 ,1, 赵旭东2, 姜义平1, 赵静1, 肖留斌,1, 郝德君,2
,1, 赵旭东2, 姜义平1, 赵静1, 肖留斌,1, 郝德君,2Cloning, Preparation of Antibody and Response Induced by 20-Hydroxyecdysone of Target of Rapamycin in Apolygus lucorum
TAN YongAn,1, ZHAO XuDong2, JIANG YiPing1, ZHAO Jing1, XIAO LiuBin,1, HAO DeJun,2通讯作者:
责任编辑: 岳梅
收稿日期:2020-07-31接受日期:2020-09-7网络出版日期:2021-05-16
| 基金资助: |
Received:2020-07-31Accepted:2020-09-7Online:2021-05-16
作者简介 About authors
谭永安,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1775KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
谭永安, 赵旭东, 姜义平, 赵静, 肖留斌, 郝德君. 绿盲蝽雷帕霉素靶蛋白的克隆、抗体制备及在蜕皮激素诱导下的应答[J]. 中国农业科学, 2021, 54(10): 2118-2131 doi:10.3864/j.issn.0578-1752.2021.10.008
TAN YongAn, ZHAO XuDong, JIANG YiPing, ZHAO Jing, XIAO LiuBin, HAO DeJun.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】绿盲蝽(Apolygus lucorum)属半翅目(Hemiptera)盲蝽科(Miridae),寄主范围广泛,可危害棉花等多种经济作物[1,2]。20世纪90年代末以来,由于Bt棉的大面积推广种植和农业产业结构的调整,绿盲蝽种群数量急剧上升,已成为棉田主要害虫,并迅速波及枣树、葡萄、樱桃、桃等多种果树,严重影响其产量及品质。生产上对化学农药的长期依赖势必会导致绿盲蝽的抗药性日渐增强,从而造成防治效果下降[3,4]。因此,目前绿盲蝽防控面临严峻挑战,亟需开拓基于新靶标的有效防控技术。【前人研究进展】雷帕霉素靶蛋白 (target of rapamycin,TOR)是一种丝氨酸/苏氨酸激酶,在细胞代谢、生长及增殖方面至关重要[5,6]。TOR可分为TOR复合物1(TOR complex 1,TORC1)和TOR复合物2(TOR complex 2,TORC2)。TORC1是TOR与RapTOR(regulatory associated protein of mTOR)及mLST8(mammalian lethal with Sec13 protein 8)形成的一个对雷帕霉素敏感的复合物,可调控细胞内蛋白质翻译、细胞凋亡及细胞周期等关键生理过程[7,8]。TORC2是由RicTOR(rapamycin insensitive cornpanion of mTOR)、Sinl(也称为Mip1)与mLST8、mTOR共同形成的对雷帕霉素不敏感的复合物,通过调控蛋白激酶B(protein kinase B,PKB)和蛋白激酶C(protein kinase C,PKC)的活化,主要参与细胞极性、空间及骨架结构的形成[9]。在昆虫中,TOR可通过直接耦合前胸腺中的营养感应,从而在蜕皮激素(20E)生成及蜕皮过程中发挥重要作用[10,11]。研究表明,抑制TOR的表达可造成幼虫发育历期延长,同时与蜕皮激素合成相关基因的表达量下调,表明昆虫在生长发育以及蜕皮变态时需要TOR的参与;此外,在幼虫营养缺乏的条件下,TOR信号的激活会诱导蜕皮激素的生成,使幼虫完成正常蜕皮及变态行为[12]。TOR信号也可受蜕皮激素的调控[13,14]。对黑腹果蝇(Drosophila melanogaster)的研究发现,当幼虫摄入营养时,蜕皮激素抑制TOR信号介导调控胰岛素样肽(ILP)生成胰岛素分泌细胞来调控果蝇的生长发育[15,16]。【本研究切入点】目前对于TOR的研究多集中在植物、哺乳动物及一些模式昆虫上[17,18,19],对绿盲蝽AlTOR的相关研究尚未见报道。【拟解决的关键问题】采用RACE技术从绿盲蝽中扩增AlTOR全长,运用生物信息学方法对该基因进行结构分析和同源进化分析,并利用qRT-PCR阐明其时空表达动态及20E对其表达的影响,同时将其在原核细胞中进行高效表达,筛选最佳表达条件,以获得纯化蛋白,最后制备特异性的抗体,为在蛋白水平进一步研究该基因的功能打下基础。1 材料与方法
试验于2018年9月至2019年12月在江苏省农业科学院植物保护研究所完成。1.1 供试昆虫
绿盲蝽于2018年采自江苏沿海棉区的大丰市蚕豆田(33.11°N,120.25°E),室内用新鲜四季豆(Phaseolus vulgaris)续代饲养,饲养条件为温度(27±0.5)℃,相对湿度(70±5)%,光周期14L:10D。1.2 主要试剂及仪器
RNA提取试剂盒(Promega),M-MLV反转录试剂盒(Promega),BD SMARTTM RACE cDNA Amplification Kit(宝生物工程(大连)有限公司),pCzn1表达载体、PFU DNA聚合酶(南京钟鼎生物技术有限公司),TOP10菌株(天根生化科技有限公司),大肠杆菌(Escherichia coli)Arctic Express(DE3)(Agilent);卡那霉素、咪唑、尿素(上海生工生物工程有限公司);限制性内切酶(TaKaRa),Protein Marker(Thermo),IPTG、Acr、Bis、Tris(Sigma),SDS(Amresco),TEMED(BIO-RAD),高糖、Tyrptone、Yeast Extract(OXOID),Agarose(上海赛百盛基因技术有限公司);其他试剂均为国产分析纯。Ni2+ IDA亲和层析胶(Novagen),HexIOS分光光度计、Allegra 21R台式高速冷冻离心机(Beckman),台式高速离心机(Sorval),Biologic LP层析系统、Mini Protean II垂直平板电泳系统、水平电泳系统(BIO-RAD),PTC-200基因扩增仪(MJ Research),20E、U73211(Sigma)。1.3 AlTOR全长基因的克隆
基于实验室构建的绿盲蝽转录组数据及已报道昆虫TOR进行BLAST搜索,筛选绿盲蝽转录组中可能的TOR基因片段序列,从而进行克隆验证。根据获得的序列设计特异性引物(表1),进行AlTOR 5′和3′端序列的克隆。取绿盲蝽3龄若虫20头,Trizol法提取总RNA后合成cDNA第1链,根据设计的特异性上游引物和下游引物,进行AlTOR 5′和3′端序列的克隆。5′端序列的克隆采用巢式PCR,第一轮反应体系:10×PCR缓冲液5.0 μL、25 mmol·L-1 MgCl2 3.0 μL、10 mmol·L-1 dNTP Mix 1.0 μL、10 μmol·L-1 5′-AlTOR-1 2.0 μL、10 μmol·L-1通用引物Abridged Anchor Primer 2.0 μL、cDNA 5.0 μL、Taq DNA聚合酶0.5 U,超纯水补足至50.0 μL;第二轮反应体系:10×PCR缓冲液5.0 μL、25 mmol·L-1 MgCl2 3.0 μL、10 mmol·L-1 dNTP Mix 1.0 μL、10 μmol·L-1 5′-AlTOR-2 1.0 μL、10 μmol·L-1通用引物Abridged Universal Amplification Primer 1.0 μL、第一轮PCR产物5.0 μL、Taq DNA聚合酶0.5 U,超纯水补足至50.0 μL;PCR反应条件:预变性94℃ 2 min,94℃ 30 s,55℃ 30 s,72℃ 1 min,循环35次,72℃延伸7 min。3′端序列的克隆也采用巢式PCR,第一轮反应体系:PCR-Grade Water 34.5 μL,10×Advantage 2 PCR Buffer 5.0 μL,10 mmol·L-1 dNTP Mix 1.0 μL,50×Advantage 2 Polymerase Mix 1.0 μL,cDNA 2.5 μL,10 μmol·L-1 3′-AlTOR-1 1.0 μL,10×UPM 5.0 μL,超纯水补足至50.0 μL;第二轮反应体系:PCR-Grade Water 34.5 μL,10×Advantage 2 PCR Buffer 5.0 μL,10 mmol·L-1 dNTP Mix 1.0 μL,50×Advantage 2 Polymerase Mix 1.0 μL,第一轮PCR产物2.5 μL,10 μmol·L-1 3′-AlTOR-2 1.0 μL,10×UPM 5.0 μL,超纯水补足至50.0 μL;PCR反应条件:预变性94℃ 2 min,94℃ 30 s,68℃ 30 s,72℃ 1 min,循环30次,72℃延伸7 min。将获得的5′和3′端及保守区域进行拼接和测序(上海生工生物工程有限公司),最终获得AlTOR全长。最后根据测序结果,设计全长基因克隆引物(AlTOR-F和AlTOR-R),进行AlTOR全长基因的克隆。Table 1
表1
表1本研究所用的引物
Table 1
| 引物名称/目的 Primer name/Purpose | 序列 Primer sequence (5′ to 3′) |
|---|---|
| 克隆Cloning | |
| 5′-AlTOR-1 | AACGACTGGGCAAGAA |
| 5′-AlTOR-2 | CAAGACGTTTGGGTAGCA |
| 3′-AlTOR-1 | AGAGACTCAGTCGCTCACTATCAAGA |
| 3′-AlTOR-2 | CATTGAGACCACACCCCAGACACA |
| AlTOR-F | ATGGTGGAAAAATATGCTCG |
| AlTOR-R | GCTAGCCAGTATACAACG |
| 表达谱分析qRT-PCR | |
| RT-AlTOR-F | GGGGTCGTCAACTGGCAGAA |
| RT-AlTOR-R | CCTCCTTCTTCTTGGGTGCGA |
| Al-β-Actin-F | ACCTGTACGCCAACACCGT |
| Al-β-Actin-R | TGGAGAGAGAGGCGAGGAT |
新窗口打开|下载CSV
1.4 AlTOR时空表达定量检测
收集绿盲蝽不同日龄若虫及羽化7 d后雌成虫的不同组织(头、胸、翅、足、中肠、卵巢和脂肪体)。其中不同日龄样本每个处理15头,不同组织样本每个处理10头雌成虫,并设计3个生物学重复,每个生物学重复设置3个技术重复。液氮处理样品后-80℃冰箱保存备用。Trizol法提取总RNA,Prime ScriptTM RT Master Mix反转录合成cDNA,-20℃保存。qRT-PCR试验步骤按照SYBR Premix Ex Taq Kit说明书上进行。所用的AlTOR引物参见表1,并以绿盲蝽持家基因β-actin为内标对照基因[20](GenBank登录号:JN616391)。扩增体系:UltraSYBR Mix 12.5 μL,上下游引物各0.5 μL,cDNA模板2 μL,ddH2O 9.5 μL;扩增条件:95℃预变性5 min,然后95℃ 10 s,60℃ 20 s,72℃ 20 s,循环40次。1.5 20E处理绿盲蝽若虫
分别用20E(2 μmol·L-1)、U73122(20E抑制剂[21],50 μmol·L-1)、20E+U73122处理、乙醇(CK)浸泡切除两端的四季豆1 min,晾干后饲喂初孵1龄若虫。每处理绿盲蝽400头,重复4次。按1.4方法收集绿盲蝽不同发育阶段和不同组织样本后,分析不同处理AlTOR表达量。1.6 AlTOR序列分析与进化树分析
AlTOR核苷酸序列分析使用Expasy在线程序(1.7 AlTOR在大肠杆菌中的表达
使用DNASTAR-Protean软件分析AlTOR蛋白Antigenic index,选取抗原性较好的区域(72—381,蛋白质理论分子量为35.83 kD)作为免疫原区域,使用原核表达获得抗原蛋白。基于PAS(PCR-based accurate synthesis)的方法,在引物的两端各设计保护性碱基合成基因6731-2S,将其连入pCzn1重组质粒用限制性内切酶EcoR I及Xho I双酶切后进行连接。酶切体系:两种限制性内切酶各0.25 μL,10×Buffer缓冲液1.0 μL,带有合适酶切位点的基因片段3 μL,用ddH2O补足至10 μL,37℃水浴3 h。连接体系:酶切回收后的载体5.0 μL,基因片段10.0 μL,T4 DNA连接酶1.0 μL,10×Buffer缓冲液2.0 μL,用ddH2O补足至20 μL,16℃反应12—16 h。连接产物转化大肠杆菌TOP10后采用PCR筛选阳性克隆。将鉴定正确的重组质粒pCzn1-AlTOR转化到大肠杆菌Arctic Express感受态细胞,具体方法:将质粒1 μL加入100 μL感受态细菌中,置冰上20 min;42℃热激90 s,迅速置冰中5 min,加入600 μL LB培养液;37℃,220 r/min振摇1 h,离心后全部涂布于含50 μg·mL-1 Amp的LB平板,37℃倒置培养过夜。挑取转化平板上的多克隆接种于含50 μg·mL-1 Amp的3 mL LB培养液的试管中,37℃ 220 r/min振摇过夜。次日按1:100接种于50 μg·mL-1 Amp的30 mL LB培养液中,37℃ 220 r/min 振摇至菌体OD600为0.6—0.8。取出1 mL培养物,10 000 r/min室温离心2 min,弃上清,用100 μL 1×上样缓冲液重悬菌体沉淀。向剩余的培养物中加入IPTG至终浓度为0.5 mmol·L-1,20℃ 220 r/min振摇过夜,诱导融合蛋白表达。取出1 mL培养物,10 000 r/min室温离心2 min,弃上清,用100 μL 1×上样缓冲液重悬菌体沉淀。剩余培养物4 000 r/min,离心10 min,弃上清,用PBS重悬菌体沉淀;重悬液进行超声波破碎后,分别取上清液与沉淀液加入上样缓冲液重悬。12% SDS-PAGE检测分析,考马斯亮蓝染色显带。
1.8 包涵体蛋白的变性和复性
先将菌体沉淀重悬于20 mL裂解液(20 mmol·L-1 Tris-HCl containing 1 mmol·L-1 PMSF,pH 8.0)中,超声破碎(功率400 W,工作4 s,间歇8 s,共20 min),收集沉淀。使用包涵体洗涤液(20 mmol·L-1 Tris,1 mmol·L-1 EDTA,2 mol·L-1尿素,1 mol·L-1 NaCl,1% Triton X-100,pH 8.0)洗涤包涵体3次。再使用溶解缓冲液(20 mmol·L-1 Tris,5 mmol·L-1 DTT,8 mol·L-1尿素,pH 8.0),溶解包涵体,4℃过夜。室温,10 000 r/min离心15 min,将上述溶液滴加缓冲液(20 mmol·L-1 Tris-HCl,0.15 mol·L-1 NaCl,pH 8.0),逐步成倍梯度稀释缓慢搅拌,将蛋白溶液装入透析袋于上述缓冲液中透析过夜。1.9 融合蛋白的Ni柱亲和纯化
利用低压层析系统,包涵体溶液以0.5 mL·min-1流速上样至Ni-IDA Binding-Buffer预平衡的Ni-IDA -Sepharose CL-6B亲和层析柱,Ni-IDA Binding-Buffer以0.5 mL·min-1流速冲洗,至流出液OD280值到达基线,再用Ni-IDA Washing-Buffer(20 mmol·L-1 Tris-HCl,20 mmol·L-1咪唑,0.15 mol·L-1 NaCl,pH 8.0) 以1 mL·min-1流速冲洗,至流出液OD280值到达基线;用Ni-IDA Elution-Buffer(20 mmol·L-1 Tris-HCl,250 mmol·L-1咪唑,0.15 mol·L-1 NaCl,pH 8.0)以1 mL·min-1流速洗脱目的蛋白,收集流出液;上述收集的蛋白溶液加入透析袋中,使用Ni-IDA Elution-Buffer进行透析过夜;12% SDS-PAGE分析后利用BSA蛋白定量试剂盒测定蛋白浓度。1.10 AlTOR多克隆抗体的制备
取纯化的AlTOR重组蛋白免疫新西兰白兔 (江苏省农业科学院兽医研究所提供)。皮下注射,每次免疫量为400 μg,2—3周免疫一次,共免疫4次。第4次免疫后在大白兔颈动脉采血,8 000 r/min离心8 min所得上清制备抗血清并准备纯化抗体。TOR蛋白与琼脂糖介质偶联制备成抗原亲和纯化层析柱,将PBS与所得抗血清等量混合后缓慢上样,待抗体抗原结合后用甘氨酸洗脱缓冲液洗脱得到所需纯化抗体,并在PBS缓冲液 (pH 7.4)中进行透析过夜(4℃),隔日进行效价测定,BCA浓度测定试剂盒对所得抗体进行浓度测定。1.11 效价测定
间接ELISA法对抗体进行效价测定。以纯化的AlTOR重组蛋白(5 g·mL-1)包被ELISA板,每孔加入100 mL,4℃过夜。用PBST(0.2 g KCl,0.2 g KH2PO4,2.9 g NaHPO4·12H2O,8 g NaC1,0.05% Tween-20,pH 7.4)洗涤3次,加入PBST封闭液(含5%牛血清白蛋白)于37℃封闭2 h。取出酶标板,弃去内液,用PBST洗板3次,加入用PBST连续倍比稀释的AlTOR多克隆抗体和阴性血清每孔100 μL,37℃恒温箱1 h。同上洗涤后,加入100 μL的用PBST以1:5 000稀释的二抗HRP-IgG,37℃放置1 h。用PBST洗板3次,每次5 min,最后加100 μL TMB底物液显色,37℃显色15 min,待终止反应后,酶标仪测定A450值。1.12 AlTOR多克隆抗体的鉴定
检测多克隆抗体的特异性Western blot测定按照SONG等[22]的方法进行并稍作修改。将原核表达的AlTOR重组蛋白及3龄绿盲蝽若虫总蛋白进行SDS-PAGE分析。转印PVDF膜后封闭,一抗为上述制备的蛋白抗体,二抗为辣根过氧化物酶标记的兔抗鼠HRP。1.13 数据分析
不同处理的AlTOR相对表达量变化以2-ΔΔCt[23]计算。差异显著性采用统计软件SAS 8.0中的Duncan氏新复极差法进行分析。2 结果
2.1 AlTOR的克隆及序列分析
利用绿盲蝽转录组筛选AlTOR基因序列,再利用RACE方法,分别进行5′和3′末端的扩增,获得了AlTOR的全长序列(图1)。序列分析结果表明,AlTOR全长1 652 bp,包括一个208 bp的5′非翻译区以及136 bp的3′端非翻译区。AlTOR开放阅读框含有1 308 bp的核苷酸,编码435个氨基酸,理论等电点预测为7.16。结构域分析表明,AlTOR基因翻译后的氨基酸序列具有典型的TOR特征,含有N-末端的SIN1结构域(288 bp)、高度保守的CRIM结构域(390 bp)及脂质与膜结合相关C-末端的PH域(363 bp)(图2)。图1
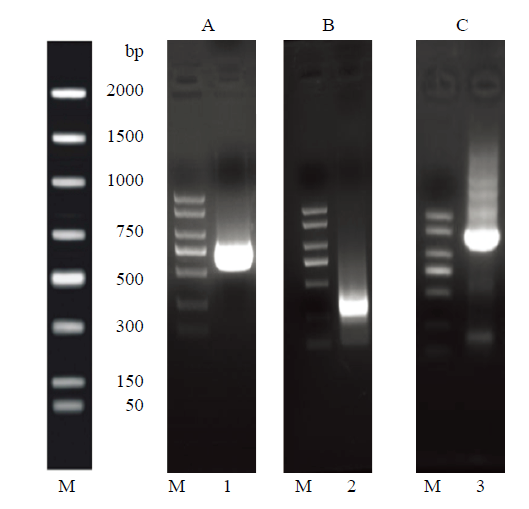 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1AlTOR的5′、3′端及全长序列PCR产物
A:5′端5′ end;B:3′端3′ end;C:全长Full sequence。M:DL2000;1:5′端第二轮PCR产物The second round of 5′ PCR product;2:3′端第二轮PCR产物The second round of 3′ PCR product;3:全长PCR产物Full sequence of PCR product
Fig. 1PCR products of 5′, 3′ end and full sequences from AlTOR
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2AlTOR核苷酸序列及推导的氨基酸序列
黄色阴影代表AlTOR N-末端的SIN1结构域;灰色阴影代表高度保守的CRIM结构域;红色阴影代表脂质与膜结合相关C-末端的PH域
Fig. 2Nucleotide and deduced amino acid sequences of AlTOR
Yellow shaded amino acids represent SIN1 domain of N-terminus, grey shaded amino acids represent highly conserved CRIM domain, and red shaded amino acids represent PH domain of the C-terminus associated with lipid-membrane binding
2.2 多重联比及进化树分析
多重联比结果如图3所示,TOR在半翅目昆虫中较为保守,尤其是CRIM结构域,AlTOR的氨基酸序列与半翅目臭虫科温带臭虫(Cimex lectularius)以及蝽科茶翅蝽(Halyomorpha halys)的TOR氨基酸序列一致性相对较高,分别为55.78%和54.38%,与其他昆虫的TOR基因序列一致性较低,如膜翅目的造纸胡蜂(Polistes dominula)(38.70%)及芜菁叶蜂(Athalia rosae)(39.33%)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3AlTOR与其他半翅目昆虫TOR氨基酸序列多重比对
TOR来源物种及GenBank登录号Origin species of TORs and their GenBank accession numbers:温带臭虫Cimex lectularius (XP_014251577);茶翅蝽Halyomorpha halys (XP_014272645);麦双尾蚜Diuraphis noxia (XP_015365434);棉蚜Aphis gossypii (XP_027843790)。红色方框为AlTOR N-末端的SIN1结构域;蓝色方框为高度保守的CRIM结构域;黄色方框为脂质与膜结合相关C-末端的PH域Red boxes represent SIN1 domain of N-terminus, blue boxes represent highly conserved CRIM domain, and yellow boxes represent PH domain of the C-terminus associated with lipid-membrane binding
Fig. 3Amino acid sequence alignment of AlTOR and other Hemiptera insects
利用MEGA 6.0软件NJ法构建了包括绿盲蝽在内的5个目20种昆虫TOR的系统进化树(图4)。结果显示,TOR作为高度保守的蛋白,在种间同源性较高;外聚群中,膜翅目、鳞翅目、蜚蠊目、缨翅目中聚类更为接近,属于同一类分支。半翅目中,绿盲蝽与温带臭虫、茶翅蝽同源性较高,亲缘关系较为接近;与蚜虫位于不同分支,同源性远低于同为半翅目的温带臭虫和茶翅蝽。
图4
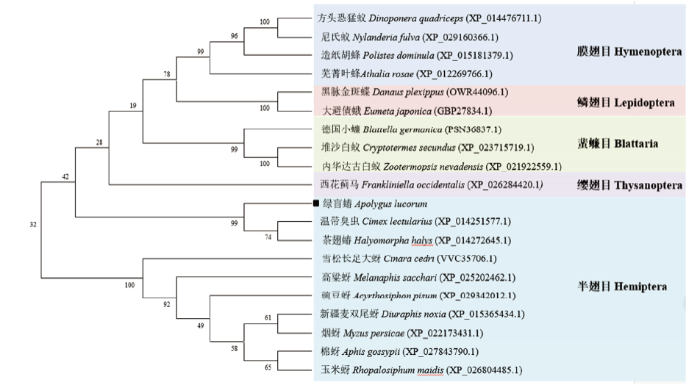 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4AlTROR与其他已知昆虫TOR氨基酸序列的系统进化树
采用Bootstrap方法分析,每个分支进行1 000次验证,数值代表Bootstrap估算值
Fig. 4Phylogenetic tree based on amino acid sequence of AlTOR and other insects
Using the Bootstrap method for analysis, 1 000 bootstrap replicates and values representing the Bootstrap estimate
2.3 AlTOR在绿盲蝽不同发育阶段及组织中的表达
AlTOR在绿盲蝽1—16日龄虫体中均有表达,表达量呈现波动式下降趋势,其中在初孵若虫(1日龄)表达量最高,8日龄若虫表达量最低,此外,AlTOR在绿盲蝽各龄期蜕皮时表达量显著升高(图5)。AlTOR在绿盲蝽雌成虫各组织中均有表达,其中在卵巢、中肠和脂肪体中高表达,而在头、胸、足中表达量较低,AlTOR的表达具有组织特异性(图6)。图5
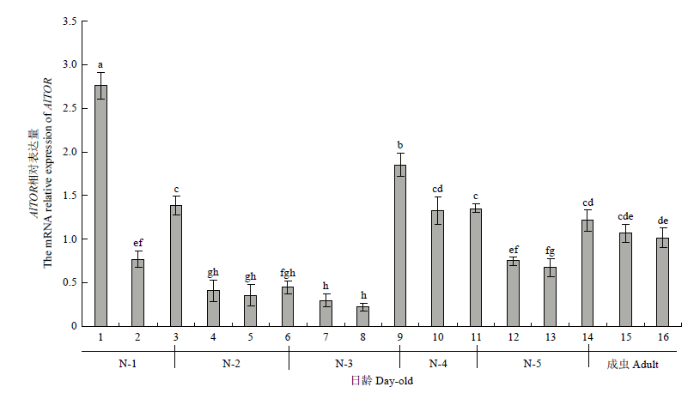 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5AlTOR在绿盲蝽不同发育阶段的表达
图中数据为平均数±标准误,柱上不同字母表示差异显著(P<0.05)。下同
Fig. 5Relative expression levels of AlTOR at different stages of A. lucorum
Date in the figure are means±SE, and different letters above bars indicate significant difference (P<0.05). The same as below
图6
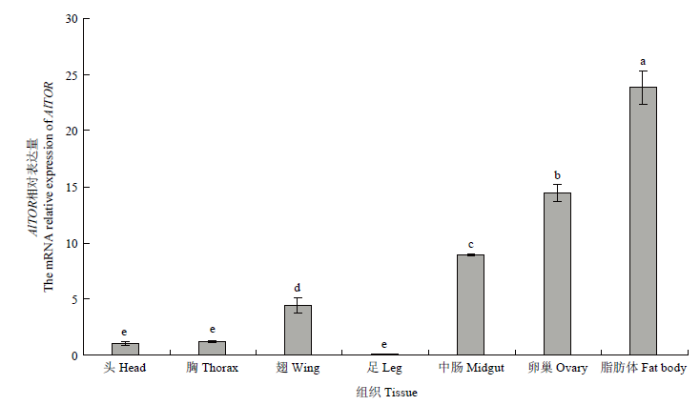 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6AlTOR在绿盲蝽雌成虫不同组织中的表达
Fig. 6Relative expression levels of AlTOR in different tissues of A. lucorum female adult
2.4 不同处理对AlTOR相对表达量的影响
与CK(乙醇)相比,当绿盲蝽分别发育至1龄蜕皮及2龄蜕皮时(4 d和6 d),20E显著上调AlTOR的表达;U73122处理后,AlTOR的表达呈现先上升后下降的趋势;此外,20E+U73122混合处理后,AlTOR的表达总体变化不大(图7)。与CK(乙醇)相比,20E处理绿盲蝽雌成虫后,各组织中AlTOR表达均显著上调,其中头、翅、卵巢和脂肪体中基因表达量分别显著提高了200.00%、118.89%、20.53%和60.82%;而当U73122处理后,绿盲蝽雌成虫的头、翅和足中AlTOR显著上调,中肠、卵巢和脂肪体中AlTOR表达量则下调(图8)。图7
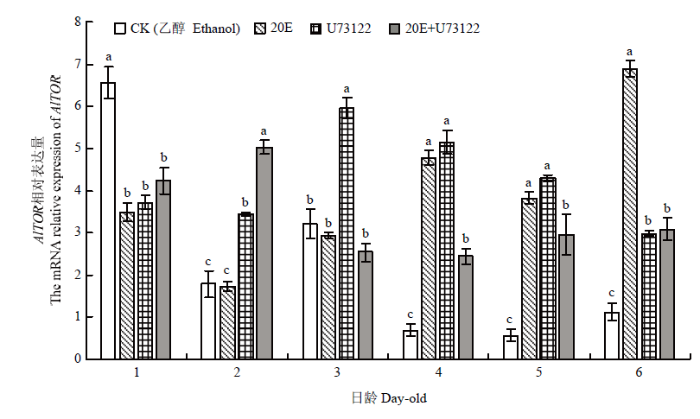 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同处理下各日龄绿盲蝽AlTOR的表达
Fig. 7Relative expression levels of AlTOR at different stages of A. lucorum under different treatments
图8
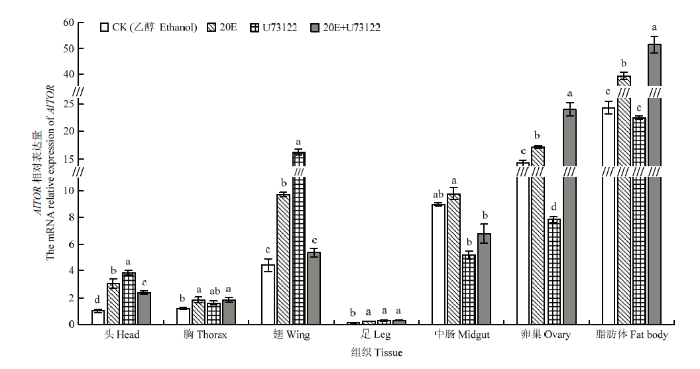 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同处理下绿盲蝽雌成虫各组织AlTOR的表达
Fig. 8Relative expression levels of AlTOR in different tissues of A. lucorum female adult under different treatments
2.5 重组蛋白AlTOR的表达、复性及纯化
重组质粒命名为pCzn1-AlTOR。随后将其转化至大肠杆菌Arctic Express(DE3)表达菌株中,经0.5 mmol·L-1 IPTG 20℃诱导12 h后,12% SDS-PAGE检测分析发现,pCzn1-AlTOR重组质粒表达一个约36 kD的蛋白(图9-A)。而不经IPTG诱导的重组质粒无此蛋白表达,说明AlTOR可在大肠杆菌细胞中诱导表达。但诱导表达的pCzn1-AlTOR表达蛋白均分布在沉淀体中,上清液中几乎没有,说明pCzn1- AlTOR表达蛋白主要以包涵体形式存在。包涵体经体外溶解、复性后,利用SDS-PAGE电泳分析pCzn1- AlTOR表达蛋白的纯化情况。用含20 mmol·L-1咪唑的缓冲液洗脱包涵体,进一步复性和纯化后在36 kD附近仅出现一条明显的特异性条带未见其他条带,说明已得到纯化的靶蛋白(图9-B)。采用蛋白定量试剂盒测定该蛋白的浓度为0.2 mg·mL-1,SDS-PAGE电泳检测该抗体的纯度达到85%以上,并最终获得了5.46 mg抗体,可以进行后续多克隆抗体制备工作。图9
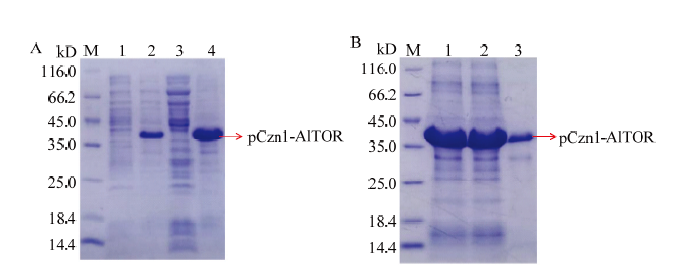 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9原核表达质粒pCzn1-AlTOR诱导表达蛋白的SDS-PAGE分析(A)和pCzn1-AlTOR表达蛋白纯化的SDS-PAGE分析(B)
M:蛋白分子量标准物Protein molecular weight marker。A:1:未经IPTG诱导的pCzn1-AlTOR表达蛋白pCzn1-AlTOR expressed protein without induced by IPTG;2:IPTG诱导的pCzn1-AlTOR表达蛋白pCzn1-AlTOR expressed protein induced by IPTG;3:重组质粒pCzn1-AlTOR表达蛋白上清Supernatant of pCzn1-AlTOR expressed protein induced by IPTG;4:重组质粒pCzn1-AlTOR表达蛋白沉淀Precipitate of pCzn1-AlTOR expressed protein induced by IPTG。B:1:未纯化的包涵体Unpurified inclusion body;2:未与Ni柱结合的杂蛋白Uncombined heterozygous protein with Ni NTA column;3:与Ni柱结合的目的蛋白Combined target protein with Ni NTA column
Fig. 9Expression SDS-PAGE analysis induced by IPTG (A) of pCzn1-AlTOR and SDS-PAGE of purified pCzn1-AlTOR fusion protein (B)
2.6 AlTOR蛋白多克隆抗体的特异性鉴定
新西兰白兔经3次免疫后,用间接ELISA法检测多克隆抗体的效价。AlTOR多克隆抗体和阴性血清(免疫前健康兔血清)按500×20—500×210连续倍比稀释。结果表明当AlTOR抗体稀释5 120倍时,OD阳性/ OD阴性>2.0,其终效价可达1:512 000。用制备的多克隆抗体分别检测原核表达的AlTOR重组蛋白及刚蜕皮的绿盲蝽3龄若虫,Western blot分析表明,所制备的多克隆抗体不仅能与绿盲蝽总蛋白结合,还可特异性与AlTOR重组蛋白反应(图10),且条带大小与预测的蛋白质大小一致,说明制备的AlTOR多克隆抗体准确、有效。图10
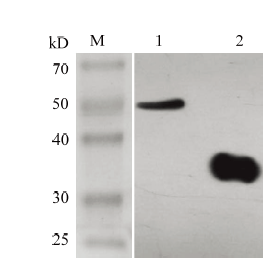 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10AlTOR多克隆抗体的Western blot 特异性分析
M:蛋白分子量标准物 Protein molecular weight marker;1:3龄若虫 3rd instar nymph;2:重组蛋白pCzn1-AlTOR Recombinant protein pCzn1- AlTOR
Fig. 10Western blot analysis of antibody against the expressed AlTOR protein
3 讨论
TOR是一类进化上保守的丝氨酸/苏氨酸(Ser/ Thr)蛋白激酶,是真核细胞响应环境信号调控生长和代谢的关键因子[6]。自TOR蛋白在酵母细胞发现后,在其他真菌、昆虫、植物以及哺乳动物中也相继被发现。TOR信号通路高度保守,对有机体自身生长和发展至关重要[24]。研究发现,植物、蠕虫、果蝇以及哺乳动物细胞中有一个TOR编码基因,而酿酒酵母中存在两个TOR编码基因[25]。TOR在细胞生长的营养调控中能通过激活下游信号通路,介导4E-BP1、S6K、eEF2等一系列翻译调节因子的磷酸化作用传递外界营养状况、生长因子等生长刺激信号,从而调节细胞内核糖体的发生及蛋白质的合成,进而综合调控细胞的生长与增殖[24]。另外,TOR还可参与调节真核细胞中mRNA和蛋白质的稳定性、细胞大小等与细胞生长密切相关的生理过程[26,27,28]。本研究通过筛选绿盲蝽转录组数据,RACE法获得了AlTOR的全长基因序列。序列分析结果表明,AlTOR的开放阅读框含有1 308 bp的核苷酸,编码435个氨基酸。多序列比对结果显示与其他半翅目昆虫TOR的氨基酸序列一致性在40%—60%,含有TOR作用靶标相关的SIN1结构域、CRIM结构域以及PH结构域。SIN1结构域是哺乳动物mSIN1的同源物,也是TOR作用的靶标之一,在信号转导过程中起着重要的作用。CRIM结构域位于TOR蛋白的中间位置,对TOR识别底物有着重要作用;另外CRIM结构域不论是在酵母细胞还是哺乳动物体内都高度保守。PH结构域同样也十分保守,其主要与脂质与膜结合相关[28]。进化树结果显示半翅目、膜翅目、鳞翅目等昆虫的TOR较为分化,位于不同的分支上。分化的原因可能这些目昆虫种类繁多、变态类型及由于不同的自然选择压造成的,如膜翅目、鳞翅目类昆虫是完全变态类昆虫,而半翅目昆虫属于不完全变态类昆虫。
TOR是昆虫体内营养信号的中心,通过氨基酸(AA)作为其调节因子,对外界摄入营养进行感知。AA/TOR途径广泛存在于真核细胞中,如酵母细胞。研究表明,除了调控葡萄糖的合成,TOR还可以通过氨基酸控制类胰岛素样肽的合成以及分泌。在果蝇成虫中,胰岛素样肽的分泌是由脂肪体产生的未配对细胞因子2(cytokine unpaired 2)远程调控从而响应营养信号[15,29]。由此可见,由TOR发出的氨基酸信号代表着营养信号的第一阶段,特别是对于需要蛋白质来启动产卵的昆虫而言[30]。此外,TOR信号还参与雌性昆虫生殖产卵,其作为营养信号的中心为雌虫生殖以及产卵提供充足的能量供给[12]。本研究发现,AlTOR在1—16日龄绿盲蝽中均有表达,表达量呈现波动式下降趋势,以初孵若虫(1日龄)中表达量最高;此外AlTOR在绿盲蝽雌成虫各组织中均有表达,其中卵巢、中肠及脂肪体中高表达,表明AlTOR的表达存在组织和时空特异性。研究表明,雌性昆虫的生殖需营养及能量资源的投入,在大多数昆虫中,营养的摄取是启动卵发育的关键触发因素。而AA/TOR途径和胰岛素途径作为营养传感器,在昆虫繁殖中起着至关重要的作用,影响生殖组织形成并调控JH和20E的生物合成[31,32,33]。TOR可刺激卵黄原蛋白在脂肪体的合成并激活卵发育;相反地,TOR途径的失活会导致繁殖受到抑制。在埃及伊蚊(Aedes aegypti)以及褐飞虱(Nilaparvata lugens)中,通过雷帕霉素抑制TOR蛋白质的合成以及RNAi介导的TOR基因沉默都会抑制卵黄原蛋白的表达,从而导致生殖力的降低[34,35]。在本研究中,AlTOR在若虫蜕皮期以及成虫羽化期表达水平显著提高,暗示TOR可能参与绿盲蝽若虫蜕皮以及羽化的变态行为;且AlTOR在卵巢、中肠及脂肪体中高表达暗示TOR可能参与绿盲蝽的生殖活动,这也是后续需要进行的研究工作。TOR可在蜕皮激素生成以及昆虫蜕皮过程中发挥重要作用。同时,蜕皮激素也会抑制TOR信号,在果蝇中,蜕皮激素会抑制幼虫进食时脂肪体内的TOR信号,进而产生未知信号,调节胰岛素生成细胞中胰岛素样肽的产生,从而控制果蝇全身性生长[32]。对家蚕的研究发现,蜕皮激素会在成虫的不同发育阶段在同一组织促进或抑制TOR信号从而调控成虫的生长发育[36]。在本研究中,绿盲蝽若虫随着龄期的增加,20E会显著上调AlTOR的表达,卵巢和脂肪体中AlTOR的表达也会由于20E的刺激而显著提高,表明AlTOR可能受20E调控,尤其是在“能量中心”脂肪体以及“生殖中心”卵巢中。细胞膜上的磷脂酶C是参与20E信号转导的关键酶,磷脂酶C可水解磷酯酰肌醇二磷酸成为第二信使IP3和DAG传递激素,神经递质和生长因子等信号行使其功能[37]。GU等研究表明,TOR信号参与家蚕前胸腺中促前胸腺激素诱导的20E合成,且磷脂酶C参与这个过程,当使用U73122抑制磷脂酶C时,会导致20E的合成受阻[38,39]。在本研究中,20E抑制剂U73211处理后,AlTOR在中肠和脂肪体中表达量显著下降,而20E处理后,AlTOR在中肠和脂肪体中表达量显著上调,暗示磷脂酶C可能参与20E对AlTOR的调控。
可溶性蛋白质的异源表达已经有较多的表达系统可用,目前很多可溶性蛋白以融合蛋白的形式得到成功表达[40,41]。本研究成功构建了pCzn1-AlTOR原核表达载体,并发现在20℃ 0.5 mmol·L-1 IPTG诱导12 h的条件下可大量表达AlTOR蛋白,之后通过变性、复性、蛋白纯化获得了纯化的靶蛋白。通过免疫新西兰白兔4次之后,采血检测,ELISA方法确定抗血清针对TOR蛋白的效价,通过ELISA检测效价得出,TOR抗体效价在512 000左右,纯度在85%以上,获得纯化的抗体为5.46 mg。Western blot分析结果也表明该多克隆抗体可同时与绿盲蝽总蛋白及AlTOR重组蛋白特异性结合,为进一步在蛋白层面研究TOR的功能打下了良好基础。
4 结论
AlTOR在绿盲蝽体内的表达谱显示出发育阶段特异性和组织特异性;20E及其抑制剂诱导后,AlTOR呈现出相反的应答反应;本研究获得的AlTOR重组蛋白的多克隆抗体具有高特异性。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1126/science.1187881URL [本文引用: 1]
DOI:10.1002/(ISSN)1526-4998URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nature11153URL [本文引用: 1]
DOI:10.1074/jbc.M209374200URL [本文引用: 1]
DOI:10.1016/j.biocel.2006.02.013URL [本文引用: 2]
DOI:10.1042/BST0370223URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1074/jbc.M110.141168URL [本文引用: 1]
DOI:10.1016/j.peptides.2018.02.001URL [本文引用: 1]
DOI:10.1016/j.jinsphys.2019.103902URL [本文引用: 1]
DOI:10.1016/j.devcel.2008.08.003URL [本文引用: 2]
DOI:10.1016/j.devcel.2009.10.008URL [本文引用: 1]
DOI:10.1016/j.devcel.2010.05.007URL [本文引用: 1]
DOI:10.1016/S0092-8674(03)00713-XURL [本文引用: 2]
DOI:10.1126/science.1119432URL [本文引用: 1]
[本文引用: 1]
DOI:10.1074/jbc.M501116200URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/ps.4203URL [本文引用: 1]
DOI:10.1074/jbc.M110.191783URL [本文引用: 1]
DOI:10.1253/circj.CJ-11-0870URL [本文引用: 1]
DOI:10.1006/meth.2001.1262URL [本文引用: 1]
DOI:10.1016/j.mad.2017.03.005URL [本文引用: 2]
DOI:10.1091/mbc.e15-08-0594URL [本文引用: 1]
[本文引用: 1]
DOI:10.1534/genetics.111.133363URL [本文引用: 1]
DOI:10.4161/cc.10.7.15063URL [本文引用: 2]
DOI:10.1016/j.cell.2012.08.019URL [本文引用: 1]
DOI:10.1016/j.bbrc.2017.01.031URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.cois.2015.08.003URL [本文引用: 2]
[本文引用: 1]
.
DOI:10.1016/j.jinsphys.2012.01.005URL [本文引用: 1]
DOI:10.3390/ijms17040438URL [本文引用: 1]
[本文引用: 1]
DOI:10.1146/annurev-physiol-030212-183750URL [本文引用: 1]
DOI:10.1016/j.ygcen.2019.01.009URL [本文引用: 1]
DOI:10.1016/j.jinsphys.2016.10.007URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}