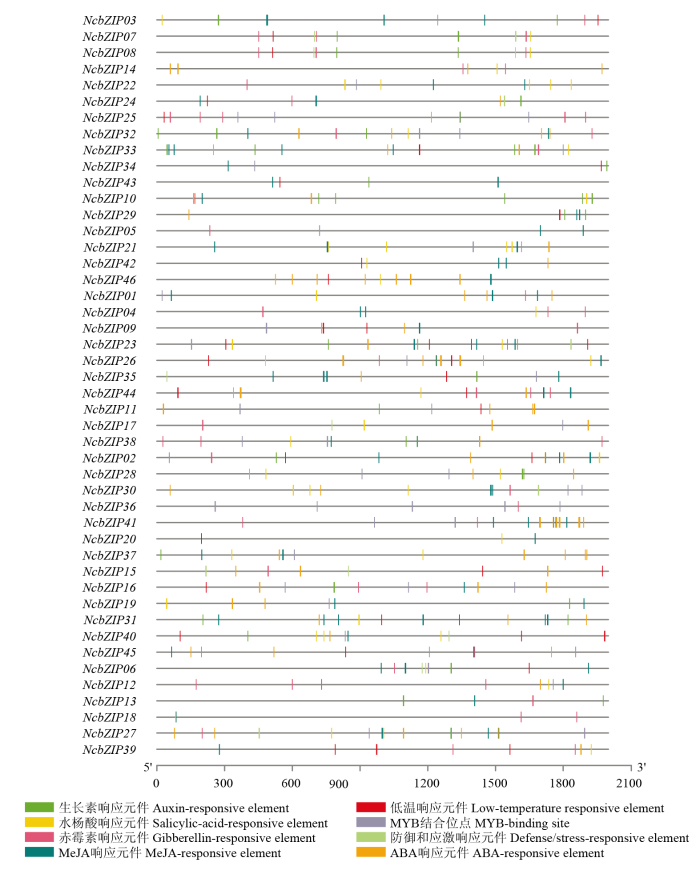 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT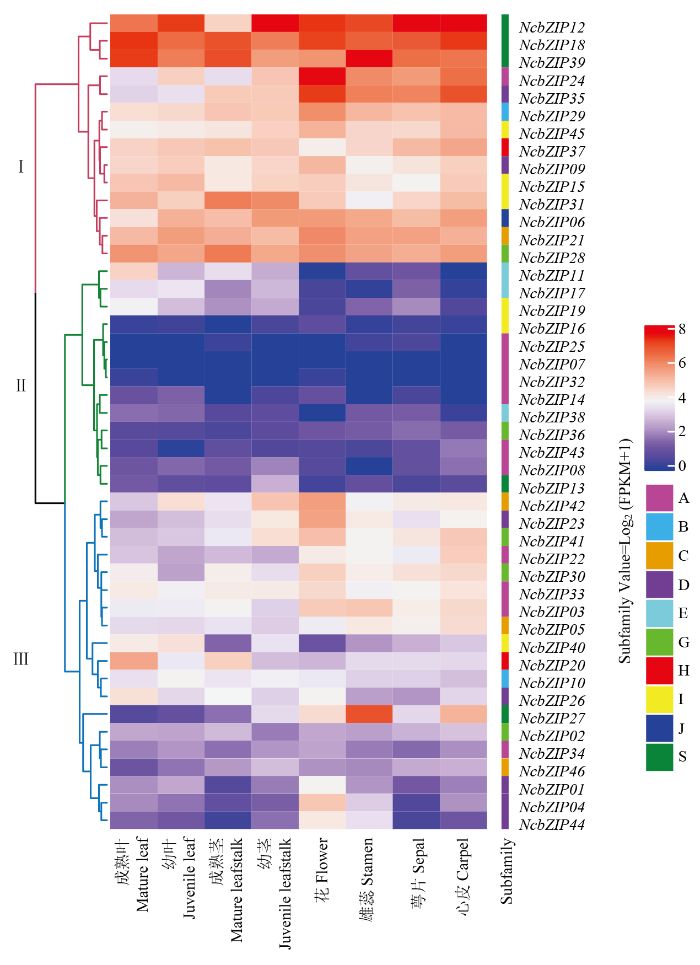 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT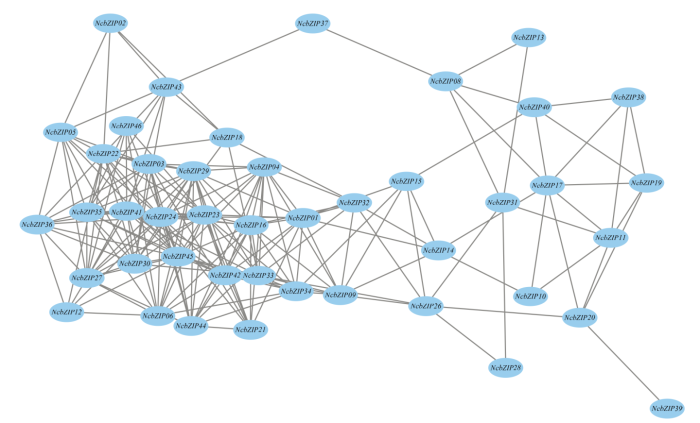 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}