Transcription Factor TEAD4 Regulates Early Embryonic Development in Pigs
ZHANG DanDan,, XU TengTeng, GAO Di, QI Xin, NING Wei, RU ZhenYuan, ZHANG XiangDong, GUO TengLong, SHENTU LuYan, YU Tong, MA YangYang, LI YunSheng, ZHANG YunHai, CAO ZuBing,Key Laboratory of Conservation and Biological Breeding of Local Livestock and Poultry Genetic Resources/College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036
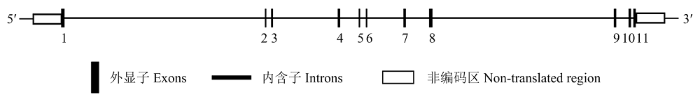
Abstract 【Background】 TEA domain transcription factor 4 (TEAD4) is known to be a member of the TEAD family of transcription factors and plays a key role in determining the characteristics of the preimplantation embryo in rodents. In mouse embryos, it was found to be involved in regulating the genealogical differentiation of trophectoderm cells in preimplantation embryos by promoting Cdx2 expression. The absence of the TEAD4 gene in mouse embryos can lead to failure of mouse blastocyst formation. However, the role of TEAD4 in early porcine embryonic development is still unclear. 【Objective】This study aimed to preliminarily elucidate the effect of TEAD4 on early porcine embryonic development, in order to lay the theoretical foundation for further exploring the molecular mechanisms of transcription factors on early porcine embryonic development. 【Method】In this study, the bioinformatics analysis of the porcine TEAD4 gene was performed by using web-based tools, including analysis of the porcine TEAD4 gene sequence, comparison of homology between pigs, human and mice, and comparison of the evolutionary relationship of TEAD4 between different species. The role of TEAD4 in early embryonic development in pigs was then tested. The mRNA expression level of TEAD4 gene in porcine oocytes and the early embryos was firstly detected by fluorescence quantitative PCR. and then, siRNA targeting TEAD4 was designed and injected into mature oocytes by microinjection technique to reduce the level of endogenous TEAD4 gene in the oocyte cytoplasm, and to determine that TEAD4 siRNA acts only on TEAD4 gene, with a view to determining the role of TEAD4 gene in early porcine embryonic development. 【Result】Sequence analysis showed that the porcine TEAD4 gene contained 11 exons and localized on chromosome 5, with spanning 37.188 kb, 1 473 bp in full mRNA length, and 1305 bp in full coding region, which encoded 434 amino acids. Homology analysis with human and mouse revealed that TEAD4 was highly conserved in different species and had the closest affinity on pig and cow. The results of fluorescence quantitative PCR showed that TEAD4 mRNA was expressed in both porcine oocytes and early embryos; compared with GV-stage oocytes, the expression of TEAD4 mRNA was lowest in MII-stage oocytes and remained low until the 4-cell stage, but reached the highest expression in the 8-cell stage, and then gradually decreased in the morula and blastocyst stages. Microinjection of siRNA targeting TEAD4 revealed that TEAD4 siRNA only acted on the endogenous TEAD4 gene in oocytes, but not on TEAD1 and TEAD3, and compared with the control and negative control siRNA groups, the injection of TEAD4 siRNA significantly reduced TEAD4 mRNA expression at the 8-cell and morula embryo periods. When TEAD4 gene expression was knocked down, observation of the developmental efficiency of porcine orphan activation and in vitro fertilization embryos showed that the developmental efficiency of TEAD4 siRNA knockdown group from 8-cell to blastocyst stage was significantly reduced compared to the control and negative control siRNA groups. 【Conclusion】 The results of this study indicated that the TEAD4 gene was highly conserved across species, with the closest affinity on pigs and bovine, and that TEDA4 might be involved in regulating the development of early porcine embryos.. Keywords:pig;early embryos;TEAD4;parthenogenetic activation;in vitro fertilization
PDF (1144KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张丹丹, 许腾腾, 高迪, 齐昕, 宁伟, 汝振远, 张翔栋, 郭腾龙, 申屠璐燕, 于童, 马洋洋, 李运生, 张运海, 曹祖兵. 转录因子TEAD4对猪早期胚胎发育的调控. 中国农业科学, 2021, 54(20): 4456-4465 doi:10.3864/j.issn.0578-1752.2021.20.018 ZHANG DanDan, XU TengTeng, GAO Di, QI Xin, NING Wei, RU ZhenYuan, ZHANG XiangDong, GUO TengLong, SHENTU LuYan, YU Tong, MA YangYang, LI YunSheng, ZHANG YunHai, CAO ZuBing. Transcription Factor TEAD4 Regulates Early Embryonic Development in Pigs. Scientia Acricultura Sinica, 2021, 54(20): 4456-4465 doi:10.3864/j.issn.0578-1752.2021.20.018
Figure Aand B: Effects of TEAD4 siRNA on TEAD4 mRNA expression in 8-cell and morula embryo. Figure Cand D: Effects of TEAD4 siRNA on mRNA expression of TEAD1 and TEAD3. a and b indicate significant differences (P<0.05). The same as below
YANQ M, LAIL X. Progress of genetically-modified pigs in the human health related research China Basic Science, 2015, 17(5):20-27. doi: 10.3969/j.issn.1009-2412.2015.05.004. (in Chinese) [本文引用: 1]
LIW L, BAOL, XIAOL. The progress of genetic-modified pigs as donors in xenotransplantation Chinese Journal of Cell Biology, 2014, 36(9):1300-1305. (in Chinese) [本文引用: 1]
HUANGY Q, LIG L, YANGH Q, WUZ F. Progress and application of genome-edited pigs in biomedical research Hereditas, 2018, 40(8):632-646. doi: 10.16288/j.yczz.18-026. (in Chinese) [本文引用: 1]
CHENB Z, GUP, JIAJ S, LIUW, LIUY M, XUT, LINX L, LINT Y, LIUY, CHENH W, XUM C, YUANJ, ZHANGJ N, ZHANGY H, XIAOD, GUW W. Optimization strategy for generating gene-edited tibet minipigs by synchronized oestrus and cytoplasmic microinjection International Journal of Biological Sciences, 2019, 15(12):2719-2732. DOI:10.7150/ijbs.35930URL [本文引用: 1]
JUNS M, PARKM, LEEJ Y, JUNGS, LEEJ E, SHIMS H, SONGH, LEED R. Single cell-derived clonally expanded mesenchymal progenitor cells from somatic cell nuclear transfer-derived pluripotent stem cells ameliorate the endometrial function in the uterus of a murine model with Asherman's syndrome Cell Proliferation, 2019, 52(3):e12597. doi: 10.1111/cpr.12597. URL [本文引用: 1]
PFEFFERP L. Building principles for constructing a mammalian blastocyst embryo Biology (Basel), 2018, 23; 7(3):41. [本文引用: 2]
OESTRUPO, HALLV, PETKOVS G, WOLFX A. HYLDIGS, HYTTELP. From zygote to implantation: morphological and molecular dynamics during embryo development in the pig Reproduction in Domestic Animals, 2009, 44(Suppl 3):39-49. DOI:10.1111/rda.2009.44.issue-s3URL [本文引用: 1]
SAIZN, PLUSAB. Early cell fate decisions in the mouse embryo Development (Cambridge, England), 2013, 145(3):R65-R80. doi: 10.1530/rep-12-0381. [本文引用: 1]
YAMANAKAY, RALSTONA, STEPHENSONR O, ROSSANTJ. Cell and molecular regulation of the mouse blastocyst Developmental Dynamics, 2006, 235(9):2301-2314. doi: 10.1002/dvdy.20844. URL [本文引用: 1]
CHENL, YABUUCHIA, EMINLIS, TAKEUCHIA, LUC W, HOCHEDLINGERK, DALEYG Q. Cross-regulation of the Nanog and Cdx2 promoters Cell Research, 2009, 19(9):1052-1061. doi: 10.1038/cr.2009.79. URL [本文引用: 1]
NISHIOKAN, INOUEK, ADACHIK, KIYONARIH, OTAM, RALSTONA, YABUTAN, HIRAHARAS, STEPHENSONR O, OGONUKIN, MAKITAR, KURIHARAH, MORIN-KENSICKIE M, NOJIMAH, ROSSANTJ, NAKAOK, NIWAH. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass Developmental Cell, 2009, 16(3):398-410. doi: 10.1016/j.devcel.2009.02.003. URL [本文引用: 1]
BOUG, LIUS, GUOJ, ZHAOY, SUNM, XUEB, WANGJ, WEIY, KONGQ, LIUZ. Cdx2 represses Oct4 function via inducing its proteasome-dependent degradation in early porcine embryos Developmental Biology, 2016, 410(1):36-44. doi: 10.1016/j.ydbio.2015.12.014. URL [本文引用: 2]
CAOZ B, XUT T, TONGX, WANGY Q, ZHANGD D, GAOD, ZHANGL, NINGW, QIX, MAY Y, YUT, KNOTTJ G, ZHANGY H. Maternal Yes-associated protein participates in porcine blastocyst development via modulation of trophectoderm epithelium barrier function Cell, 2019, 8(12):1606. [本文引用: 1]
LINK C, PARKH W, GUANK L. Regulation of the hippo pathway transcription factor TEAD Trends in Biochemical Sciences, 2017, 42(11):862-872. doi: 10.1016/j.tibs.2017.09.003. URL [本文引用: 1]
HOIDENJ K, CUNNINGHAMC N. Targeting the Hippo Pathway and Cancer through the TEAD Family of Transcription Factors Cancers(Basel), 2018, 10(3):81. [本文引用: 1]
ZHAOB, YEX, YUJ, LIL, LIW, LIS, YUJ, LINJ D, WANGC Y, CHINNAIYANA M, LAIZ C, GUANK L. TEAD mediates YAP- dependent gene induction and growth control Genes & Development, 2008, 22(14):1962-1971. doi: 10.1101/gad.1664408. URL [本文引用: 1]
TSIKAR W, MAL, KEHATI, SCHRAMMC, SIMMERG, MORGANB, FINED M, HANFTL M, MCDONALDK S, MOLKENTINJ D, KRENZM, YANGS, JIJ. TEAD-1 overexpression in the mouse heart promotes an age-dependent heart dysfunction The Journal of Biological Chemistry, 2010, 285(18):13721-13735. doi: 10.1074/jbc.m109.063057. URL [本文引用: 1]
HOMEP, SAHAB, RAYS, DUTTAD, GUNEWARDENAS, YOOB, PALA, VIVIANJ L, LARSONM, PETROFFM, GALLAGHERP G, SCHULZV P, WHITEK L, GOLOST G, BEHRB, PAULS. Altered subcellular localization of transcription factor TEAD4 regulates first mammalian cell lineage commitment Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(19):7362-7367. doi: 10.1073/pnas.1201595109. [本文引用: 2]
YAGIR, KOHNM J, KARAVANOVAI, KANEKOK J, VULLHORSTD, DEPAMPHILISM L, BUONANNOA. Transcription factor TEAD4 specifies the trophectoderm lineage at the beginning of mammalian development Development (Cambridge, England), 2007, 134(21):3827-3836. doi: 10.1242/dev.010223. URL [本文引用: 2]
NISHIOKAN, YAMAMOTOS, KIYONARIH, SATOH, SAWADAA, OTAM, NAKAOK, SASAKIH. Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos Mechanisms of Development, 2008, 125(3/4):270-283. doi: 10.1016/ j.mod.2007.11.002. URL [本文引用: 2]
KUMARRAM P, SOMAR, PRATIKH, BISWARUPS, BHASWATIB, WILKINSHEATHER M, HEMANTKUMARC, AVISHEKG, JESSICAM F, ARINDAMP, PARTHAK, SWERDLOWRUSSELL H, SOUMENP. Regulation of energy metabolism during early mammalian development: TEAD4 controls mitochondrial transcription Development, 2018, 145(19): dev162644. doi: 10.1242/dev.162644. [本文引用: 2]
KANEKOK J, DEPAMPHILISM L. TEAD4 establishes the energy homeostasis essential for blastocoel formation Development (Cambridge, England), 2013, 140(17):3680-3690. doi: 10.1242/ dev.093799. URL [本文引用: 2]
AKIZAWAH, KOBAYASHIK, BAIH, TAKAHASHIM, KAGAWAS, NAGATOMOH, KAWAHARAM. Reciprocal regulation of TEAD4 and CCN2 for the trophectoderm development of the bovine blastocyst Reproduction (Cambridge, England), 2018, 155(6):563-571. doi: 10.1530/rep-18-0043. URL [本文引用: 1]
SAKURAIN, TAKAHASHIK, EMURAN, HASHIZUMET, SAWAIK. Effects of downregulating TEAD4 transcripts by RNA interference on early development of bovine embryos The Journal of Reproduction and Development, 2017, 63(2):135-142. DOI:10.1262/jrd.2016-130URL [本文引用: 2]
BENW R. Expression of Tead1-3 in Mouse Uterus during peri-i mplantation period [D]. Harbin: Northeast Agricultural University, 2014. (in Chinese) [本文引用: 1]
XIAOL, MAL, WANGZ, YUY, LYES J, SHANY, WEIY. Deciphering a distinct regulatory network of TEAD4, CDX2 and GATA3 in humans for trophoblast transition from embryonic stem cells Biochimica et Biophysica Acta Molecular Cell Research, 2020, 1867(9):118736. doi: 10.1016/j.bbamcr.2020.118736. URL [本文引用: 1]
 ,, 许腾腾, 高迪, 齐昕, 宁伟, 汝振远, 张翔栋, 郭腾龙, 申屠璐燕, 于童, 马洋洋, 李运生, 张运海, 曹祖兵
,, 许腾腾, 高迪, 齐昕, 宁伟, 汝振远, 张翔栋, 郭腾龙, 申屠璐燕, 于童, 马洋洋, 李运生, 张运海, 曹祖兵

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT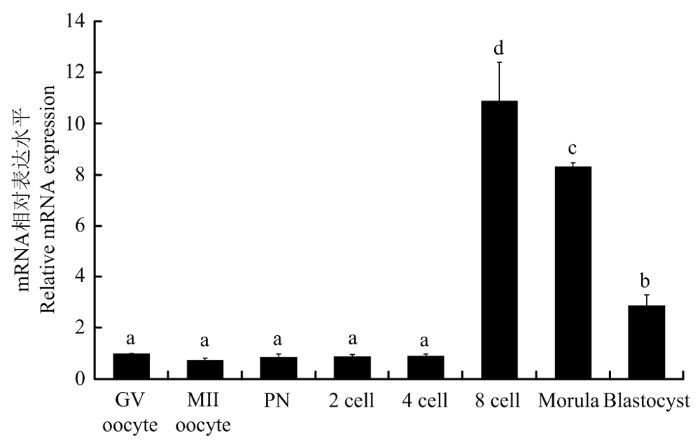 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT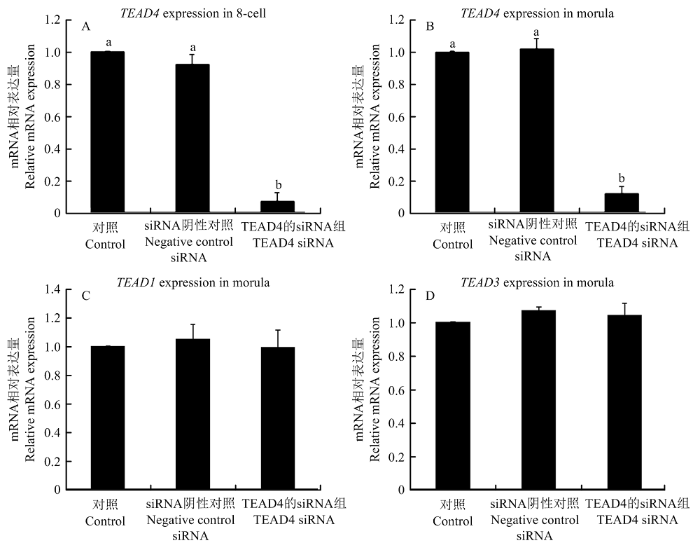 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}