 ,1, 陈银基,1, 周光宏2
,1, 陈银基,1, 周光宏2The Mechanism of Myofibrillar Protein Gel Functionality Influenced by Modified Sugarcane Dietary Fiber
ZHUANG XinBo,1, CHEN YinJi,1, ZHOU GuangHong2通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-11-3接受日期:2021-03-24
| 基金资助: |
Received:2020-11-3Accepted:2021-03-24
作者简介 About authors
庄昕波,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (4344KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
庄昕波, 陈银基, 周光宏. 改性甘蔗膳食纤维对猪肉肌原纤维蛋白凝胶特性的影响[J]. 中国农业科学, 2021, 54(15): 3320-3330 doi:10.3864/j.issn.0578-1752.2021.15.015
ZHUANG XinBo, CHEN YinJi, ZHOU GuangHong.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】香肠是一种典型的肉糜制品,由瘦肉和脂肪经斩拌灌肠后蒸煮而成。香肠配方中含有高达30%的动物脂肪[1]。有关研究发现,过量动物脂肪摄入会增加肥胖、心血管疾病、高血压、冠心病等一系列“富贵病”的发生几率[2,3,4]。因此,为了迎合消费者绿色健康的饮食需求,开发科学组配、健康营养的低脂香肠是目前肉制品加工行业发展的新趋势,这对保障居民的身体健康具有重要的意义。【前人研究进展】香肠中的脂肪具有重要的加工特性,表现为以共聚物的形式填充在蛋白凝胶网络中以提高凝胶强度、增强香肠的多汁性以及生成芳香类物质[5,6,7]。因此,直接降低脂肪含量或单纯用水替代脂肪都会导致香肠食用品质急剧下降,难以被消费者接受。利用不溶性膳食纤维替代脂肪开发低脂香肠是当前加工肉制品行业的研究热点。从营养角度,膳食纤维被誉为“第七大营养素”,能够改善肠道益生菌群和预防结肠癌[5]。世界卫生组织建议膳食纤维每日摄入量应达到25 g[3,5]。从加工特性角度,添加不溶性膳食纤维有利于改善低脂香肠的质构和降低香肠的蒸煮损失。相关研究显示,在鱼糜凝胶中添加不溶性膳食纤维可以有效提高产品的品质特性,例如凝胶强度和持水性[8]。此外,豆壳膳食纤维作为脂肪替代物能够有效降低低脂猪肉馅饼中的胆固醇含量,同时,处理组中水分含量和碳水化合物含量显著提高。添加豆壳膳食纤维处理组相比对照组,其质构和蒸煮损失都显著降低;同时,色泽、嫩度、多汁性、馅饼风味和顾客接受程度并没有发生显著变化[9]。【本研究切入点】当前肉制品行业的研究重点在于探究不同植物源不溶性膳食纤维对低脂香肠品质的改善效果[9,10,11,12]。而不溶性膳食纤维对低脂香肠凝胶特性的影响机制还鲜有研究。目前,关于多糖改善肌原纤维蛋白凝胶特性的机制主要是膨胀效应(淀粉类)和填充效应(凝胶性多糖)[13,14,15]。不溶性膳食纤维具有稳定的物理结构,既不能像淀粉一样通过糊化膨胀效应改善凝胶强度,也不能像魔芋胶一样通过水合作用形成凝胶填充在肌原纤维蛋白凝胶网络结构,从而提高凝胶强度[13,14,15]。因此,不溶性膳食纤维如何影响肌原纤维蛋白凝胶特性值得深入研究。【拟解决的关键问题】本试验以改性甘蔗膳食纤维-肌原纤维蛋白为模拟体系,利用石蜡切片和扫描电镜分别观察膳食纤维-蛋白的空间分布和凝胶三维网络结构;利用质构仪研究复合凝胶断裂形变时的应力应变。进而根据以上指标分析改性甘蔗膳食纤维作为脂肪替代物的可行性;建立肌原纤维蛋白凝胶网络结构—膳食纤维-蛋白空间分布—复合凝胶质构三者之间的相互关系,阐释不溶性膳食纤维对肌原纤维蛋白凝胶特性的影响机制。1 材料与方法
试验于2019年在南京农业大学国家肉品质量与安全控制中心实验室进行。1.1 试验材料
猪肉购于苏果超市;甘蔗渣取自于南京苜蓿园农贸市场;过氧化氢、氢氧化钠、盐酸、氯化钠、EGTA、碳酸一氢钠、碳酸二氢钠、氯化镁、戊二醛,购于西格玛试剂有限公司。所有试剂均为分析纯。1.2 仪器与设备
DHG-90338S-Ⅲ烘箱,中国上海沪粤明科学仪器有限公司;pH计,瑞士METTLER公司;HK-10B粉碎机,中国广州市旭朗机械设备有限公司;T25DS25I KA均质机,德国IKA有限公司;IKA磁力搅拌器,德国IKA有限公司;Avanti JXN-30高速冷冻离心机,美国Beckman公司;CM-700d色差计,日本美能达公司;TW20水浴锅,德国Julabo公司;MCR301流变仪,奥地利Anon Paar公司;TA-XT Plus质构仪,英国Stable Micro systems公司;Axio Lab. A1正置荧光显微镜,德国Olympus Optical公司;MM-12绞肉机,中国丰伟机械有限公司;CM1950切片机,德国徕卡公司。1.3 试验方法
1.3.1 甘蔗膳食纤维的制备 参照SANGNARK等[16]的方法制备甘蔗膳食纤维。将取自南京苜蓿园农贸市场的甘蔗渣处理成 0.5 cm左右的小长条,用自来水洗净3遍去除残留的糖分。将50 g的甘蔗渣放置在5 000 mL 1%过氧化氢溶液(pH 11.5)中搅拌12 h后,用6 mol∙L-1的HCl中和并过滤,用清水洗涤2遍去除酸碱中和产生的盐分。将以上过程重复2次。甘蔗膳食纤维放置在50℃的烘箱中烘干直至重量恒定。将烘干的改性甘蔗膳食纤维放置在粉碎机中进行超微粉碎并筛分,过100目筛的标记为100目,剩余通过50目的标记为50目,筛分后改性甘蔗膳食纤维用塑料袋进行封装保存。在对比物化特性的差异时,试验所用的改性甘蔗膳食纤维和未改善性膳食纤维的粒径同为100目。1.3.1.1 膳食纤维成分分析 根据SANGNARK等[16]的方法分析甘蔗粗纤维和甘蔗膳食纤维中纤维素、半纤维素和木质素的组分。
1.3.1.2 色差的测定方法 使用CR-40色差计对压平的甘蔗渣和甘蔗膳食纤维进行测定,标准白色比色板为L*=96.86,a*=-0.15,b*=1.87。每个样品测定5次。其中L*代表亮度值,a*代表红度值,b*代表黄度值。
1.3.1.3 持水力和持油力 参照SANGNARK等[16]的方法测定甘蔗膳食纤维的持水力和持油力。将3 g干燥样品与过量的去离子水充分搅拌混合并静置2 h后,用细丝筛过滤多余的水分,并称重。样品的持水力定义为甘蔗膳食纤维吸水后与吸水前重量之比。将5 g干燥的样品与过量的大豆油充分混合并放置在室温条件下静置2 h,然后将样品离心(1 500×g,10 min)后,用尼龙膜过滤多余的大豆油称重。持油力定义为甘蔗膳食纤维吸油后与吸油前重量之比。
1.3.2 肌原纤维蛋白的提取和复合体系的制备 剔除猪后腿肉中的结缔组织和脂肪,用绞肉机将其绞碎(0.6 mm孔板),封装于双层真空包装袋(PE/尼龙),每袋500 g,储存于-20℃。提取前取出猪后腿肉并放置于0—4℃冷库中解冻至中心温度为0℃。参照HAN等[17]的方法提取肌原纤维蛋白,并稍作改动。具体操作步骤如下:将50 g斩拌后的肉糜和四倍体积的磷酸缓冲溶液(10 mmol∙L-1 NaH2PO4/Na2HPO4,0.1 mmol∙L-1 NaCl,2 mmol∙L-1 MgCl2,1 mmol∙L-1 EGTA,pH 7.0,4℃)混合匀浆(6 000 r/min,30 s)。将匀浆后混合溶液用双层纱布进行过滤后离心(3 000×g,15 min,4℃)取沉淀,重复上述过程两遍,最终沉淀物即为纯肌原纤维蛋白。参照HAN等[17]的方法用双缩脲法测定纯肌原纤维蛋白浓度,双缩脲标准线性回归方程的决定系数R2=0.98。将甘蔗膳食纤维干粉添加到蛋白浓度为50 mg∙mL-1的肌原纤维蛋白溶液(0.6 mol∙L-1 NaCl,pH 7.0)中。根据肌原纤维蛋白溶液质量,甘蔗膳食纤维(50目和100目)添加量分别为1.0%、2.0%和3.0%。其中未添加甘蔗膳食纤维的试验组为对照组;添加1.0%、2.0%和3.0%的50目甘蔗膳食纤维的试验组分别命名T150、T250和T350;添加1.0%、2.0%和3.0%的100目甘蔗膳食纤维的试验组分别命名T100、T200和T300。用玻璃棒搅拌混合体系3 min,以确保甘蔗膳食纤维在肌原纤维蛋白溶液体系中的均匀分布。将部分样品放入80℃的水浴锅中煮制20 min后,置于冷库中冷却至4℃,另一部分样品用于动态流变测定。
1.3.3 动态流变测定 参照JIANG等[14]的方法使用MCR301型流变仪测定蛋白-甘蔗膳食纤维溶液体系动态流变特性。样品均匀涂抹于50 mm不锈钢圆形平板探头之间,间隙为1 mm。根据样品的应力扫描和振荡扫描测定的样品线性粘弹区间,以设定剪切应力为5 Pa,振荡频率为0.1 Hz。测试程序:样品温度保持在20℃,静置5 min后,从20℃升温至80℃,加热速度为2℃∙min-1。在加热过程中,在恒定的应力和扫描频率下对样品进行持续剪切,每个样品测量5次。
1.3.4 复合凝胶的形变特性 参照WU等[13]的方法测定复合凝胶的形变特性。将复合凝胶切割成直径2 cm、高度2 cm的圆柱。使用质构仪测定样品复合凝胶的形变特性,具体参数如下:下降速度2.0 mm∙s-1,测试速度1.0 mm∙s-1,质构探头为P50型号,压缩比为90%。其中,复合凝胶断裂形变时的应变计算公式为:
式中,H0:样品的初始高度,H:样品发生断裂形变时的压缩高度;其中的复合凝胶断裂形变时的应力计算公式为:
式中,F:压缩过程中所测得的力的大小,A:样品接触探头的表面积。
1.3.5 离心损失 参照ZHUANG等[11]的方法测定复合蛋白凝胶的离心损失。取8 g凝胶样品置于10 mL的塑料离心管中,上方塞入吸水纸用于吸水。之后对样品离心(10 000×g,4℃)10 min。持水力即为凝胶样品离心后的重量占离心前重量的百分比。每个样品测定5次。
1.3.6 微观结构观察 将凝胶样品切割成侧面积为1 cm2大小的长方体,并用4%的多聚甲醛进行结构固定。将样品从固定液取出,进行切片和染色后,用正置荧光显微镜放大1 000倍观察切片的微观结构。每个样品取5个不同的观察视野。
将凝胶样品切割成侧面1 cm2大小的长方体,并用2.5%的戊二醛磷酸缓冲溶液固定24 h。固定后用磷酸缓冲溶液对样品进行多次漂洗,用不同浓度酒精对样品脱水。脱水干燥后的样品粘于金属台上进行喷金镀膜。用S-3000N扫描电镜进行放大2 000倍观察,加速电压为20 kV。每个样品分别观察5个视野。参照HAN等[17]的方法对扫描电镜图片进行二维化处理。将扫描电镜图像导入ImageJ v1.47后,利用其Fraclac 2.5 v插件对其进行处理分析。首先,在图像的像素调整到640×432后,对其进行二维化处理。分维分析反映的是不规则复杂结构占有空间的有效性。盒计数法为在图像上定义几个尺寸缩小的网格,并计算每个网格中包含蛋白三维网络结构的像素数量[18]。分形维数基于下列方程计算得出:
式中,Nε:不同尺寸网格中所包含的像素数量,ε:网格的大小尺寸,D:凝胶蛋白三维网络结构的分维维度。缺项分析是反映凝胶三维网络结构中水孔隙的均匀程度[19]。缺项分析定义为在图像中定义N个数量相同大小的网格,计算这些网格中代表水相形成水孔隙目标像素的变异系数。缺项分析基于下列方程计算得出:
式中,Λ:目标像素的缺项,σ:所有大小网格内目标像素数量的标准差,μ:所有大小网格内目标像素数量的平均值。
1.3.7 数据分析 试验设计采用完全随机区组的方法,所有处理重复3次。所有数据用SAS 9.0统计软件进行分析,采用单因素方差分析,比较方法采用Duncan多重比较分析,显著差异性为P<0.05。
2 结果
2.1 碱性过氧化氢处理对膳食纤维物化特性的影响
图1显示为甘蔗膳食纤维经过碱性过氧化氢处理前后的对比图。碱性过氧化氢处理使甘蔗膳食纤维亮度值提高近35%(P<0.05),显著降低其红度值和黄度值。此外,碱性过氧化氢法还可显著提高甘蔗膳食纤维的持水力和持油力(P<0.05)。碱性过氧化氢处理能够松散纤维结构,使更多的亲水基团和疏水基团暴露,使其持水力和持油力显著提高[16]。表1显示碱性过氧化氢处理对甘蔗膳食纤维组分的改变:纤维素含量从45.24%显著增加到54.34%,而木质素的含量从18.20%显著减少到8.84%,表明碱性过氧化氢处理能够降解木质素,从而提高甘蔗膳食纤维中纤维素和半纤维素的比例[16]。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1甘蔗膳食纤维碱性过氧化氢处理前后对比图
A:处理前;B:处理后
Fig. 1Comparison diagram of sugarcane dietary fiber without or with alkaline hydrogen peroxide treatment
A:Before treatment;B:After treatment
Table 1
表1
表1改性前后甘蔗膳食纤维的物化特性
Table 1
| 样品名称 Treatment | 亮度值 L* | 红度值 a* | 黄度值 b* | 持水力 WHC | 持油力 WOC | 纤维素 Cellulose (%) | 半纤维素 Hemicellulose (%) | 木质素 Lignin (%) |
|---|---|---|---|---|---|---|---|---|
| 改性前 Without AHP treatment | 57.89±0.62b | 3.49±0.17a | 22.34±0.09a | 4.90±46b | 3.23±21b | 45.25±2.62b | 27.49±2.17 | 18.20±1.08a |
| 改性后 With AHP treatment | 77.50±0.36a | 0.96±0.11b | 15.20±0.08b | 10.52±21a | 5.56±37a | 53.28±2.36a | 24.33±2.11 | 8.84±1.09b |
新窗口打开|下载CSV
2.2 离心损失
香肠体系中水分含量高达50%,其主要通过毛细管作用力被束缚于肌原纤维蛋白凝胶网络结构中。离心损失能够直接反映肌原纤维蛋白的凝胶程度以及其对体系中水分的束缚能力[20,21]。如表2所示,添加甘蔗膳食纤维能够显著降低复合凝胶体系的离心损失,且随着添加比例提高,离心损失显著下降,说明添加膳食纤维能够促进热诱导凝胶过程中肌原纤维蛋白之间的相互交联,使最终的凝胶结构对水分的束缚能力显著提高。相同添加比例条件下,甘蔗膳食纤维粒径越大,复合凝胶的离心损失越小。但是在甘蔗膳食纤维添加比例为3%时,粒径越小的处理组(T300)离心损失最低(3.45%)。在研究小麦膳食纤维对鱼糜的凝胶强度时也发现,当膳食纤维添加比例为8%时,鱼糜的凝胶强度最大[22]。离心损失反映的是肌原纤维蛋白凝胶结构对水分的束缚能力,间接说明甘蔗膳食纤维能够促进肌原纤维蛋白交联形成致密的三维网络结构,但是在T350组和T300的离心损失无显著差异,这可能达到了添加极限,类似文献报道鱼糜的小麦膳食纤维添加比例极限为8%[22]。Table 2
表2
表2不同添加比例和粒径的甘蔗膳食纤维对复合凝胶离心损失、动态流变特性、分维维度以及缺项值的影响
Table 2
| 样品名称 Treatments | 离心损失 Centrifugal loss (%) | T1 (℃) | T2 (℃) | G'终值 Final G' | 增幅 Amplitude | 分维维度 Fracture dimensions | 缺项值 Lacunary |
|---|---|---|---|---|---|---|---|
| 对照 CK | 9.39±0.25a | 47.9 | 59.9 | 3128±65g | 2028±65g | 1.8117±0.0042e | 0.336±0.017a |
| T150 | 5.78±0.07c | 47.9 | 59.9 | 4423±83e | 3056±45e | 1.8335±0.0073d | 0.26±0.008b |
| T100 | 6.24±0.21b | 48.16±0.37 | 59.9 | 3809±119f | 2543±78f | 1.8278±0.0040d | 0.273±0.005bc |
| T250 | 4.16±0.13e | 47.9 | 59.9 | 6011±95c | 3911±95c | 1.8490±0.0041bc | 0.23±0.008d |
| T200 | 4.69±0.10d | 47.9 | 59.9 | 4856±62d | 3423±48d | 1.8385±0.0052cd | 0.247±0.012cd |
| T350 | 3.78±0.14f | 47.9 | 59.9 | 8445±112a | 5345±112a | 1.8670±0.0056a | 0.19±0.008e |
| T300 | 3.45±0.31f | 48.16±0.37 | 59.9 | 6841±58b | 4174±122b | 1.8555±0.0042ab | 0.227±0.012d |
新窗口打开|下载CSV
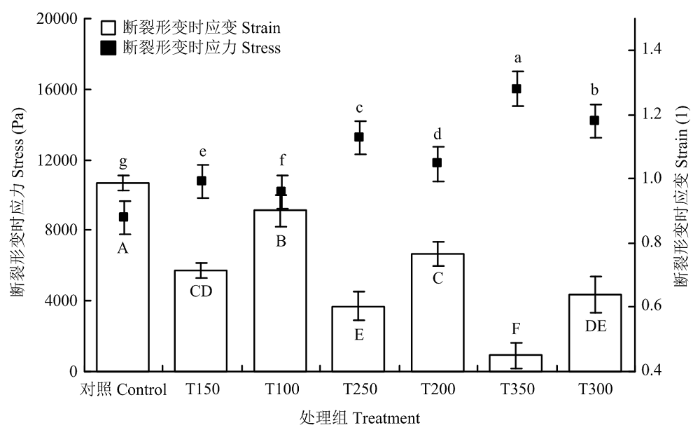
2.3 复合凝胶的形变特性
将质构仪所测定的试验结果通过公式模型转化成复合凝胶在发生断裂形变时的应力应变。断裂形变时的应力能够反映凝胶食品在咀嚼过程中的硬度,其与硬度呈正比关系。断裂形变时的应变能够反映凝胶食品在咀嚼过程中的弹性,其与弹性成正比关系。图2显示膳食纤维添加比例的提高,复合凝胶的断裂形变时应力发生了显著的提高,但是断裂形变时应变却发生了显著的降低。对照组断裂形变时的应力、应变分别为8 718 Pa和0.99,其应力值是所有处理组中最低值,应变值是所有处理组中的最高值。添加3% 50目甘蔗膳食纤维处理组断裂形变时的应力、应变分别为16 024 Pa和0.44,其应力值是所有处理组中的最高值,应变值是所有处理组中的最低值。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同添加比例和粒径的甘蔗膳食纤维对复合凝胶断裂形变时应力应变的影响
不同小写字母表示应力差异显著(P<0.05),不同大写字母表示应变差异显著(P<0.05)
Fig. 2True fracture stress and true fracture strain of composite MP gel influenced by sugarcane dietary fiber with various concentration and particle size
Different lowercase letters indicate significant difference of stress (P<0.05), Different capital letters indicate significant difference of strain (P<0.05)
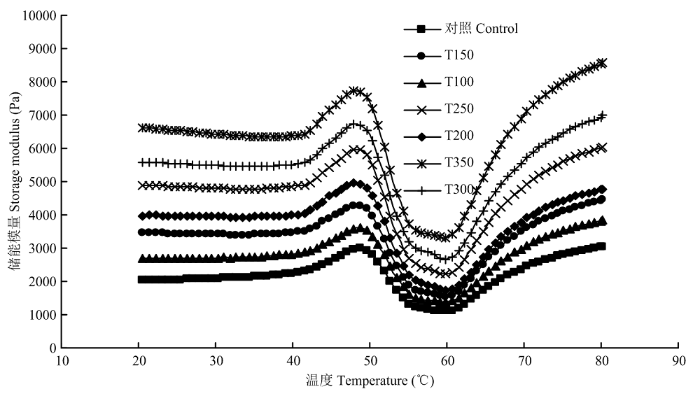
2.4 动态流变特性
曲线G'反映肌原纤维蛋白在升温过程中蛋白构象的折叠和展开以及基团之间的相互作用。曲线G'的变化反映肌原纤维蛋白加热过程中经历的3个主要阶段:1)42℃—48℃,肌球蛋白头部发生交联形成二聚体,导致G'上升;2)54℃—58℃,肌球蛋白尾部链结构解聚,引起G'下降;3)最后阶段为60℃以后,肌球蛋白因解聚而暴露的疏水基团发生交联形成三维网络结构,G'发生急剧上升[1]。曲线G'显示添加不同比例和粒径不溶性甘蔗膳食纤维没有改变肌原纤维蛋白升温过程中3个转变阶段,说明甘蔗膳食纤维没有和肌原纤维蛋白发生共价或者非共价交联,进而改变肌原纤维蛋白构象的折叠展开和基团的相互交联。但是,随着甘蔗膳食纤维比例的添加和粒径增大,复合体系G'数值显著增加(图3)。有关研究证明:凝胶体系的G'的大小和质构的硬度成正相关,说明添加甘蔗膳食纤维能够提高复合凝胶的质构,这和离心损失结果相吻合。尤其处理组(T350)的最终G'值(8 445 Pa)是未添加组(3 128 Pa)的两倍多(表2)。为了进一步比较添加甘蔗膳食纤维对G'在最后形成三维网络结构凝胶阶段的影响,试验对最后阶段的G'急剧上升的幅度进行了量化比较。分析结果发现对照组G'的上升程度为2 028 Pa,添加3% 50目甘蔗膳食纤维处理组G'的上升程度显著增加5 345 Pa。有关研究也发现小麦膳食纤维添加到鱼糜中后,凝胶的G'也发生了显著增加的现象[23]。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同添加比例和粒径甘蔗膳食纤维对肌原纤维蛋白热凝胶过程中储能模量的影响
Fig. 3Changes in storage modulus G′ of MP emulsions with various concentrations and particle sizes of SDF
2.5 微观结构观察
石蜡切片清晰地显示不同添加比例和粒径的甘蔗膳食纤维在肌原纤维蛋白凝胶网络结构中的空间分布,其中红色部分是染色后的肌原纤维蛋白,白色部分中蜂窝状结构的是甘蔗膳食纤维。图4显示甘蔗膳食纤维和肌原纤维蛋白之间没有发生交联。膳食纤维的中空结构类似于一个海绵体,能够吸附大量的水分,简单的以物理填充的形式镶嵌在肌原纤维蛋白凝胶结构中。石蜡切片显示对照组(纯蛋白组)结构致密且分布均一。添加甘蔗膳食纤维后,结构致密且分布均一的肌原纤维蛋白凝胶网络结构中会形成大量的大小和形状各异的空洞。随着添加比例和粒径的增大,空洞的数量和体积也显著增加。石蜡切片显示添加膳食纤维形成的空洞结构会降低肌原纤维蛋白凝胶结构的均一性。此外,大量文献发现多糖并不能和肌原纤维蛋白发生交联,通常通过物理镶嵌形式填充在蛋白凝胶网络结构中[24,25]。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4甘蔗膳食纤维在蛋白凝胶中的空间分布
A:对照组;B:T150;C:T100;D:T250;E:T200;F:T350;G:T300。下同
Fig. 4The spatial distribution of SDF in the MP gels
A:Control;B:T150;C:T100;D:T250;E:T200;F:T350;G:T300。The same as below
扫描电镜清晰地显示了甘蔗膳食纤维对肌原纤维蛋白凝胶三维网络的影响变化。为了量化肌原纤维蛋白凝胶三维网络的变化,本试验运用图像分析软件对二维化后的扫描电镜图片进行分维分析、缺项分析(表2)。如图5中的白色部分为肌原纤维蛋白凝胶网络结构,黑色部分为水空隙(或水沟壑)。对照组图片显示:肌原纤维蛋白凝胶网络结构中存在大量的相互交联的水沟壑,将肌原纤维蛋白网络结构分割成片状化(图5)。此外,对照组的肌原纤维蛋白呈粗丝状甚至部分区域呈现团状结构,这说明在加热过程中这些肌原纤维蛋白的折叠结构在加热过程中没有充分的交联并未形成致密的三维网络结构。在溶液状态下,肌原纤维蛋白盐溶过程中吸附大量的水分,随着温度升高,肌原纤维蛋白尾部链结构展开并暴露大量疏水基团,同时伴随着部分吸附的水分渗出,渗出的水分会阻碍后续蛋白疏水基团之间的交联作用,导致整个肌原纤维蛋白三维网络结构的疏松[26,27]。图像处理软件分析结果显示对照组肌原纤维蛋白凝胶网络结构的分维维度(1.8117)显著小于其他添加甘蔗膳食纤维组(P<0.05),而缺项值(0.336)显著高于其他添加甘蔗膳食纤维组(P<0.05)(表2)。分维维度越小,反映蛋白凝胶网络结构之间的相互交联作用弱,结构比较松散。缺项值越小说明蛋白凝胶网络结构中的水孔隙分布均匀且大小相似,间接地反映肌原纤维蛋白三维网络结构的致密度高。
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5甘蔗膳食纤维对肌原纤维蛋白凝胶微观结构(左)和二维化微观结构(右)的影响
Fig. 5Scanning electron micrographs (left) of gel with various IDF addition and their corresponding binarized images (right)
随着甘蔗膳食纤维添加比例的提高,肌原纤维蛋白凝胶三维网络结构发生了显著变化。从肌原纤维蛋白三维网络结构角度看,肌原纤维蛋白凝胶三维网络结构中粗丝状和团状结构完全消失了,取而代之的是相互交联的致密细丝网状结构,尤其处理组G。从水孔隙(或者水沟壑)角度看,随着甘蔗膳食纤维的添加比例和粒径提高,肌原纤维蛋白三维网络结构中水孔隙的数量显著减少,并且彼此之间也没有相互交联。当甘蔗膳食纤维添加量达到3%时,肌原纤维蛋白三维网络结构中相互交叉的水孔隙基本消失,只有数量不多且体积小的水孔隙均匀地镶嵌在致密的蛋白网络结构中。图像处理软件分析结果显示:T350处理组蛋白凝胶网络结构的分维维度(1.867)为所有处理组中最高(P<0.05),而缺项值(0.19)为所有处理组中最低(P<0.05)。
3 讨论
低脂化是为了凝胶肉制品的发展趋势,然而脂肪替代物的添加通常会改变原有的凝胶结构,导致产品口感劣变而不被消费者所接受[28,29]。通过碱性过氧化氢法对甘蔗膳食纤维进行改性,提高其亮度值、持水力和持油力等物化特性。将膳食纤维添加到肌原纤维蛋白凝胶体系中发现,复合凝胶的断裂应力和动态流变特性都得到了显著的提高,证明甘蔗膳食纤维作为优质脂肪替代物的可行性。肌原纤维蛋白凝胶网络结构—膳食纤维-蛋白空间分布—复合凝胶质构三者之间的相互关系如图6所示。断裂形变时应力反映复合凝胶的硬度,应力值与硬度成正比;断裂形变时应变反映复合凝胶的弹性,应变与弹性成正比[30]。甘蔗膳食纤维能够稳定水相,减少热诱导凝胶过程中蛋白网络结构内部水沟壑的形成,间接地促进肌原纤维蛋白之间的交联,形成致密的均一的网络结构。膳食纤维的添加比例和粒径与水相稳定能力成正比,例如:T350处理组具有最高的复合凝胶硬度。然而,吸水后的甘蔗膳食纤维只是简单的物理镶嵌在肌原纤维蛋白三维凝胶网络中,并且形成大小和形状各异的空洞。在复合凝胶网络结构中,肌原纤维蛋白凝胶网络是弹性结构,膳食纤维结构是中空松散结构,随着膳食纤维添加比例和粒径的提高,导致最终复合凝胶的弹性显著变差,例如:T350处理组具有最差的复合凝胶弹性。因此,膳食纤维通过稳定水相间接促进肌原纤维蛋白之间的交联,从而提高复合凝胶的硬度。但是膳食纤维本身是非弹性结构,简单的物理镶嵌在蛋白凝胶网络中形成大量的空穴,会显著降低复合凝胶的弹性。使用小粒径的甘蔗膳食纤维作为脂肪替代物能够显著提高复合凝胶的凝胶强度,同时极大限度地保持复合凝胶的弹性,研究结果为甘蔗膳食纤维在低脂营养香肠中的运用提供了理论指导。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6肌原纤维蛋白凝胶网络结构—膳食纤维-蛋白空间分布—复合凝胶质构三者之间的相互关系
Fig. 6The relationship between the MP gel networks, SDF- MP spatial distribution and the textural property of composite gel
4 结论
改性甘蔗膳食纤维显著影响复合凝胶的质构品质,主要表现在两个方面:一是提高了复合凝胶断裂形变时的应力,T350组值最高(14 159 Pa);二是降低了复合凝胶的断裂形变时的应变,对照组值最高(0.99)。膳食纤维在肌原纤维蛋白凝胶的空间分布显示,吸水后的甘蔗膳食纤维只是简单的物理镶嵌在肌原纤维蛋白三维凝胶网络中,并且形成大小和形状各异的空洞。肌原纤维蛋白凝胶微观结构显示甘蔗膳食纤维能够稳定水相,减少热诱导凝胶过程中蛋白网络结构内部水沟壑的形成,间接地促进肌原纤维蛋白之间的交联,形成致密的均一的网络结构。本研究基于对蛋白骨架结构—膳食纤维-蛋白填充结构—凝胶质构之间的相互关系,揭示了甘蔗膳食纤维改善肌原纤维蛋白凝胶特性的机制。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.meatsci.2004.11.021URL [本文引用: 2]
DOI:10.3945/ajcn.2008.26285URL [本文引用: 1]
DOI:10.1007/s13197-013-1010-2URL [本文引用: 2]
DOI:10.1016/S0308-8146(98)00076-4URL [本文引用: 1]
DOI:10.1080/10408398.2012.682230URL [本文引用: 3]
DOI:10.1016/j.foodres.2011.07.003URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.foodchem.2008.05.054URL [本文引用: 1]
DOI:10.1016/j.meatsci.2016.02.035URL [本文引用: 2]
DOI:10.1016/j.meatsci.2012.11.025URL [本文引用: 1]
DOI:10.1016/j.foodres.2017.07.063URL [本文引用: 2]
DOI:10.1016/j.lwt.2018.06.053URL [本文引用: 1]
DOI:10.1016/j.foodchem.2015.01.125URL [本文引用: 3]
[本文引用: 3]
DOI:10.1016/j.foodchem.2016.09.068URL [本文引用: 2]
DOI:10.1016/S0308-8146(02)00257-1URL [本文引用: 5]
DOI:10.1016/j.foodres.2014.05.062URL [本文引用: 3]
DOI:10.1021/jf970348mURL [本文引用: 1]
DOI:10.1259/dmfr/85149245URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.foodchem.2013.10.010URL [本文引用: 2]
DOI:10.1016/j.foodchem.2013.02.111URL [本文引用: 1]
DOI:10.1016/j.foodhyd.2014.11.021URL [本文引用: 1]
DOI:10.1016/S0268-005X(00)00025-4URL [本文引用: 1]
DOI:10.1038/s41598-017-11711-1URL [本文引用: 1]
DOI:10.1016/j.foodhyd.2016.04.014URL [本文引用: 1]
DOI:10.1016/j.meatsci.2016.01.002URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.foodhyd.2016.04.007URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}