 ,1, 郝婷丽2, 王海江1, 张泽1, 吕新,1
,1, 郝婷丽2, 王海江1, 张泽1, 吕新,1Advances in Cotton Growth and Development Modelling and Its Applications in China
HOU TongYu,1, HAO TingLi2, WANG HaiJiang1, ZHANG Ze1, Lü Xin,1通讯作者:
责任编辑: 杨鑫浩
收稿日期:2020-06-19接受日期:2020-11-25网络出版日期:2021-03-16
| 基金资助: |
Received:2020-06-19Accepted:2020-11-25Online:2021-03-16
作者简介 About authors
侯彤瑜,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (669KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
侯彤瑜, 郝婷丽, 王海江, 张泽, 吕新. 棉花生长发育模型及其在我国的研究和应用进展[J]. 中国农业科学, 2021, 54(6): 1112-1126 doi:10.3864/j.issn.0578-1752.2021.06.004
HOU TongYu, HAO TingLi, WANG HaiJiang, ZHANG Ze, Lü Xin.
开放科学(资源服务)标识码(OSID):

棉花(Gossypium hirsutum L.)是一种重要的经济作物和战略物资。棉花栽培种原属多年生植物,不仅具有无限生长的习性,而且其营养生长与生殖生长并进重叠的时间长,约占棉花生育期4/5。将棉花作为一年生作物进行栽培生产时,需要不间断采取多种促-控措施(如水肥、化调、脱叶催熟等技术)综合协调棉花营养生长和生殖生长间的矛盾,既要搭好高产的“营养架子”,又要确保优良的生殖生长,使棉花栽培管理的技术难度陡然增加。在棉田生态系统中,棉花的生长发育是中心环节,为了便于棉花的生产管理,农业研究人员开发了一系列棉花生长发育模拟模型[1,2]。这些模型大都从棉花的内部生理过程及其与生长环境的动态交互出发,解释作物生长发育和产量品质的形成,为实现棉田生产管理的优化决策提供了极大的帮助和便利。本文对国内外棉花生长发育模型研究进展进行文献综述,以期更好发挥棉花生长发育模型在我国棉花产业现代化中的作用。
1 棉花模拟模型简介
棉花生长发育模拟模型以计算机程序为主要媒介,以机理性或经验性数学公式为表达方式,定量描述棉花的光合、呼吸、蒸腾、生长、发育及蕾铃脱落等生理过程,系统阐述棉花基因、环境和栽培措施之间的交互关系,棉花形态结构、生理功能与生长环境之间的交互关系以及棉花地上-地下协同生长与生长环境间的交互关系,实现棉花生长发育进程的动态模拟和产量品质的提前预测,为棉花育种和栽培决策提供技术支持[3,4,5,6,7]。从模型功能特点上来划分,棉花生长发育模拟模型可以分为生理生态过程模型、形态结构模型和功能-结构模型3类。棉花生长模拟模型一般包括参数输入、生育模拟、结果输出3部分。输入参数主要包括品种遗传属性、土壤理化特征、温光热气象资源以及栽培管理措施[8,9]。生育模拟是棉花生理生态过程模型的核心,一般包括生育期、干物质积累、干物质分配、器官形态结构建成、土壤-植物动力学交互、土壤-植物-大气连续系统等方面的机理过程[3,5-6,10],部分模型还包含病虫害-植物交互子模块,为优化棉花病虫害管理提供决策支持[11]。棉花生长发育模拟模型一般以小时或日为时间步长进行模拟,在每个时间节点都会有一系列结果输出,主要包括植物器官生长发育、土壤理化参数的变化、病虫害发展规律、产量和品质等,为棉花生产的优化决策提供支持。通过将棉花生长发育模拟模型与其他模型相结合,还可以进一步丰富输出结果,如与棉花经济金融模型相耦合实现棉花生产效益和期货投资的评估[12],与气候变化或气候灾害模型相耦合实现棉花生产灾害和保险评估[13,14],与三维动画模型相结合实现棉花生长发育的可视化,从而为棉花栽培管理教学提供支持[2]。
2 棉花模拟模型国内外研究进展
2.1 棉花生理生态过程模型
棉花生理生态过程模型的研究起始于20世纪60—70年代,重点关注棉花基础生理代谢过程中能量流动和物质平衡的定量模拟,依据定量关系构建方法的不同可分为统计模型和机理模型2类。统计模型基于统计学原理而构建,构建过程和参数体系相对简单,但模型的运行结果对时间和空间条件的变化比较敏感,实际使用中需要提前对模型参数进行本地化调校,以符合当地的棉花生产实际。而机理模型则大多基于植物学原理构建,机理性较强,可以在一定程度上超越时空限制而在多个区域取得相对较好的模拟效果,但是机理模型的构建过程需付出很大的努力和代价,使用起来也复杂,往往需要输入较为详细的参数,亦在一定程度上限制了此类模型的大面积推广和运用(表1)。Table 1
表1
表1国外经典棉花生理生态过程模型要点比较
Table 1
| 模型要素 Model components | GOSSYM[5] | Cotton2K[10] | OZCOT[6] | CSM-CROPGRO-Cotton[8] | |
|---|---|---|---|---|---|
| 模型输入 Input | 环境 Environment | 日尺度气象数据;土壤初始水、氮含量 Daily weather data, initial soil water and nitrogen content | 小时尺度气象数据;土壤初始水、氮含量 Hourly weather data, initial soil water and nitrogen content | 日尺度气象数据;土壤初始水、氮含量 Daily weather data, initial soil water and nitrogen content | 日尺度气象数据;土壤初始水、氮含量 Daily weather data, initial soil water and nitrogen content |
| 品种 Cultivar | 品种遗传参数 Genetic coefficients | 品种遗传参数 Genetic coefficients | 品种遗传参数 Genetic coefficients | 品种遗传参数 Genetic coefficients | |
| 栽培 Management operations | 播期、密度、灌溉、施肥、化调、脱叶 Planting date, plant density, irrigation, fertilizer, growth regulators and defoliation | 播期、密度、滴灌、施肥、耕作、化调、脱叶 Planting date, plant density, drip irrigation, fertilizer, tillage, growth regulators and defoliation | 播期、密度、灌溉、施肥、脱叶 Planting date, plant density, irrigation, fertilizer and defoliation | 播期、密度、灌溉、施肥、留茬、耕作、脱叶 Planting date, plant density, irrigation, fertilizer, residue, tillage and defoliation | |
| 生育进程 Phenology | 基于日平均温度和碳氮供需平衡调节生育进程 Develop based on daily thermal time and C:N ratio | 基于小时尺度平均温度和碳氮供需平衡调节生育进程 Develop based on hourly thermal time and C:N ratio | 基于出苗到现蕾所需积温的经验值计算现蕾时间 Based on the empirical accumulating day degrees between sowing and the appearance of the first square | 基于生理日数模拟生育进程 Develop based on physiological degree days | |
| 物质积累和分配 Dry matter accumulation and allocation | 光合 Photosynthesis | 基于群体冠层对太阳辐射的截获效率计算潜在光合 Canopy-level radiation interception | 基于群体冠层对太阳辐射的截获效率计算潜在光合 Canopy-level radiation interception | 基于群体冠层对太阳辐射的截获效率计算潜在光合 Canopy-level radiation interception | 将单叶小时尺度的潜在光合整合为日尺度的冠层光合 Leaf-level biochemistry |
| 呼吸 Respiration | 基于光强、温度及生物量的经验公式 Uses an empirical function of respiration based on light, air temperature and biomass | 由器官的物质构成确定生长呼吸;由光合生产量确定维持呼吸 Calculates growth and maintenance respiration and photorespiration | 基于果节数量和温度调节因子的经验公式 Uses empirical functions of respiration based on fruiting site count and air temperature | 由器官的物质构成确定生长呼吸;由光合生产量确定维持呼吸 Calculates growth and maintenance respiration | |
| 分配 Partitioning | 根据各类器官的需求将积累的干物质按比例分配 Allocates carbon to individual growing organs based on the organ's contribution to the total demand | 根据各类器官的需求将积累的干物质按比例分配 Allocates carbon to individual growing organs based on the organ's contribution to the total demand | 将积累的干物质分配给每个棉铃,以估计棉铃的生长 Allocates carbon to cohort pools for developing bolls | 按照生殖器官优先原则和各器官生长需求实现干物质动态分配 Reproductive tissues have first priority, then allocates carbon to single pools for leaves, stems and roots | |
| 器官生长 Organ growth | 受温度、水分、氮素及碳水化合物供应状态调控的潜在生长 Potential growth with the stresses related to air temperature, water, C, and N | 受温度、水分、氮素及碳水化合物供应状态调控的潜在生长 Potential growth with the stresses related to air temperature, water, C, and N | 受温度、水分、氮素及碳水化合物供应状态调控的潜在生长 Potential growth with the stresses related to air temperature, water, C, and N | 受温度、水分、氮素及碳水化合物供应状态调控的潜在生长 Potential growth with the stresses related to air temperature, water, C, and N | |
| 蕾铃脱落 Shedding of buds and bolls | 根据蕾铃的碳氮供应模拟生理性脱落;兼顾虫害、阴雨对蕾铃脱落的影响 Physiological shedding based on the carbon and nitrogen stress, and the other shedding based on the insects or weather stresses | 根据蕾铃的碳氮供应模拟生理性脱落;兼顾虫害、阴雨对蕾铃脱落的影响 Physiological shedding based on the carbon and nitrogen stress, and the other shedding based on the insects or weather stress | 根据棉铃承载力与实际载铃量的比值确定棉铃脱落状态 Shedding based on the ratio of load to carrying capacity | 当干物质分配无法满足生殖器官生长需求时即发生脱落 Shedding occurs when dry matter allocation cannot meet the growth needs of reproductive organs | |
| 模型要素 Model components | GOSSYM[5] | Cotton2K[10] | OZCOT[6] | CSM-CROPGRO-Cotton[8] | |
| 水分平衡 Water Balance | 土壤 Soil | 2D RHIZOS模型 2D RHIZOS model | 2D RHIZOS 模型 2D RHIZOS model | Ritchie水分平衡 Ritchie model | Ritchie水分平衡 Ritchie model |
| 蒸散 ET | Ritchie水分平衡 Ritchie model | CIMIS彭曼公式 CIMIS Penman model | Ritchie水分平衡 Ritchie model | FAO-56彭曼公式 FAO-56 | |
| 氮素平衡 Nitrogen Balance | 2D RHIZOS模型 2D RHIZOS model | 对2D RHIZOS模型土壤氮素动态平衡模块进行了优化 Optimized 2D RHIZOS model | 基于氮素池实现土壤-棉花-器官间氮素动态平衡 Dynamic nitrogen pools | 由土壤-棉花氮平衡模块实现土壤-棉花-器官间氮素动态平衡 Based on the soil carbon and nitrogen balance sub module | |
| 模型输出 Output | 棉花产量、株式图、水分利用效率、氮肥利用效率等 Yield, plant maps, WUE, NUE | 棉花产量、株式图、水分利用效率、氮肥利用效率等 Yield, plant maps, WUE, NUE | 棉花产量、载铃量、水分利用效率、氮肥利用效率等 Yield, boll load, plant maps, WUE, NUE | 棉花产量、生物量、载铃量、水分利用效率、氮肥利用效率等 Yield, biomass, boll load, plant maps, WUE, NUE | |
| 应用情况 Application | 机理性强,输入参数多,使用前需进行参数校验,在美国植棉区已经广泛应用[1] Mainly applied in the cotton belt of U.S.[1] | 针对干旱半干旱环境和栽培措施的优化使其在以色列[10]、新疆[28]等植棉区应用较多 Mainly applied in arid and semi-arid environments such as Israel[10] and Xinjiang China[28] | 重点关注棉铃生长和脱落,对底层生理考虑较少,受澳大利亚官方支持应用广泛[2] Mainly applied in Australia [2] | 模块化组织提升了开发应用便捷性,目前在美国东南植棉区应用较多[2,32] Mainly appliied in in the southeastern U.S. [2,32] | |
新窗口打开|下载CSV
棉花生理生态过程模型一般用于农田尺度生产管理优化、产量品质预测、土地生产力评价等方面,在部分植棉区域的农业生产中得到广泛应用[2]。但是,由于模型构建过程往往针对特定的时空区域,而模型本身又需要较多的输入参数,因此,如何在区域尺度进一步提升模型参数的获取和模型结果的精度是此类模型未来发展的重要方向之一[15,16]。
2.1.1 国外棉花生理生态过程模型发展及其特点
(1)GOSSYM模型 在1970年,美国农业工程师STAPLETON[17]开发了一套以辐射和温度为驱动因子,模拟养分亏缺和虫害发生对棉花生长发育影响的棉花生长发育模拟系统。通过逐步引入水、氮平衡[18,19]、光合生产[20,21]、土壤物理和根系生长模拟[5,22-23]、数据输入、管理和模拟结果的解析[24]等模块而构建了更加完善的棉花模拟模型和决策支持系统GOSSYM/ COMAX(图1)。GOSSYM/COMAX模型以物质平衡原理为基础,可以实现水、碳和氮在土壤和植株中运移转化过程的模拟。目前该模型成功在全美棉花种植区域得到推广应用[24,25]。
图1
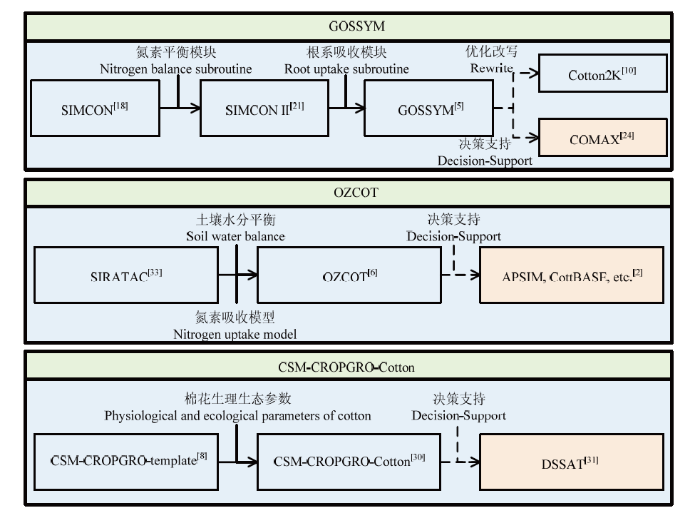 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1国外经典棉花生理生态过程模型发展示意图
Fig. 1A brief history of cotton growth and development modelling
(2)Cotton2K模型 为了提高GOSSYM模型对干旱地区灌溉棉花生长发育的模拟精度,MARANI对GOSSYM模型进行了修改和重写,即Cotton2K模型[10](图1)。该模型要求输入小时尺度的气象数据(表1),将干旱地区对棉花生长发育最敏感的蒸腾和光合等生理模块和土壤水分供应模块精细化到小时尺度,并在此基础上对植物生长、物候变化、滴灌过程、土壤氮的矿化和硝化等过程进行更新和细化[26],从而实现了干旱地区棉花植株-土壤-田间管理间交互作用的优化模拟。目前该模型已成功用于国内外干旱地区棉花生长模拟及管理决策[27,28,29]。
(3)CSM-CROPGRO-Cotton模型 DSSAT(Decision Support System for Agrotechnology Transfer,www. DSSAT.net)是由作物模拟模型支持的决策支持系统,是目前使用最广泛的农业生产模型系统之一,其用于模拟棉花生长发育的模块为CSM-CROPGRO-Cotton模型。该模型依托CSM(Cropping System Model)通用模块对自然环境要素和人为管理决策的影响进行模拟[8],基于棉花基因型数据、生理生态参数及田间试验结果,对系统中主要用于双子叶植物生长发育过程进行模拟的样板模型CORPGRO进行参数化,用得到的CROPGRO-Cotton模块进行棉花生长发育及产量的模拟[30](图1)。目前,CSM-CROPGRO-Cotton模型包含在最新版DSSAT中,可以直接下载使用[31]。目前该模型在美国东南植棉区应用较多[2,32]。
(4)OZCOT模型 为了对棉田植保方案提供决策支持,澳大利亚联邦科学与工业研究组织和农业决策部门共同研制了棉花害虫计算机管理系统SIRATAC模型[33]。该模型包含了一个以棉铃生长和存活为核心的模块[34],可以对棉铃从生长到成熟(或脱落)进行全程监控。通过对SIRATAC模型的光合和呼吸过程进行了修改和优化,并引入作物-土壤水分平衡模型和氮素吸收模型[35],HEARN提出了OZCOT模型[6](图1)。在澳大利亚,OZCOT是目前唯一受官方支持的棉花生长发育模拟模型[2],是HydroLOGIC[36]、Whopper Croper[37]等棉花灌溉决策支持模型的关键内核,同时也是APSIM中进行棉花生长发育模拟的核心。
此外,还有些为特殊目的而开发的棉花生理生态过程模型,如WALLET等[38]为分析全球气候变化背景下CO2浓度和气温的升高对棉花生长发育的影响而开发了COTCO2模型,LIANG等[39]将改进后的GOSSYM模型与区域气候-天气预测模型(CWRF)相耦合,并基于该模型研究了区域气候变化对棉花产量的影响[14]。还有些则通过对通用作物模型添加或修改相关模块后使其适用于棉花生长发育的模拟,如荷兰Wageningen大学开发WOrldFOodStudies模型[40],由联合国FAO(Food and Agriculture Organization)开发的AuqaCrop通用植物生长发育模型[41],禾本科通用作物模型GRAMI[42,43,44]和CropSyst[45]等都可以用于棉花生长发育的模拟。
2.1.2 我国棉花生理生态过程模型发展及其特点 20世纪80—90年代我国****在借鉴国内外棉花模拟模型开发经验的基础上,对棉花生长发育模拟进行了一系列探索。吴国伟等[46]建立了用于与棉虫动态模型偶联的棉花生长发育模型,以明确最佳栽培措施,优化棉虫动态经济阈值。翟连荣等[47]针对国外作物-害虫管理模型的缺陷,通过耦合棉铃虫取食模型、种群动态模型及棉花生长发育模型,增加了部分蕾铃去除及棉花遭受虫害后自身的补偿作用,构建了田间棉铃虫的管理系统。刘文等[9]以气象要素和植物生育状态变量驱动棉花生育期、生长、呼吸、干物质分配、蕾铃消长、水分胁迫等方面的生理过程,从潜在生产和水分胁迫两个层次上建成棉花生长发育与产量形成逐日动态模拟模型。冯利平等[48]结合田间试验和专家经验,研制了可用于棉花生产优化管理、生长发育和产量形成预测的决策系统。
潘学标等[3]在参考国外多个成熟棉花生长发育模型经验(美国的GOSSYM模型、荷兰的MACROS模型以及澳大利亚的OZCOT模型)的基础上,根据我国黄淮海地区棉花生产实际情况,构建了以日为时间步长,可用于定量模拟气象、土壤及栽培管理措施等农业生产要素对棉花生长发育、形态发生及产量品质影响的棉花生长发育模型COTGROW。该模型包含棉花碳素代谢、水分平衡、氮素平衡、干物质生产与分配等模块,结合我国棉花的生产实际,对棉花生长发育与环境条件及栽培措施的交互关系进行了分析和模拟,可以进行播期、密度、施肥、灌溉、生长调节剂等生产管理决策的推荐,能够实现棉花生长发育和产量形成的动态模拟。
虽然COTGROW模型针对中国棉花品种和生产管理实际进行了优化,但是该模型无法实现覆膜滴灌、整枝打顶、棉麦间套作等栽培技术措施的模拟。针对Cotton2K模型对干旱地区滴灌棉花进行优化的特点,杨艳敏等[49]利用新疆棉花试验数据对该模型进行参数校验,并增加了整枝和打顶模块,使该模型的模拟结果更加符合新疆棉区棉花生长管理的实际[28]。ZHANG等[50]在荷兰SUCROS作物生长发育基础模式的基础上提出了SUCROS-cotton模型,通过引入我国特有的棉麦间套作种植体系以及地膜覆盖等栽培管理措施,使其在生态环境、种植密度、棉花品种等具有较大差异的黄河流域棉区和西北内陆棉区均取得较好模拟效果,为中国不同棉区的棉花种植区划、理想株型塑造和管理措施优化提供理论和技术支持。
棉铃、棉籽和棉纤维的生长发育过程与棉花产量品质的形成密切相关,定量研究棉铃发育及纤维品质形成是棉花生长模拟研究的重要内容。马富裕[51]通过在我国三大棉区同步开展棉花“异地分期种植”试验,定量分析了温度、光照和水分等生态因子对不同品种棉铃干物质增长及品质形成的影响,并建立了棉铃干物质积累及纤维品质形成模拟模型。LI等[52]研究了棉铃发育进程与温度、光照、氮素供应及品种要素的定量关系,提出了棉铃生理发育时间的概念,建立了棉铃成熟进程模拟模型。根据棉铃、棉籽和棉纤维的同步发育特性,以棉籽作为“库端”驱动生物量、油脂和蛋白质在棉籽中的积累过程,以日为时间步长定量模拟了棉籽干物质质量、物质组分含量及其品质的形成过程[53]。在此基础上,CHEN等[54]进一步考虑了水分胁迫对棉铃发育成熟进程的影响,并构建了棉籽中胚的干物质积累及其物质组分动态变化模拟模型。
相对于棉籽而言,棉纤维生长发育及其品质参数的定量模拟对于准确了解棉花产量、品质和经济效益而言更具意义。在准确模拟棉铃生理发育时间的基础上,ZHAO等[55,56]基于棉铃与棉纤维的同步发育特性确定棉纤维的伸长期、加厚期和成熟期,通过棉铃不同生长阶段的辐热积与棉纤维长度和强度的关系以及不同着生部位棉铃的氮素供应状况实现了棉纤维长度、强度及马克隆值的模拟。这些模型的开发和应用不仅能够提升棉花产量、品质的模拟精度,对于系统研究外界环境对棉花生殖器官生长发育的影响,分析和预测棉花生产价格和效益具有重要意义。
2.2 棉花的形态结构模型
棉花形态结构指棉花各个器官(节间、叶片、叶柄、花铃、根系)之间的拓扑连接关系,以及在此基础之上定义的各个器官大小、形态及其在三维空间位置和方向[57,58]。与棉花生理生态模型对作物群体尺度的关注不同,形态结构模型以棉株个体为研究中心,更加重视对植株个体及其器官真实形态结构的精确描述,并以三维可视化的方式反映棉花形态结构的生长发育规律[59]。植物形态结构模型大致可以分为静态模型和动态模型两大类[57,59]。其中静态模型是指基于接触式或非接触式数据采集仪,获取特定时刻植物不同器官的空间坐标信息,采用计算机图形学方法获得视觉上具有真实感的三维植物形态[57]。静态模型主要用于研究与植物空间结构相关的性质,对植物冠层结构的动态演变规律缺乏关注。动态模型是指基于植物生长过程中拓扑结构和几何结构的演变原理和规律,依赖于特定的计算机图形学算法(如字符串替换系统[60]、参考轴技术[61,62]、多尺度图树法[58]等)而建立的能够反映植物生长过程中形态结构动态变化的模型,为模拟环境要素对植物冠层结构生长动态的影响提供了便利。
目前,棉花形态结构模型在冠层光分布的精确模拟研究等方面显示出了很大的优势[63],但是该类模型在构建过程中基本不考虑其底层生理功能,难以定量模拟棉花植物生长发育随环境而改变的状况,也没有器官生物量等信息,因而这类模型在棉花实际生产领域的应用存在着很大的局限性[57,59]。
2.2.1 国外棉花形态结构模型及其特点 MUTSAERS[64]通过对棉花植株和冠层的精细分层,开发了能够考虑棉花形态发生过程的生长发育模拟模型KUTUN。该模型对棉花冠层进行分层模拟时,每层仅包含一段主茎节间、一个主茎叶以及在该主茎叶的叶腋着生的果枝,模型假设每一层的光合产物会优先满足相应层生殖器官和营养器官生长发育的需求,模拟了每一层器官的发生生长、光合同化及干物质分配的循环过程。该模型虽然在一定程度上考虑了棉花植株器官的空间分布特性对棉花生长发育的影响,然而从模型架构和假设来看,该模型对棉花各器官的形态结构做了大量简化,属于棉花生理生态过程模型到棉花形态结构模型的过渡。
在棉花静态形态结构模型构建方面,LANG[65]在1973年即采用自制的三维数字化仪测定了棉花叶片的三维空间坐标,分析了棉花冠层中叶面积及叶倾角的分布规律,研究了棉花叶片的向光性运动。SINOQUET等[66]采用三维数字化仪获取表征棉花叶片位置和朝向的空间三维坐标,之后采用图像合成软件构建了棉花静态三维结构模型,基于该模型进一步模拟了棉花叶片和冠层对太阳直射辐射和天空光散射辐射的截获效率,结合叶片的光响应特性计算了冠层的光合效率。与棉花生理生态过程模型相比,基于棉花形态结构模型能够更加精准地描述不同棉花叶片的辐射截获特征[66]。
在棉花动态形态结构模型构建方面,DAUZAT[63]采用三维数字化仪在现蕾期、开花期及脱叶前,分别测定了棉花主茎节、果枝节、叶枝节及对应节位上叶柄和叶片的三维坐标,并采用多尺度图树算法构建了棉花植株的拓扑结构。依据每周测定的棉株节数及叶面积的动态变化规律,结合棉花器官发生、叶面积扩展、叶柄伸长、果枝等几何结构参数随生育期和外界环境的变化特征,采用插值法构建了棉花冠层结构动态模型。基于该模型进一步模拟了冠层光截获随冠层叶面积指数的定量变化,提出了冠层光质可能是造成不同种植密度棉花主茎节间及果枝朝向可塑性变化的主要原因。
2.2.2 我国棉花形态结构模型及其特点 在棉花静态形态结构模型构建方面,杨娟等[67]将棉花植株的拓扑连接划分为主茎和果枝两大类,基于棉花各器官的数字图像获取造型控制点二维坐标,基于控制点坐标采用NURBS曲面方法建立了棉花的各种器官的三维结构模型,结合棉株拓扑结构和器官三维形态,形象直观地再现了棉花植株的三维结构。该研究重点从计算机图形学角度提出了构建视觉逼真棉花器官的技术,对其生物学意义关注较少。
在棉花动态形态结构模型构建方面,周娟等[68]依据对棉花盆栽试验数据的系统分析,定量描述了棉花形态建成与品种、环境和技术措施间的动态关系,基于数学建模技术实现棉花形态器官三维生长形成过程的动态模拟;依据前人研究和数字图像提取了不同生育期棉株拓扑连接信息,结合计算机图形学方法构建了棉花虚拟生长结构模型VGSC。该模型可以动态模拟棉花植株各器官随生长度日、氮素、水分及DPC化控等要素的变化过程。
陈超等[69]应用COTGROW棉花生理生态过程模型进行棉花生长发育、干物质积累、产量形成等过程的定量模拟,基于田间试验数据系统分析了棉花各器官的生物量-形态间的定量关系,据此构建了基于棉花生理生态过程的形态结构模拟模型,为进一步实现棉花器官的三维可视化模拟提供了基础。与VGSC模型相比,该模型没有完成棉花植株生长动态的三维可视化,但是其对棉花生长发育底层过程描述的机理性更强。
2.3 棉花的功能-结构模型
随着研究的深入,棉花生长发育模拟模型趋向于依据一定的规则将棉花的底层生理代谢功能模型与棉花形态结构描述相耦合,实现棉花生理功能-形态结构的动态交互,在更精准的时间尺度和空间维度上进行棉花生长发育状况的动态模拟[70,71,72]。为此,研究人员提出了2种建模思路[73]:一类是以某些生育变量为依托,直接将成熟的植物生理代谢模型与形态结构模型相结合,实现植物生长过程中结构功能的互反馈,如L-Cotton模型[71]、COTONS模型[72]等;另一类是针对植物结构和功能的特点,在同一模型中对二者的交互进行统一设计和模拟,以并行方式模拟二者的反馈机制,如GreenLab模型[74,75]等。目前,此类模型的研究仍处于初级阶段,各种功能模型和结构模型的耦合方式仍存在很多需要研究的问题,但是该模型代表了棉花生长发育模型的最终方向,对棉花生产管理和高效生产展现了极大的发展潜力[76]。2.3.1 国外棉花功能-结构模型发展及其特点 DE REFFYE等[77]将植物生长发育与蒸腾效率间的生理代谢关系加入到植物形态结构模拟系统AMAPpara中,利用植物的蒸腾效率计算干物质生产,基于源-库分配理论将干物质分配给各器官,基于异速生长率计算各器官的几何结构参数。之后,根据各器官几何结构参数与水力学参数的关系进一步模拟冠层的蒸腾效率,从而实现植物功能-结构交互作用的模拟。通过田间试验对模型参数进行校验,模拟结果能够很好地反映棉花生长发育和形态结构的动态演变,但是该模型对生殖器官生长发育过程的描述比较简单。
HANAN[71]将基于计算机图形学原理的植物结构模拟模型L-System[60]与传统棉花生长发育模型OZCOT[6]结合起来,构建了植株尺度的棉花功能-结构模型L-Cotton。与AMAPpara相比,该模型更加关注棉花冠层内每个蕾铃的发生、生长、发育及其脱落等生殖生长和发育过程,基于碳平衡原理实现了水、氮等环境胁迫对棉花冠层内蕾铃生长数量、脱落数量及未脱落蕾铃大小的动态调节和对棉花最终产量影响的模拟,并对模拟结果进行三维可视化表达。RENTON等[78]以叶片生物量生产为媒介,将基于L-System模拟的植物结构与基于典范模型(Canonical Model)方法构建的植物功能模型相耦合,模拟了叶片脱落后棉花植株的补偿生长效应。
JALLAS等[72]将棉花生长模型GOSSYM的模拟结果与棉花冠层三维结构模型相耦合进行棉花生长发育的模拟。模拟过程首先由GOSSYM模型依据气象、土壤、品种及田间管理措施等输入要素,实现棉花器官的发生、脱落、连接等拓扑结构参数和器官干物质、面积、长度、直径等几何结构参数的动态模拟,之后根据田间试验调查统计规律,将GOSSYM模拟的各器官形态结构参数动态转换为三维空间中由简化多边形及其组合构建的具有视觉真实感的棉花器官,并通过计算机图形学算法对模拟结果进行可视化表达。他们认为采用三维虚拟模型对棉花生长发育结果进行表达能够提高农民对模型的认识,使模型的使用更加高效,有助于通过植物模型提高农业新技术的转化,降低转化成本。
2.3.2 我国棉花功能-结构模型发展及其特点 通用植物功能-结构模型GreenLab起源于AMAP模型,其结构模型和功能模型均基于一般性的假设,通过一系列参数优化过程即可实现各种特定植物功能-结构动态交互的模拟[57,79]。ZHAN等[74]采用田间试验数据对GreenLab模型及逆行参数优化,模拟了棉花植株的干物质生产、分配及形态结构对冠层下部叶片脱落的响应过程。张吴平等[75]采用GreenLab模型的原理,基于不同类型根个体获取生物量的能力和异速生长规则,实现了根个体生物量分配及几何结构生长的模拟,根据根个体空间伸展规则与空间体元技术,构建了棉花根系生长发育模拟模型。
开源植物功能-结构建模平台GroIMP是基于L-system的重写规则所建立,能够进行植物功能-结构动态交互模拟[80]。陈超等[81]对COTGROW模型的发育和形态发生模块做了改进,并将其与GroIMP相耦合[80],实现了棉花地上部器官形态建成和生长过程模拟的可视化,应用GroIMP中的光分布模块模拟了不同密度棉花冠层的光截获量。GU等[70]则基于SUCROS- cotton模型和GroIMP平台开发了棉花功能-结构模型CottonXL,模拟了植物生长调节剂缩节胺对不同植株密度棉花生长发育和植株结构的影响。WANG等[82]基于CottonXL模型,在单个棉铃尺度上模拟了棉花播种时间、打顶时间以及是否进行薄膜覆盖等管理措施对棉纤维长度、纤维强度和麦克隆值的影响。
3 棉花模拟模型在我国应用进展
美国是世界上开发和应用棉花生长发育模型最早,也是最成功的国家。GOSSYM/COMAX模型在20世纪80年代已经在全美所有植棉区域得到广泛应用[25],对于优化美国棉花生产管理、推动棉花产业节本增效具有发挥了巨大的作用[2]。目前,我国是世界第二大植棉国,产量约占全球产量的1/4[83],但是棉花生长发育模型在我国棉花生产实际中的应用数量、应用水平和应用场景仍处于初级阶段。由于我国棉花种植过程中人工参与的技术环节偏多,水肥管理效率偏低,导致植棉成本显著增加,单位面积总成本约为美国的3.4倍[84]。因此,在我国棉花生产实际中加大棉花生长发育模拟模型应用水平对于推动我国棉花生产发展,提升我国植棉效益方面潜力巨大。在作物生长发育和产量模拟方面,董占山等[85]运用黄河流域棉区田间试验数据对GOSSYM模型进行了品种参数校验,通过适当调整模型品种参数,GOSSYM模型可以实现中熟品种中棉所12、中早熟品种中棉所17、早熟品种中棉所16的生长发育和产量形成过程的模拟。赵黎等[86]基于新疆石河子148团田间试验数据对GOSSYM模型进行了参数校验,应用校验后的模型模拟了北疆主栽品种新陆早6号和新陆早7号的生长发育和产量形成过程。结果表明部分修改和校正后的GOSSYM模型可以运用于模拟棉花的生育期、产量以及棉花主要性状、干物质重量。
在灌溉管理方面,杨艳敏等[49]利用新疆棉花试验数据对Cotton2K模型进行校验的基础上,增加了整枝和打顶相关模块,使该模型更加适合新疆棉区棉花生长发育的模拟[28],并成功运用该模型对华北平原和新疆的棉花生产和耗水进行了模拟,分析2个地区棉花的耗水特征[87]。吴立峰等[88]针对新疆特色膜下滴灌栽培管理模式,通过新疆石河子的棉花田间试验对DSSAT-CROPGRO-Cotton模型进行了本地化模型参数率定,并对该模型进行了敏感性分析和不确定性分析,实现了灌溉制度的优化。LI等[89]则通过田间试验和STICS算法优化了CSM-CROPGRO-cotton模型中潜在蒸散和土面蒸发的计算方法,显著提升了该模型对新疆膜下滴灌棉花的生长发育和土壤水分变化的模拟精度。TAN等[90]则应用AquaCrop模型模拟了不同灌溉方式对膜下滴灌棉花产量、土壤水分及土壤盐分含量变化的影响。
在施肥管理决策方面,孙莉等[91]依据新疆北部地区不同施肥量田间试验,对Cotton2K模型进行了参数校验,采用该模型对棉花植株中氮素浓度进行了模拟。针对模拟结果与实测值差异偏大的问题,作者认为应该在控制田间试验土壤初始含氮量的基础上,进一步考虑不同生育时期棉花氮肥吸收的差异性[91]。由于棉花生长发育模型所需的各种环境参数难以及时大面积获取,目前我国棉花实际生产中大多采用3S技术实时获取棉花营养状况和生长状态,并依据棉花需肥规律进行施肥决策,如吕新等[92]应用地理信息系统、数学模型等方法建立了土壤肥力评价模型和评价体系,结合肥料试验建立了棉花施肥决策支持系统,具有生成土壤肥力分布图、分析土壤肥力资源、提供棉花施肥咨询与配方推荐等功能。
在病虫害管理方面,虽然国内外均建立了与棉花生长发育状况耦合性较强的病虫害管理模型[32,47],但是这些模型在我国棉花实际生产中应用较少。目前,我国棉花病虫害管理主要通过高清图像[93]、高光谱遥感[92,93]等技术手段采集棉花叶片的图像或光谱信息,采用深度学习等图像和光谱特征分析方法进行棉花病害[94,95,96]及虫害[97,98]的类型诊断和程度评估。
在区域尺度棉花生产系统评估方面,潘学标等[15]采用作物模拟模型和相关分析相结合的方法开发了以光能利用为核心,同时考虑气候、土壤、作物品种、管理措施影响的综合评估模型,应用该模型模拟了新疆莎车田间棉花的干物质生长动态、叶面积动态,并进一步分析了该模型在评估气候变异对新疆棉花生产影响中的应用前景。陈超等[99]和王雪娇等[13,100]则进一步应用该模型成功预测了新疆棉花冷害和产量动态。
4 我国棉花生长发育模型研究应用的建议
从20世纪60年代以来,棉花生长发育模拟模型经历了60年的发展,从棉花生理生态过程模型到棉花形态结构模型,再到棉花功能-结构模型,在世界范围内得到了广泛的研究和应用。目前,我国的棉花生长发育模拟模型基本还是以引进消化国外的模型为主,在模型的本地化研究、尺度提升及其推广示范方面仍存在一些问题,需要在以后的研究和应用中加以改进和完善。4.1 棉花生长发育模型的本地化研究和应用
我国棉花生产主要集中在黄河流域棉区、长江流域棉区和西北内陆棉区,其中新疆棉区以占全国植棉面积76.1%,占全国植棉总产量84.9%的体量,成为我国最大的棉区(国家统计局,2019)。我国不同植棉区间地理区位和气候环境差异较大,在长期植棉历史中逐渐形成各具特色的棉花生产栽培管理技术规范,如新疆棉区经过多年探索逐步形成的以“矮、密、早、膜、滴”为主要内容的综合植棉技术(图2)。因此,在借鉴国内外棉花生长发育模型优势的基础上,针对不同植棉区域特色播种方式(如地膜覆盖[89])、灌溉方式(如滴灌[101])和栽培方式(密植打顶[28])等生长管理措施,进一步开展棉花地上和地下部生长发育、营养生长和生殖生长及其与环境交互、棉花功能和结构交互作用的机理和模型的开发研究[102],加强多环境要素胁迫(高温、干旱、病虫害等)对棉花生长发育综合影响的机理和模型研究,加强地膜残留、脱叶剂、缩节胺等对土壤和生物环境影响的评估模型研究,加强棉花生长发育模型与棉花经济效益模型的耦合研究[103],是我国棉花生长发育模型未来研究的重点方向。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2新疆棉区典型膜下滴灌生产场景示意图
Fig. 2Schematic diagram of typical cotton production scenarios of drip irrigation under mulch in Xinjiang
4.2 棉花生长发育模型由单点到区域的尺度提升
目前的棉花生长发育模型多基于温室控制试验或田间小区试验样本构建,偏重棉花生长发育动态机理,需要输入大量难以获取的参数才能获得较为准确的模拟结果,因此主要用于单点或者田间尺度棉花生长发育过程的模拟和预测,大面积推广受到限制[2]。随着搭载各种传感器的农业物联网技术和星-空-地一体化多层次农业遥感技术的发展,能够在大时空范围内快速、准确、定量地获取棉花生长发育模型所需要的土壤、植被、气象等输入参数,同时也可以定量反演如土壤水分含量、叶面积指数、叶绿素含量、光合和蒸腾生理等土壤和植被状态,为模型的校验和验证提供数据。因此,系统研究棉花生长发育模型与农业物联网数据和农业遥感反演数据之间参数同化和模型耦合的方法原理,将多源农业监测技术所实时获取的棉花冠层状态信息与棉花生长发育模型进行有效耦合[104],综合棉花生长发育模型在点上、遥感观测在面上的优势,提升模型应用的时空范围是未来棉花生长发育模型研究的趋势之一[16]。4.3 棉花生长发育模型的推广和应用
棉花生长发育模型输入参数多、操作难度大的特点导致其在我国棉花生产实际中的应用和推广水平较低。随着物联网、大数据、云计算、人工智能等新一代信息技术在我国“三农”领域中的快速发展和广泛应用[105],为棉花生长发育模型的推广和应用提供了新的契机。基于新一代信息技术对现有棉花生长发育模型进行适宜性改进和提升,使其与棉田大数据采集网络相耦合,实现棉花生长发育模型的参数同化和精度提升,为棉花生产管理的水、肥、药智能决策和精准施用服务;与农机具作业参数和农机具作业质量监测技术相耦合,为棉田农机具作业质量提供数据监控与智能管理等大数据智能分析服务,提升农机具作业质量和运维管理能力;与棉花安全理论、经济预警理论和国内外棉花市场特点相融合,实现棉花市场预警和价格预测服务;与土壤质量环境评价和环境污染评价模型相耦合,阐述棉花生产措施(连作、地膜、脱叶剂等)对生态环境的影响评价等,是棉花生长发育模型研究的趋势之一。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
[本文引用: 11]
[本文引用: 3]
[本文引用: 3]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 5]
[本文引用: 6]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 2]
[本文引用: 6]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17812886 [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 6]
[本文引用: 1]
DOI:10.13031/2013.24082URL [本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
DOI:10.1007/s11427-012-4425-5URLPMID:23314868 [本文引用: 1]

Grass carp (Ctenopharyngodon idella) is an important species of freshwater aquaculture fish in China. However, grass carp reovirus (GCRV) can cause fatal hemorrhagic disease in yearling populations. Until now, a strategy to define the antigenic capacity of the virus's structural proteins for preparing an effective vaccine has not been available. In this study, some single-chain variable fragment antibodies (scFv), which could specifically recognize grass carp IgM, were selected from a constructed mouse naive antibody phage display cDNA library. The identified scFv C1B3 clone was shown to possess relatively higher specific binding activity to grass carp IgM. Furthermore, ELISA analysis indicated that the IgM level in serum from virus-infected grass carp was more than two times higher than that of the control group at 5-7 days post infection. Moreover, Western blot analysis demonstrated that the outer capsid protein VP7 has a specific immuno-binding-reaction with the serum IgM from virus-infected grass carp. Our results suggest that VP7 can induce a stronger immune response in grass carp than the other GCRV structural proteins, which implies that VP7 protein could be used as a preferred immunogen for vaccine design.
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ecolmodel.2012.06.015URL [本文引用: 1]
DOI:10.1016/S2095-3119(13)60206-3URL [本文引用: 1]
Crop performance is determined by the combined effects of the genotype of the crop and the environmental conditions of the production system. This study was undertaken to develop a dynamic model for simulating environmental (temperature and solar radiation) and N supply effects on fiber fineness, maturity and micronaire. Three different experiments involving genotypes, sowing dates, and N fertilization rates were conducted to support model development and model evaluation. The growth and development duration of fiber fineness, maturity, and micronaire were scaled by using physiological development time of secondary wall synthesis (PDTSWSP), which was determined based on the constant ratio of SWSP/BMP. PTP (product of relative thermal effectiveness (RTE) and photosynthetically active radiation (PAR), MJ m(-2)) and subtending leaf N content per unit area (N-A, g m(-2)) and critical subtending leaf N content per unit area (CNA, g m(-2)) of cotton boll were calculated or simulated to evaluate effects of temperature and radiation, and N supply. Besides, the interactions among temperature, radiation and N supply were also explained by piecewise function. The overall performance of the model was calibrated and validated with independent data sets from three field experiments with two sowing dates, three or five flowering dates and three or four N fertilization rates for three subsequent years (2005, 2007, and 2009) at three ecological locations. The average RMSE and RE for fiber fineness, maturity, and micronaire predictions were 372 m g(-1) and 5.0%, 0.11 m g(-1) and 11.4%, 0.3 mg(-1) and 12.3%, respectively, indicating a good fit between the simulated and observed data. It appears that the model can give a reliable prediction for fiber fineness, maturity and micronaire formation under various growing conditions.
[本文引用: 5]
[本文引用: 5]
[本文引用: 2]
URL [本文引用: 3]
URL [本文引用: 3]
DOI:10.1016/0022-5193(68)90080-5URLPMID:5659072 [本文引用: 2]
[本文引用: 1]
URL [本文引用: 1]
DOI:10.1093/aob/mcm316URLPMID:18184646 [本文引用: 2]
BACKGROUND AND AIMS: Cotton shows a marked plasticity vs. density in terms of branch development and geometry, internode elongation and leaf expansion. This paper proposes interpretations for observed plasticity in terms of light quantity and quality. METHODS: 3-D virtual plants were reconstructed from field observations and 3-D digitization and were used to simulate the light regime in cotton stands of different densities. KEY RESULTS: All densities showed the same linear relationship between LAI and the sum of light intercepted by the canopy, from seedling emergence up to flowering. Simulated R : FR ratio profiles can very likely explain (1) the longer first internodes on main stem and branches and (2) the azimuthal re-orientation of branches toward the inter-row. CONCLUSIONS: Simulation tools were used to analyse plant plasticity in terms of light quantity and quality. The methodology applied here at the stand scale will now be continued at the plant scale to further strengthen the above hypotheses.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1093/aob/mct309URLPMID:24489020 [本文引用: 2]
BACKGROUND AND AIMS: Cotton (Gossypium hirsutum) has indeterminate growth. The growth regulator mepiquat chloride (MC) is used worldwide to restrict vegetative growth and promote boll formation and yield. The effects of MC are modulated by complex interactions with growing conditions (nutrients, weather) and plant population density, and as a result the effects on plant form are not fully understood and are difficult to predict. The use of MC is thus hard to optimize. METHODS: To explore crop responses to plant density and MC, a functional-structural plant model (FSPM) for cotton (named CottonXL) was designed. The model was calibrated using 1 year's field data, and validated by using two additional years of detailed experimental data on the effects of MC and plant density in stands of pure cotton and in intercrops of cotton with wheat. CottonXL simulates development of leaf and fruits (square, flower and boll), plant height and branching. Crop development is driven by thermal time, population density, MC application, and topping of the main stem and branches. KEY RESULTS: Validation of the model showed good correspondence between simulated and observed values for leaf area index with an overall root-mean-square error of 0.50 m(2) m(-2), and with an overall prediction error of less than 10% for number of bolls, plant height, number of fruit branches and number of phytomers. Canopy structure became more compact with the decrease of leaf area index and internode length due to the application of MC. Moreover, MC did not have a substantial effect on boll density but increased lint yield at higher densities. CONCLUSIONS: The model satisfactorily represents the effects of agronomic measures on cotton plant structure. It can be used to identify optimal agronomic management of cotton to achieve optimal plant structure for maximum yield under varying environmental conditions.
[本文引用: 3]
[本文引用: 3]
URL [本文引用: 1]
为确定植物生长可视化建模的一般方法,在总结国内外相关研究成果的基础上,归纳了基于结构-功能反馈机制的植物生长可视化建模的步骤,研究了建模过程中涉及的植物形态结构描述模型、结构模型与功能模型的结合、可视化模型以及模型的有效性检验等关键性技术问题,最后以具体作物生长可视化建模为例阐述了该技术的应用,并对关键性技术问题进行了讨论。研究结果表明,基于结构-功能反馈机制的作物生长模型,不但能可视化模拟不同环境因素下作物的生长发育状况,而且可在作物理想株型培育、群体产量预测、栽培管理等领域发挥实际作用。
URL [本文引用: 1]
为确定植物生长可视化建模的一般方法,在总结国内外相关研究成果的基础上,归纳了基于结构-功能反馈机制的植物生长可视化建模的步骤,研究了建模过程中涉及的植物形态结构描述模型、结构模型与功能模型的结合、可视化模型以及模型的有效性检验等关键性技术问题,最后以具体作物生长可视化建模为例阐述了该技术的应用,并对关键性技术问题进行了讨论。研究结果表明,基于结构-功能反馈机制的作物生长模型,不但能可视化模拟不同环境因素下作物的生长发育状况,而且可在作物理想株型培育、群体产量预测、栽培管理等领域发挥实际作用。
[本文引用: 2]
URL [本文引用: 2]
采用GREENLAB植物功能-结构模型的原理构建了棉花根系模型.该模型以根系基本生长单元为基础,模拟了棉花根系的拓扑结构;根据不同类型的根个体获取生物量能力的不同,通过模拟不同根个体(库)对植株地上部分配给根系的生物量(源)的竞争,实现了生物量在根系中的分配;根据异速生长规则实现了根个体几何结构计算,从而模拟出根个体的轴向生长、径向生长.最后,根据根个体空间伸展规则与空间Voxel元技术,实现了棉花根系的空间结构的虚拟.
URL [本文引用: 2]
采用GREENLAB植物功能-结构模型的原理构建了棉花根系模型.该模型以根系基本生长单元为基础,模拟了棉花根系的拓扑结构;根据不同类型的根个体获取生物量能力的不同,通过模拟不同根个体(库)对植株地上部分配给根系的生物量(源)的竞争,实现了生物量在根系中的分配;根据异速生长规则实现了根个体几何结构计算,从而模拟出根个体的轴向生长、径向生长.最后,根据根个体空间伸展规则与空间Voxel元技术,实现了棉花根系的空间结构的虚拟.
URLPMID:19995824 [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1469-8137.2005.01330.xURL [本文引用: 1]
DOI:10.1093/aob/mcj033URLPMID:16390847 [本文引用: 1]
BACKGROUND AND AIMS: There are three reasons for the increasing demand for crop models that build the plant on the basis of architectural principles and organogenetic processes: (1) realistic concepts for developing new crops need to be guided by such models; (2) there is an increasing interest in crop phenotypic plasticity, based on variable architecture and morphology; and (3) engineering of mechanized cropping systems requires information on crop architecture. The functional-structural model GREENLAB was recently presented that simulates resource-dependent plasticity of plant architecture. This study introduces a new methodology for crop parameter optimization against measured data called multi-fitting, validates the calibrated model for maize with independent field data, and describes a technique for 3D visualization of outputs. METHODS: Maize was grown near Beijing during the 2000, 2001 and 2003 (two sowing dates) summer seasons in a block design with four to five replications. Detailed morphological and topological observations were made on the plant architecture throughout the development of the four crops. Data obtained in 2000 was used to establish target files for parameter optimization using the generalized least square method, and parameter accuracy was evaluated by coefficient of variance. In situ plant digitization was used to establish 3D symbol files for organs that were then used to translate model outputs directly into 3D representations for each time step of model execution. KEY RESULTS AND CONCLUSIONS: Multi-fitting against several target files obtained at different growth stages gave better parameter accuracy than single fitting at maturity only, and permitted extracting generic organ expansion kinetics from the static observations. The 2000 model gave excellent predictions of plant architecture and vegetative growth for the other three seasons having different temperature regimes, but predictions of inter-seasonal variability of biomass partitioning during grain filling were less accurate. This was probably due to insufficient consideration of processes governing cob sink size and terminal leaf senescence. Further perspectives for model improvement are discussed.
DOI:10.1071/FP08052URLPMID:32688828 [本文引用: 2]
The programming language XL ('eXtended L-system language') is an extension of Java, which supports the specification and execution of relational growth grammars, a variant of parallel graph grammars. XL is a powerful generalisation of the well-known L-system approach to functional-structural plant modelling. Some features of XL are discussed that are particularly useful for combining structure and function and for querying plant architectural data, and an exemplary functional-structural plant model of young beech trees is presented that is implemented in XL and includes PAR distribution, assimilate allocation and morphological plasticity. Together with a simpler model of spruce trees, this beech model is included in a virtual landscape with a mixed-species forest stand where competition for light occurs. The open-source platform GroIMP was used for the complete model development process and for visualising the results.
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1371/journal.pone.0197284URLPMID:29750816 [本文引用: 1]
Cotton root growth can be affected by different nitrogen fertilizer rates. The objective of the present study was to quantify the effects of nitrogen fertilization rate on cotton root growth and distribution using minirhizotron and soil coring methods. A secondary objective was to evaluate the minirhizotron method as a tool for determining nitrogen application rates using the root distribution as an index. This study was conducted on a Bt cotton cultivar (Jimian 958) under four nitrogen fertilization rates, i.e., 0, 120, 240 and 480 kg ha(-1) (control, low, moderate and high levels, respectively), in the Yellow River basin of China from 2013-2015. The sampling process, details of each method as well as the root morphology and root distribution were measured. The operational processes, time and labor needed for the soil core method were all greater than those for the minirhizotron method. The total root length density and the length density in most soil layers, especially in the upper soil layers, first increased but then decreased as nitrogen fertilization increased, and the same trend was observed for both methods. Compared with N0, the total root length density under moderate nitrogen fertilization by the soil coring method increased by more than 94.82%, in 2014 and 61.11% in 2015; while by the minirhizotron method the corresponding values were 28.24% in 2014 and 57.47%, in 2015. Most roots were distributed in the shallow soil layers (0-60 cm) in each method. However, the root distribution with the soil coring method (>73.11%) was greater than that with the minirhizotron method (>47.07%). The correlations between the root morphology indexes of shallow soil depth measured using the two methods were generally significant, with correlative coefficients greater than 0.334. We concluded that the minirhizotron method could be used for cotton root analysis and most cotton roots distributed in upper soil layers (0-60cm). In addition, a moderate nitrogen rate (240 kg ha-1) could increase root growth, especially in the shallow soil layers. The differences observed with the minirhizotron method were clearer than those observed with the soil coring method.
[本文引用: 1]
DOI:10.1038/s41477-020-0625-3URLPMID:32296143 [本文引用: 1]
Predicting the consequences of manipulating genotype (G) and agronomic management (M) on agricultural ecosystem performances under future environmental (E) conditions remains a challenge. Crop modelling has the potential to enable society to assess the efficacy of G x M technologies to mitigate and adapt crop production systems to climate change. Despite recent achievements, dedicated research to develop and improve modelling capabilities from gene to global scales is needed to provide guidance on designing G x M adaptation strategies with full consideration of their impacts on both crop productivity and ecosystem sustainability under varying climatic conditions. Opportunities to advance the multiscale crop modelling framework include representing crop genetic traits, interfacing crop models with large-scale models, improving the representation of physiological responses to climate change and management practices, closing data gaps and harnessing multisource data to improve model predictability and enable identification of emergent relationships. A fundamental challenge in multiscale prediction is the balance between process details required to assess the intervention and predictability of the system at the scales feasible to measure the impact. An advanced multiscale crop modelling framework will enable a gene-to-farm design of resilient and sustainable crop production systems under a changing climate at regional-to-global scales.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}