 ,1, 莫凯琴1, 刘璐1, 李萍芳1, 金晨钟,1, 杨芳,2
,1, 莫凯琴1, 刘璐1, 李萍芳1, 金晨钟,1, 杨芳,2Effect of Overexpression of OsRRK1 Gene on Rice Leaf Development
MA YinHua,1, MO KaiQin1, LIU Lu1, LI PingFang1, JIN ChenZhong,1, YANG Fang,2通讯作者:
责任编辑: 李莉
收稿日期:2020-07-16接受日期:2020-08-26网络出版日期:2021-03-01
| 基金资助: |
Received:2020-07-16Accepted:2020-08-26Online:2021-03-01
作者简介 About authors
马银花,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (3073KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
马银花, 莫凯琴, 刘璐, 李萍芳, 金晨钟, 杨芳. 过量表达OsRRK1对水稻叶片发育的影响[J]. 中国农业科学, 2021, 54(5): 877-886 doi:10.3864/j.issn.0578-1752.2021.05.001
MA YinHua, MO KaiQin, LIU Lu, LI PingFang, JIN ChenZhong, YANG Fang.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】在水稻中,叶片是植物形态建成的一个重要指标,与植物株型的形成密切相关[1]。叶片形态的改良,直接或间接的影响到作物光合作用,合适的叶片卷曲导致叶片直立,可提高光合效率,加速干物质积累和增加谷物产量,是当前水稻高产育种关注的重点[2,3]。而在水稻生产中,开展叶形基因如何调节叶片发育的研究,对水稻叶形的改良和理想株型的育种具有十分重要的意义。【前人研究进展】水稻叶片是植株进行光合作用的主要场所,叶片卷曲是叶片性状中的一个重要组成部分[4]。水稻卷叶形成的主要因素是叶片中泡状细胞的膨胀程度及其渗透压,生理性逆境(缺水和盐胁迫等)都会使叶片发生卷曲,但这种条件下的卷叶性状是可逆的且不能遗传,而由基因控制的卷叶性状一般能够遗传且表型稳定[2]。在水稻中的卷叶类型可简单分为3种:正卷、反卷和扭曲,目前已报道了超过35个叶卷曲突变体,且大部分研究表明都与泡状细胞生长的状况与叶片卷曲密切相关。在已克隆的水稻卷叶基因中,HU等[5]研究NRL1(the narrow and rolled leaf 1)突变以后由于泡状细胞数目变少,叶片发生正卷,并且叶片也出现变窄现象。LI等[6]研究过表达ACL1(abaxially curled leaf 1)增加泡状细胞的数目导致反卷。FANG等[7]研究发现RL14(rolling-leaf 14)通过影响叶片次生细胞壁的形成影响叶片卷曲,突变后引起正卷。ZOU等[8]研究发现Roc5(outermost cell-specific gene 5)突变后由于增加泡状细胞的数目和面积导致反卷。XU等[9]研究发现过表达OsZHD1可以增加泡状细胞的数目和影响排列导致反卷。HIBARA等[10]研究发现ADL1(adaxialized leaf 1)突变以后由于表皮细胞在近轴面和远轴面都变大,并且远轴面与泡状细胞无法分开而导致叶片反卷。李聪等[11]发现了一个新LBD家族基因OsLBD3-7可以通过调节泡状细胞,影响水稻叶片卷曲。FUJINO等[12]研究NAL7突变以后由于泡状细胞变小引起叶片卷曲,同时叶片还出现变窄现象。SLL1编码的是一个SHAQKYF家族的转录因子,属于KANADI家族,突变以后由于远轴面厚壁细胞的缺失从而导致叶片卷曲,且卷曲程度很高[13]。SRL1突变以后也是通过影响泡状细胞的形成进而调控叶片卷曲[14]。受体类激酶(receptor-like kinases,RLKs)在植物生长发育以及生物和非生物的胁迫过程中发挥着重要作用[15],有一类RLKs缺乏细胞外受体结构域,被称为胞质类受体激酶(receptor-like cytoplasmic kinases,RLCKs),RLCKs是一类无胞外结构域和跨膜结构域的蛋白激酶,只含有胞内激酶结构域,是RLK蛋白激酶超家族中一类特殊的蛋白家族。研究发现这类蛋白在拟南芥中有200个成员,而在水稻中有379个成员[16],在水稻中,关于RLCKs基因的研究很少,仅有OsRLCK55、OsRLCK185、OsRLCK57、OsRLCK102、OsRLCK107、OsRLCK118、OsRLCK176等基因被报道,且它们都属于RLCKⅦ亚家族[17,18,19,20]。目前,对RLCKⅥ亚家族的成员的研究还不够深入,在拟南芥中只有研究了AtRBK1、AtRBK2、AtRRK1和AtRRK2的功能。AtRBK1和AtRBK2可以与Rop GTPase直接互相作用。AtRBK1和AtRBK2参与致病疫霉菌和葡萄孢菌的防御反应[21]。AtRRK1和AtRRK2调节拟南芥的生长发育以及防御反应[22]。OsRRK1是个多效性基因,并且是水稻类蛋白受体激酶RLCKⅥ家族成员首个被报道的蛋白[23],进一步明确OsRRK1对水稻叶片发育的影响,可为水稻育种和基因功能的研究提供借鉴。【本研究切入点】RLCKⅥ家族基因的功能在其他物种中有过报道,但是对水稻中RLCKⅥ家族成员的功能研究鲜有报道。前期研究发现,作为RLCKⅥ家族成员之一的OsRRK1参与水稻的生长发育和对褐飞虱的抗性反应。【拟解决的关键问题】本研究通过构建OsRRK1超量表达载体,利用农杆菌介导的遗传转化方法,转化生育期较短易于转化的粳稻品种合江19,得到转基因植株。通过对转基因植株的分子鉴定、叶片形态观察、叶绿素含量测定,明确OsRRK1对水稻叶片生长发育的影响。1 材料与方法
试验于2014年9月到2019年10月在武汉大学杂交水稻国家重点实验室/湖南人文科技学院湖南省农田杂草防控技术与应用协同创新中心完成。1.1 供试材料
大肠杆菌(Escherichia coli)菌株TOP10和农杆菌(Agrobacterium tumefaciens)菌株EHA105由武汉大学杂交水稻国家重点实验室何光存教授课题组提供;水稻材料粳稻品种合江19(Oryza sativa L. subsp. japonica cv. ‘Hejiang 19’)购自国家种质资源库;合江19为一早熟水稻品种,因其生育期短,在本研究中用作水稻转基因研究的受体材料;扩增序列用的模板来源于籼稻品种Kasalath(Oryza sativa L. subsp. indica cv. Kasalath)。载体pCXUN由美国Ohio State University王国梁教授提供。1.2 OsRRK1超量表达载体的构建
依据水稻基因组数据库公布的目标基因序列,通过Primer5软件设计用于基因扩增的上下游引物,即OE-OsRRK1-F和OE-OsRRK1-R,提取水稻品种Kasalath的基因组DNA作为模板,通过PCR扩增获得目的片段,将扩增的DNA片段与用相同的XcmⅠ酶消化的pCXUN载体(含有玉米泛素ubiquitin启动子以及nos终止子)相连接,然后转化到大肠杆菌(Escherichia coli)菌株TOP10中,通过测序将连接正确的菌株提取质粒,即为构建成功的过量表达载体,命名为OE-OsRRK1。1.3 农杆菌介导的对粳稻品种合江19的遗传转化
将构建成功的载体提取质粒,然后电转化进入农杆菌EHA105的感受态细胞备用。水稻遗传转化按照TOKI等[24]方法进行,水稻种子经过诱导-继代-前培养→农杆菌培养和悬浮→感染和共培养→农杆菌去除,之后将转化后的愈伤组织接种于含50 mg·L-1潮霉素和400 mg·L-1羧苄青霉素的N6培养基上[25],筛选抗性愈伤组织。在32℃持续光照培养2周后,将生长旺盛的胚性愈伤组织转移到含50 mg·L-1潮霉素B和250 mg·L-1羧苄青霉素的再生培养基上持续光照诱导芽的分化。2周后将幼苗转移至含50 mg·L-1潮霉素B和200 mg·L-1羧苄青霉素的生根培养基上诱导生根[26]。获得完整植株后,炼苗7—10 d,移至温室培养。1.4 PCR检测阳性植株
以转基因植株叶片提取的DNA为模板,运用潮霉素基因引物(hyg-L和hyg-R)进行检测,或用载体序列设计的引物UBIS(F)/UBIA(R)进行扩增检测,所用的引物序列如表1所示。Table 1
表1
表1用于构建载体的引物
Table 1
| 引物Primer | 引物序列Primer sequence (5′-3′) | |
|---|---|---|
| OE-OsRRK1-F | ATGAGGCCTCTGTACCTGC | |
| OE-OsRRK1-R | CTAATTGCTCAAAGATGATGAGC | |
| UBIS(F) | TGTTTCTTTTGTCGATGCTCACCC | |
| UBIA(R) | TTCTATCGCGGCTTAACGTAATTCA | |
| hyg-L | GCTCCATACAAGCCAACCAC | |
| hyg-R | GAAAAAGCCTGAACTCACCG | |
新窗口打开|下载CSV
1.5 转基因植株的Southern blot分析
利用CTAB法抽提水稻大量基因组DNA,其步骤参考CHEN等[27]方法进行:DNA限制性内切酶消化→凝胶电泳与Southern blot转膜→膜的预杂交→杂交→探针标记→洗膜→压片→放射自显影。1.6 转基因植株的Northern blot分析
采用Trizol法提取转基因水稻叶片的总RNA,参考CHEN等[27]方法进行:制备MOPS/甲醛变性凝胶进行RNA电泳→转膜→膜的预杂交→杂交→探针标记→洗膜→压片→放射自显影。1.7 转基因植株的荧光定量PCR(qRT-PCR)分析
将转基因植株T2代的纯合阳性株系进行播种、待长至抽穗期,取水稻剑叶,每个转基因株系取3株。采用Trizol法提取转基因水稻叶片的总RNA。RNA的反转录按RevertAidTM First Strand cDNA Synthesis Kit(Fermentas)的操作步骤进行。PCR反应体系为DNase/RNase-Free ddH2O (TIANGEN)2.9 μL、2×Super mix 4 μL、Primers (5 mmol·L-1) 0.6 μL和cDNA template 0.5 μL。
采用Bio-Rad CFX Manager分析荧光定量PCR分析结果,首先对其溶解曲线,QC(质控)等进行分析,去除不达标数据,之后用软件自带Gene Study功能按(1+E)—tudy算法分析目的基因的表达。
1.8 叶片卷曲指数的测定
选取T2代纯合植株抽穗期的剑叶进行测量,展开叶片,测量其最宽处距离,即为叶片最大宽度(maximum leaf width,Lw);测量同位置叶片两边缘间自然状态下距离,即为叶片自然状态的宽度(natural leaf width,Ln)。计算出叶片卷曲指数(leaf rolling index,LRI),LRI(%)=(Lw-Ln)/Lw×100%;每个转基因的独立株系随机选取20株进行测量,重复3次,取平均值。差异显著性用t检验进行分析。1.9 石蜡切片和苯胺蓝染色
在水稻抽穗期时,选取同样条件下生长于大田中的OE-OsRRK1植株和WT植株的剑叶。取样位置为正中间最宽处,进行材料固定→脱水→透明→渗蜡→包埋→修块和切片→脱蜡和复水→染色→镜检等基本试验操作。1.10 叶绿素含量测定
取新鲜、干净的野生水稻叶片和过量表达OsRRK1水稻植株叶片,首先对叶绿素测定计进行校准,之后将叶片置于测定计夹片处,接着2个LED灯源同时发出红光和红外线,2种光穿透叶片打到接收器上,被转换为电信号。通过这两种波长范围内的透光系数来确定叶片当前叶绿素的相对含量(SPAD值)。随机选取20株的剑叶进行测量,结果用20次的数据平均值±标准差表示(n=20)。差异显著性用t检验进行分析。2 结果
2.1 OE-OsRRK1载体的构建
以基因组DNA为模板扩增含有OsRRK1的ORF序列(图1-A)。获得长度为1 179 bp的片段。将其连接至转化载体PCXUN。通过酶切验证,表明OE-OsRRK1载体构建成功(图1)。图1
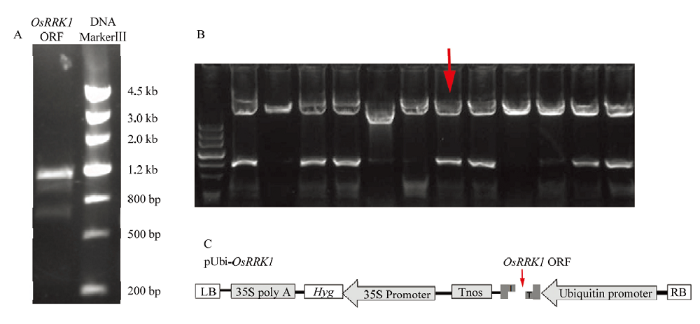 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1OsRRK1超量表达载体的构建
A:OsRRK1全长ORF的扩增;B:重组载体的酶切鉴定;C:构建成功的OE-OsRRK1载体
Fig. 1Construction of OsRRK1 overexpression vector
A: Amplification of OsRRK1 full-length ORF; B: Restriction identification of recombinant vector; C: Successfully constructed OE-OsRRK1 vector
2.2 转基因植株的获得
采用农杆菌介导法,成功将OE-OsRRK1载体导入水稻合江19的基因组中。从农杆菌侵染的水稻愈伤组织块中,获得了抗性愈伤组织块。将生长旺盛的胚性愈伤组织诱导发芽生根,获得T0代转基因再生植株47株,用于进一步的转基因分子验证。2.3 OE-OsRRK1转基因植株的分子鉴定
将获得的OE-OsRRK1植株作为材料,用外源潮霉素Hyg设计引物,通过PCR扩增进行初步鉴定(图2-A),获得OE-OsRRK1阳性转化植株44株。图2
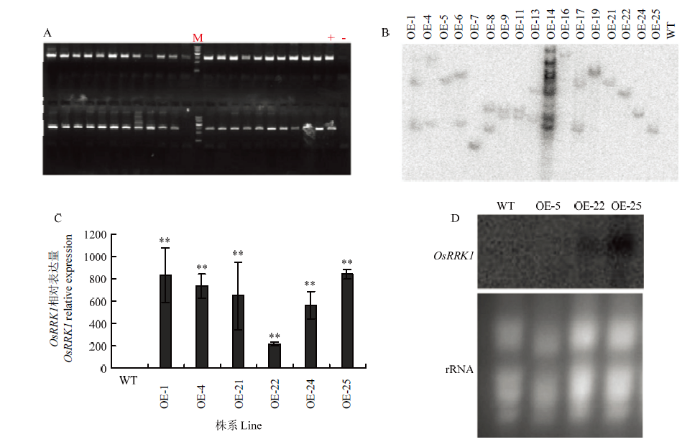 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2 OsRRK1过量表达(OE-OsRRK1)转基因水稻植株的分子鉴定
A:OE-OsRRK1转基因植株中目的基因的PCR检测;M:MarkerIII;+:阳性质粒对照;-:未转基因水稻阴性对照;其余为部分OE-OsRRK1转基因水稻植株。B:OE-OsRRK1转基因植株的Southern blot分析。C:OE-OsRRK1转基因植株的实时荧光定量PCR分析,**在0.01水平的差异显著性。下同。D:OE-OsRRK1转基因植株的Northern blot分析。WT:野生型合江19;OE-1—OE-25:部分OE-OsRRK1转基因水稻植株,其中OE-5、OE-7、OE-16、OE-19、OE-21、OE-22、OE-24和OE-25为单拷贝株系,OE-5:单拷贝表达量程度低的株系;OE-22:单拷贝表达量程度中等的株系;OE-25:单拷贝表达量程度高的株系
Fig. 2Molecular identification of transgenic rice plants with OsRRK1 overexpression (OE-OsRRK1)
A: PCR detection of target genes in OE-OsRRK1 transgenic plants; M: MarkerIII; +: Positive plasmid control; -: Non-transgenic rice negative control; The rest are some OE-OsRRK1 transgenic rice plants. B: Southern blot analysis of OE-OsRRK1 transgenic plants. C: Fluorescence quantitative PCR analysis of OE-OsRRK1 transgenic plants; ** t-test, P<0.01. The same as below. D: Northern blot analysis of OE-OsRRK1 transgenic plants; WT: Wild-type Hejiang 19; OE-1-OE-25: Part of OE-OsRRK1 transgenic rice plants, in which OE-5, OE-7, OE-16, OE-19, OE-21, OE-22, OE-24, OE-25 were single copy strains; OE-5: Strains with single copy of low expression level; OE-22: Single copy of medium expression level of strains; OE-25: Strains with single copy of high expression strains
从第1代OE-OsRRK1的阳性株系随机选出17个进行Southern blot分析。利用超霉素引物扩增的产物为探针,通过限制性内切酶的消化、凝胶电泳、转膜、杂交、洗膜和自显影等过程,确定每个转基因株系中外源基因转入的拷贝数(图2-B),其中OE-5、OE-7、OE-16、OE-19、OE-21、OE-22、OE-24和OE-25为单拷贝株系,其他为多拷贝。为了研究OE-OsRRK1转基因植株中OsRRK1在RNA水平的表达情况,选取抽穗期的剑叶进行取样提RNA进行qRT-PCR分析(图2-C)。OsRRK1在不同的转基因株系中都出现过量表达的情况,但过表达的程度不同,OE-1、OE-4和OE-25株系超量表达程度高,OE-5株系超量表达程度很低,OE-21、OE-24和OE-22株系超量表达程度居中,OsRRK1的表达程度高低跟拷贝数没有直接关系,多拷贝的株系也不一定就比单拷贝的株系表达程度高,而几份单拷贝的株系表达程度也高低不等。
为进一步鉴定OE-OsRRK1转基因植株中OsRRK1的RNA水平表达情况,选择抽穗期的剑叶提取RNA,进行Northern blot分析,选取OE-5、OE-22和OE-25 3个单拷贝转基因株系和WT对照。由图2-D可知,OsRRK1在WT和OE-5中几乎检测不到,在OE-22中信号较弱,而在OE-25中信号很强(图2-D),这与qRT-PCR分析结果一致。
2.4 OE-OsRRK1转基因植株叶片卷曲分析
过表达OsRRK1的水稻叶片会出现卷曲,选取单拷贝并且表达程度最高的OE-25作为取样材料,可见OE-25相对于WT明显的叶片卷曲(图3-A)。通过计算叶片的卷曲指数(图3-B),可知WT对照的卷曲为0,表达量程度很低的OE-5也是0,表达量程度居中的OE-21、OE-24和OE-22株系,叶片卷曲指数也是居中,表达量程度高的OE-1、OE-4和OE-25株系叶片卷曲指数也高,卷曲的程度与OsRRK1的表达量有关,表达量越高卷曲程度越高,与基因的拷贝数无关。图3
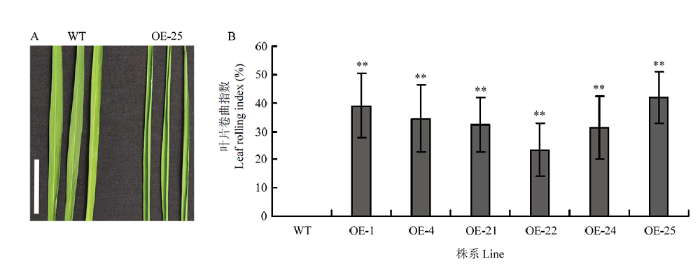 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3WT和OE-OsRRK1植株的叶片表型
A:WT和OE-25植株在抽穗期的剑叶,Bars=5 cm;B:WT和OE-OsRRK1转基因植株的叶片卷曲指数
Fig.3Leaf phenotypes of WT and OE-OsRRK1 transgenic plants
A: The flag leaves of WT and OE-25 plants at heading stage, Bars=5 cm; B: Leaf curl index of WT and OE-OsRRK1 transgenic plants
为了分析OE-OsRRK1转基因植株叶片卷曲的原因,选取转基因水稻和对照合江19抽穗期的剑叶进行石蜡切片,观察水稻横切后各细胞的变化(图4-A),结果发现,OE-OsRRK1转基因植株叶片横切中不管是中轴、大维管束还是小维管束两边的泡状细胞数量减少,面积变小,并且表达量最高的OE-25的泡状细胞退化更明显,面积最小。为了对泡状细胞的数量进行量化,将一片水稻叶片横切后所有的泡状细胞统计,对照中泡状细胞数目均值约为4.6个,而转基因株系中泡状细胞数目都小于4.6,跟对照相比具有极显著差异,并且表达程度最高的OE-25株系泡状细胞数量均值只有约3.4个(图4-B)。同时用Image J软件对泡状细胞的面积进行统计,转基因株系与对照相比明显变小,并且具有极显著差异(图4-C)。
图4
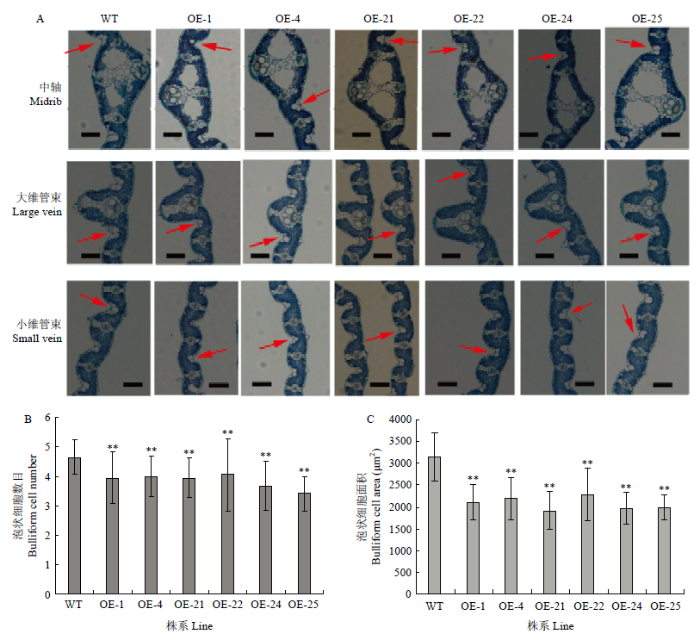 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4OE-OsRRK1植株叶片泡状细胞的数目和面积
A:WT和OE-OsRRK1植株中的剑叶的横切图(Bars=100 μm);红色箭头指示泡状细胞;B:WT和OE-OsRRK1植株叶片的泡状细胞的数目的统计;C:WT和OE-OsRRK1植株中的剑叶细胞的面积。WT:野生型合江19;OE-1、OE-4、OE-21、OE-22、OE-24和OE-25:OE-OsRRK1转基因植株
Fig. 4Bulliform cell number and size in the leaves of OE-OsRRK1 plants
A: Leaf cross-section of OE-OsRRK1 plants (Bars=100 μm); the red arrow indicates the bulliform cell; B: The bulliform cell number in WT and OE-OsRRK1 plants; C: The bulliform cell area in WT and OE-OsRRK1 plants. WT: Wildtype hejiang19; OE-1, OE-4, OE-21, OE-22, OE-24, and OE-25: OE-OsRRK1 transgenic plants
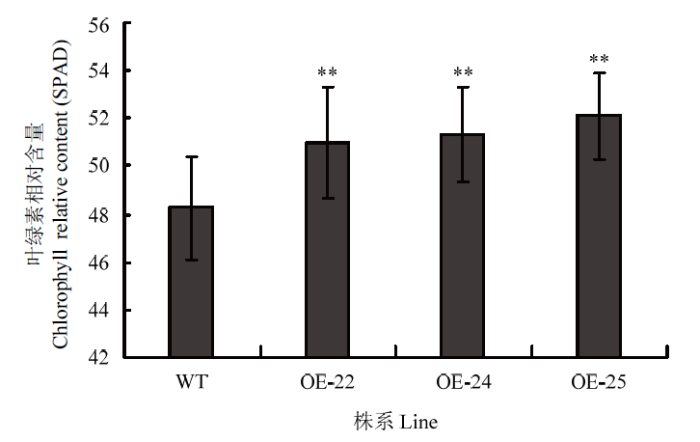
2.5 OE-OsRRK1转基因植株叶片叶绿素含量分析
通过分析OE-OsRRK1转基因植株叶片和WT叶绿素含量的变化(图5),发现OE-OsRRK1植株的叶绿素含量要比WT植株的高,具有极显著性差异。推测过量表达OsRRK1会导致叶绿素含量增高。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5WT和OE-OsRRK1植株的叶绿素相对含量统计
WT:野生型合江19;OE-22、OE-24和OE-25:OE-OsRRK1转基因植株
Fig. 5Chlorophyll relative content of WT and OE-OsRRK1 plants
WT: Wild type of Hejiang19; OE-22, OE-24, and OE-25: OE-OsRRK1 transgenic plants
3 讨论
转基因植物中外源基因拷贝数是影响目的基因表达水平和遗传稳定性的重要因素[28]。本研究构建了OsRRK1过量表达的载体、通过农杆菌介导法转到粳稻品种合江19基因组中,获得OE-OsRRK1转基因植株。通过Southern blot分析转基因株系外源基因插入的拷贝数;qRT-PCR和Northern blot分析OsRRK1在转基因株系中的表达水平,发现OsRRK1的表达程度高低跟拷贝数没有直接关系,多拷贝的株系也不一定就比单拷贝的株系表达程度高,而几份单拷贝的株系表达程度也高低不等。在对高产水稻的理想株型研究中,卷叶性状的研究被广泛关注,因为叶片适度卷曲导致叶片直立不披垂,可提高光合效率,加速干物质积累和增加谷物产量[3],所以适度卷叶是实现水稻育种潜在产量突破的理想性状。目前,关于卷叶基因研究中最多的就是通过影响泡状细胞的发育来调控叶卷的这类基因。本研究中OE-OsRRK1转基因植株与对照相比叶片卷曲明显,泡状细胞数目更少、面积更小,退化明显,其他各细胞无明显变化,说明过表达OsRRK1可以通过减少泡状细胞的数目和影响泡状细胞面积大小导致水稻叶片正卷,目前这种调控的具体机制以及参与的信号转导途径还有待进一步探究。此外,有研究表明水稻的高产与其生长周期的光合作用密不可分,光合作用是产量形成的基础,作物的干物质生产直接来源于光合作用[29,30]。叶绿素是水稻叶片进行光合作用的重要物质之一,其含量是决定作物光合效率和生物量或产量的重要因素,而叶片叶绿素含量的高低表征水稻光合效率与生长状况等重要生长指标,其与作物产量呈正相关,因此适量增高叶绿素含量有利于提高产量[31,32,33,34,35]。在OE-OsRRK1转基因植株与野生水稻植株的叶绿素含量分析中发现3个OE-OsRRK1代表性株系的叶绿素含量均高于野生水稻植株,具有极显著差异。这表明由于水稻中OsRRK1表达量增多,使得OsRRK1表达量升高后可能促进了叶片中叶绿素的合成,导致叶绿素含量的增加。
4 结论
OsRRK1能够通过调控泡状细胞的发育影响OE-OsRRK1转基因植株叶片的卷曲,并且卷曲的程度与OsRRK1的呈正相关,可通过调控OsRRK1的表达量来调控转基因水稻叶片卷曲程度。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2013.00767URL [本文引用: 2]

叶片形态是水稻“理想株型”的重要组成部分,是当前水稻高产育种关注的重点。本文通过对已克隆多个叶形相关调控基因综述了水稻叶片形态(叶片卷曲度、倾角、披散程度以及叶片宽度)建成的分子遗传学研究进展。综合分析认为,水稻叶片的卷曲主要是通过卷叶基因调控叶片近轴/远轴间的发育、泡状细胞的发育及其膨胀和渗透压、厚壁组织的形成以及叶片角质层的发育等来实现。影响植株空间伸展姿态的叶倾角主要通过叶角基因调控油菜素内酯的信号传导来影响叶枕细胞的生长发育;唯一被克隆的影响叶片披垂度的披叶基因DL1是通过控制叶片中脉发育而改变叶片形态的;而窄叶基因则主要通过调控生长素的合成与极性运输、维管组织的发育和分布,影响叶片维管束数目及宽度。但到目前为止,所有已克隆的叶形调控基因间相互调控关系的研究还不够深入,还不能完整清晰地勾勒水稻叶形建成和发育的分子调控网络。因此,在已有的研究基础上更深入地探索水稻叶片形态建成的分子调控机制,对进一步构建相关的调控网络,塑造水稻理想株型具有重要意义。
DOI:10.3724/SP.J.1006.2013.00767URL [本文引用: 2]
叶片形态是水稻“理想株型”的重要组成部分,是当前水稻高产育种关注的重点。本文通过对已克隆多个叶形相关调控基因综述了水稻叶片形态(叶片卷曲度、倾角、披散程度以及叶片宽度)建成的分子遗传学研究进展。综合分析认为,水稻叶片的卷曲主要是通过卷叶基因调控叶片近轴/远轴间的发育、泡状细胞的发育及其膨胀和渗透压、厚壁组织的形成以及叶片角质层的发育等来实现。影响植株空间伸展姿态的叶倾角主要通过叶角基因调控油菜素内酯的信号传导来影响叶枕细胞的生长发育;唯一被克隆的影响叶片披垂度的披叶基因DL1是通过控制叶片中脉发育而改变叶片形态的;而窄叶基因则主要通过调控生长素的合成与极性运输、维管组织的发育和分布,影响叶片维管束数目及宽度。但到目前为止,所有已克隆的叶形调控基因间相互调控关系的研究还不够深入,还不能完整清晰地勾勒水稻叶形建成和发育的分子调控网络。因此,在已有的研究基础上更深入地探索水稻叶片形态建成的分子调控机制,对进一步构建相关的调控网络,塑造水稻理想株型具有重要意义。
URL [本文引用: 2]
以两个珍汕97B为遗传背景的卷叶近等基因系分别与明恢63所配的、叶型为半卷及平展的杂交组合为材料,进行栽培密度试验,对光合特性、物质生产与产量形成进行研究。结果表明,卷叶的单叶正面光合速率稍低于平展叶,而其背面光合速率稍高于平展叶,叶片中后期衰老程度,卷叶显著小于展叶;群体光合速率、群体干物质生产以及产
URL [本文引用: 2]
以两个珍汕97B为遗传背景的卷叶近等基因系分别与明恢63所配的、叶型为半卷及平展的杂交组合为材料,进行栽培密度试验,对光合特性、物质生产与产量形成进行研究。结果表明,卷叶的单叶正面光合速率稍低于平展叶,而其背面光合速率稍高于平展叶,叶片中后期衰老程度,卷叶显著小于展叶;群体光合速率、群体干物质生产以及产
DOI:10.11983/CBB17236URL [本文引用: 1]
卷叶性状是水稻(Oryza sativa)重要的育种指标之一。研究表明, 水稻叶片适度卷曲对植株的光合作用、株型及增产等均具有重要作用。该文综述了水稻叶片卷曲的相关研究进展, 对水稻叶片卷曲的细胞学形成机制和相关调控基因的分子机制进行了阐述, 以期深入阐明水稻叶片卷曲的细胞学和分子机制, 促进卷叶性状在水稻育种中的应用。
DOI:10.11983/CBB17236URL [本文引用: 1]
卷叶性状是水稻(Oryza sativa)重要的育种指标之一。研究表明, 水稻叶片适度卷曲对植株的光合作用、株型及增产等均具有重要作用。该文综述了水稻叶片卷曲的相关研究进展, 对水稻叶片卷曲的细胞学形成机制和相关调控基因的分子机制进行了阐述, 以期深入阐明水稻叶片卷曲的细胞学和分子机制, 促进卷叶性状在水稻育种中的应用。
URLPMID:20155303 [本文引用: 1]
URLPMID:20494951 [本文引用: 1]
URLPMID:22329407 [本文引用: 1]
DOI:10.1104/pp.111.176016URL [本文引用: 1]
Leaf rolling is considered an important agronomic trait in rice (Oryza sativa) breeding. To understand the molecular mechanism controlling leaf rolling, we screened a rice T-DNA insertion population and isolated the outcurved leaf1 (oul1) mutant showing abaxial leaf rolling. The phenotypes were caused by knockout of Rice outermost cell-specific gene5 (Roc5), an ortholog of the Arabidopsis (Arabidopsis thaliana) homeodomain leucine zipper class IV gene GLABRA2. Interestingly, overexpression of Roc5 led to adaxially rolled leaves, whereas cosuppression of Roc5 resulted in abaxial leaf rolling. Bulliform cell number and size increased in oul1 and Roc5 cosuppression plants but were reduced in Roc5-overexpressing lines. The data indicate that Roc5 negatively regulates bulliform cell fate and development. Gene expression profiling, quantitative polymerase chain reaction, and RNA interference (RNAi) analyses revealed that Protodermal Factor Like (PFL) was probably down-regulated in oul1. The mRNA level of PFL was increased in Roc5-overexpressing lines, and PFL-RNAi transgenic plants exhibit reversely rolling leaves by reason of increases of bulliform cell number and size, indicating that Roc5 may have a conserved function. These are, to our knowledge, the first functional data for a gene encoding a homeodomain leucine zipper class IV transcriptional factor in rice that modulates leaf rolling.
URLPMID:24385091 [本文引用: 1]
DOI:10.1016/j.ydbio.2009.07.042URLPMID:19665012 [本文引用: 1]
The adaxial-abaxial axis in leaf primordia is thought to be established first and is necessary for the expansion of the leaf lamina along the mediolateral axis. To understand axis information in leaf development, we isolated the adaxialized leaf1 (adl1) mutant in rice, which forms abaxially rolled leaves. adl1 leaves are covered with bulliform-like cells, which are normally distributed only on the adaxial surface. An adl1 double mutant with the adaxially snowy leaf mutant, which has albino cells that specifically appear in the abaxial mesophyll tissue, indicated that adl1 leaves show adaxialization in both epidermal and mesophyll tissues. The expression of HD-ZIPIII genes in adl1 mutant increased in mature leaves, but not in the young primordia or the SAM. This indicated that ADL1 may not be directly involved in determining initial leaf polarity, but rather is associated with the maintenance of axis information. ADL1 encodes a plant-specific calpain-like cysteine proteinase orthologous to maize DEFECTIVE KERNEL1. Furthermore, we identified intermediate and strong alleles of the adl1 mutant that generate shootless embryos and globular-arrested embryos with aleurone layer loss, respectively. We propose that ADL1 plays an important role in pattern formation of the leaf and embryo by promoting proper epidermal development.
[D].
URL [本文引用: 1]
[D].
URL [本文引用: 1]
DOI:10.1007/s00438-008-0328-3URLPMID:18293011 [本文引用: 1]
Elucidation of the genetic basis of the control of leaf shape could be of use in the manipulation of crop traits, leading to more stable and increased crop production. To improve our understanding of the process controlling leaf shape, we identified a mutant gene in rice that causes a significant decrease in the width of the leaf blade, termed narrow leaf 7 (nal7). This spontaneous mutation of nal7 occurred during the process of developing advanced back-crossed progeny derived from crosses of rice varieties with wild type leaf phenotype. While the mutation resulted in reduced leaf width, no significant morphological changes at the cellular level in leaves were observed, except in bulli-form cells. The NAL7 locus encodes a flavin-containing monooxygenase, which displays sequence homology with YUCCA. Inspection of a structural model of NAL7 suggests that the mutation results in an inactive enzyme. The IAA content in the nal7 mutant was altered compared with that of wild type. The nal7 mutant overexpressing NAL7 cDNA exhibited overgrowth and abnormal morphology of the root, which was likely to be due to auxin overproduction. These results indicate that NAL7 is involved in auxin biosynthesis.
URLPMID:19304938 [本文引用: 1]
DOI:10.1104/pp.112.199968URL [本文引用: 1]
Leaf rolling is an important agronomic trait in rice (Oryza sativa) breeding and moderate leaf rolling maintains the erectness of leaves and minimizes shadowing between leaves, leading to improved photosynthetic efficiency and grain yields. Although a few rolled-leaf mutants have been identified and some genes controlling leaf rolling have been isolated, the molecular mechanisms of leaf rolling still need to be elucidated. Here we report the isolation and characterization of SEMI-ROLLED LEAF1 (SRL1), a gene involved in the regulation of leaf rolling. Mutants srl1-1 (point mutation) and srl1-2 (transferred DNA insertion) exhibit adaxially rolled leaves due to the increased numbers of bulliform cells at the adaxial cell layers, which could be rescued by complementary expression of SRL1. SRL1 is expressed in various tissues and is expressed at low levels in bulliform cells. SRL1 protein is located at the plasma membrane and predicted to be a putative glycosylphosphatidylinositol-anchored protein. Moreover, analysis of the gene expression profile of cells that will become epidermal cells in wild type but probably bulliform cells in srl1-1 by laser-captured microdissection revealed that the expression of genes encoding vacuolar H+-ATPase (subunits A, B, C, and D) and H+-pyrophosphatase, which are increased during the formation of bulliform cells, were up-regulated in srl1-1. These results provide the transcript profile of rice leaf cells that will become bulliform cells and demonstrate that SRL1 regulates leaf rolling through inhibiting the formation of bulliform cells by negatively regulating the expression of genes encoding vacuolar H+-ATPase subunits and H+-pyrophosphatase, which will help to understand the mechanism regulating leaf rolling.
URLPMID:21765177 [本文引用: 1]
URLPMID:19825577 [本文引用: 1]
DOI:10.1016/j.chom.2013.02.007URL [本文引用: 1]
CERK1 is a lysine motif-containing plant pattern recognition receptor for chitin and peptidoglycan. Chitin recognition by OsCERK1 triggers rapid engagement of a rice MAP kinase cascade, which leads to defense response activation. How the MAP kinase cascades are engaged downstream of OsCERK1 remains obscure. Searching for host proteins that interact with Xoo1488, an effector of the rice pathogen Xanthomonas oryzae, we identified the rice receptor-like cytoplasmic kinase, OsRLCK185. Silencing OsRLCK185 suppressed peptidoglycan-and chitin-induced immune responses, including MAP kinase activation and defense-gene expression. In response to chitin, OsRLCK185 associates with, and is directly phosphorylated by, OsCERK1 at the plasma membrane. Xoo1488 inhibits peptidoglycan-and chitin-induced immunity and pathogen resistance. Additionally, OsCERK1-mediated phosphorylation of OsRLCK185 is suppressed by Xoo1488, resulting in the inhibition of chitin-induced MAP kinase activation. These data support a role for OsRLCK185 as an essential immediate downstream signaling partner of OsCERK1 in mediating chitin-and peptidoglycan-induced plant immunity.
DOI:10.1007/s11105-015-0951-1URL [本文引用: 1]
DOI:10.1111/pce.12696URL [本文引用: 1]
URL [本文引用: 1]
叶绿素是植物叶绿体内参与光合作用的重要色素,叶绿素合成缺陷突变是一类明显的性状突变,其在理论研究和实际应用方面均具有重要的意义.本文介绍了国内外在水稻叶绿素合成缺陷突变体的发掘、作用机理及其基因定位等方面的研究进展.
URL [本文引用: 1]
叶绿素是植物叶绿体内参与光合作用的重要色素,叶绿素合成缺陷突变是一类明显的性状突变,其在理论研究和实际应用方面均具有重要的意义.本文介绍了国内外在水稻叶绿素合成缺陷突变体的发掘、作用机理及其基因定位等方面的研究进展.
DOI:10.1111/j.1365-313X.2007.03384.xURLPMID:18088316 [本文引用: 1]
In plants, Rop/Rac GTPases have emerged as central regulators of diverse signalling pathways in plant growth and pathogen defence. When active, they interact with a wide range of downstream effectors. Using yeast two-hybrid screening we have found three previously uncharacterized receptor-like protein kinases to be Rop GTPase-interacting molecules: a cysteine-rich receptor kinase, named NCRK, and two receptor-like cytosolic kinases from the Arabidopsis RLCK-VIb family, named RBK1 and RBK2. Uniquely for Rho-family small GTPases, plant Rop GTPases were found to interact directly with the protein kinase domains. Rop4 bound NCRK preferentially in the GTP-bound conformation as determined by flow cytometric fluorescence resonance energy transfer measurements in insect cells. The kinase RBK1 did not phosphorylate Rop4 in vitro, suggesting that the protein kinases are targets for Rop signalling. Bimolecular fluorescence complementation assays demonstrated that Rop4 interacted in vivo with NCRK and RBK1 at the plant plasma membrane. In Arabidopsis protoplasts, NCRK was hyperphosphorylated and partially co-localized with the small GTPase RabF2a in endosomes. Gene expression analysis indicated that the single-copy NCRK gene was relatively upregulated in vasculature, especially in developing tracheary elements. The seven Arabidopsis RLCK-VIb genes are ubiquitously expressed in plant development, and highly so in pollen, as in case of RBK2. We show that the developmental context of RBK1 gene expression is predominantly associated with vasculature and is also locally upregulated in leaves exposed to Phytophthora infestans and Botrytis cinerea pathogens. Our data indicate the existence of cross-talk between Rop GTPases and specific receptor-like kinases through direct molecular interaction.
URLPMID:19285078 [本文引用: 1]
DOI:10.3389/fpls.2017.01783URL [本文引用: 1]
URLPMID:16961734 [本文引用: 1]
URLPMID:7920717 [本文引用: 1]
DOI:10.3724/SP.J.1259.2013.00489URL [本文引用: 1]
蓝藻是古老的原核光合生物, 不仅在植物进化上有重要地位, 也是可利用的生物资源及重要的基因资源。Slr1515是来源于集胞藻6803(Synechocystis sp. PCC 6803)的基因, 其同源基因在蓝藻中普遍存在, 但对其生物学功能所知有限。以易于转化的水稻品种中花11(Oryza sativa subsp. japonica cv. ‘Zhonghua 11’)为材料, 通过农杆菌介导的遗传转化方法将Slr1515基因导入水稻胚性愈伤组织中。经过诱导分化, 获得了一系列转基因幼苗。通过PCR、GFP荧光及免疫印迹检测, 筛选出单拷贝插入的转基因阳性株系, 并对野生型和转基因株系的表型进行比较分析。结果表明, 灌浆期转基因株系的剑叶长度比同期的野生型提高了27.7%。进一步的测定结果显示, 转基因株系的剑叶叶绿素含量和PSII最大量子产率分别比野生型提高了0.65倍和4.4%。推测Slr1515基因可能在改善水稻农艺性状方面有一定价值。
DOI:10.3724/SP.J.1259.2013.00489URL [本文引用: 1]
蓝藻是古老的原核光合生物, 不仅在植物进化上有重要地位, 也是可利用的生物资源及重要的基因资源。Slr1515是来源于集胞藻6803(Synechocystis sp. PCC 6803)的基因, 其同源基因在蓝藻中普遍存在, 但对其生物学功能所知有限。以易于转化的水稻品种中花11(Oryza sativa subsp. japonica cv. ‘Zhonghua 11’)为材料, 通过农杆菌介导的遗传转化方法将Slr1515基因导入水稻胚性愈伤组织中。经过诱导分化, 获得了一系列转基因幼苗。通过PCR、GFP荧光及免疫印迹检测, 筛选出单拷贝插入的转基因阳性株系, 并对野生型和转基因株系的表型进行比较分析。结果表明, 灌浆期转基因株系的剑叶长度比同期的野生型提高了27.7%。进一步的测定结果显示, 转基因株系的剑叶叶绿素含量和PSII最大量子产率分别比野生型提高了0.65倍和4.4%。推测Slr1515基因可能在改善水稻农艺性状方面有一定价值。
DOI:10.1105/tpc.107.190180URL [本文引用: 2]
URL [本文引用: 1]
转基因植物中外源基因拷贝数是影响目的基因表达水平和遗传稳定性的重要因素,因此外源基因拷贝数的检测成为转基因研究的关键.利用高通量、快速、灵敏的SYBR Green Ⅰ荧光定量实时PCR法,检测了转大麦烟酰胺合成酶基因(NAS1)水稻中外源基因拷贝数.以蔗糖磷酸合成酶基因(SPS)作为水稻的内源参照基因,通过梯度稀释法,分别获得了NAS1和SPS基因的Ct值与起始模板数的相关性标准曲线,相关系数分别为0.999 76和0.995 71,相关性高.通过目的基因NAS1和水稻内源参照基因SPS起始模板数的比较,获得了目的基因在转基因水稻中的拷贝数,在8株转基因株系中,1株为假阳性,1株拷贝数为1,3株拷贝数为2,其余3株拷贝数分别为3、4和7,而阴性对照拷贝数为0.这种方法快速、简便、准确,可以满足转基因育种工作中对后代优良株系的选择.
URL [本文引用: 1]
转基因植物中外源基因拷贝数是影响目的基因表达水平和遗传稳定性的重要因素,因此外源基因拷贝数的检测成为转基因研究的关键.利用高通量、快速、灵敏的SYBR Green Ⅰ荧光定量实时PCR法,检测了转大麦烟酰胺合成酶基因(NAS1)水稻中外源基因拷贝数.以蔗糖磷酸合成酶基因(SPS)作为水稻的内源参照基因,通过梯度稀释法,分别获得了NAS1和SPS基因的Ct值与起始模板数的相关性标准曲线,相关系数分别为0.999 76和0.995 71,相关性高.通过目的基因NAS1和水稻内源参照基因SPS起始模板数的比较,获得了目的基因在转基因水稻中的拷贝数,在8株转基因株系中,1株为假阳性,1株拷贝数为1,3株拷贝数为2,其余3株拷贝数分别为3、4和7,而阴性对照拷贝数为0.这种方法快速、简便、准确,可以满足转基因育种工作中对后代优良株系的选择.
URL [本文引用: 1]
【目的】明确外源Bt基因插入对水稻倒1叶光合速率及光合作用相关生理特性的影响。【方法】以转Bt基因水稻及亲本水稻为试材,研究盆栽及田间条件下转Bt基因及亲本水稻苗期、拔节期、抽穗期和成熟期倒1叶光合特性及其相关光合酶活性、光合产物积累的动态变化。【结果】转Bt基因及亲本水稻生理活动峰值均出现在拔节期。与亲本水稻相比,转Bt基因水稻苗期叶片净光合速率显著低于亲本;而拔节期和抽穗期,转Bt基因水稻倒1叶净光合速率、叶绿素含量和乙醇酸氧化酶活性显著高于亲本水稻(P<0.05),且拔节期转Bt水稻倒1叶气孔导度和胞间CO2浓度也显著高于亲本水稻(P<0.05);成熟期,转Bt基因与亲本水稻倒1叶光合特性无显著性差异(P>0.05)。【结论】与亲本水稻相比,外源Bt基因的插入对水稻叶片光合特性产生短暂影响,但这种影响不具有持续性。
URL [本文引用: 1]
【目的】明确外源Bt基因插入对水稻倒1叶光合速率及光合作用相关生理特性的影响。【方法】以转Bt基因水稻及亲本水稻为试材,研究盆栽及田间条件下转Bt基因及亲本水稻苗期、拔节期、抽穗期和成熟期倒1叶光合特性及其相关光合酶活性、光合产物积累的动态变化。【结果】转Bt基因及亲本水稻生理活动峰值均出现在拔节期。与亲本水稻相比,转Bt基因水稻苗期叶片净光合速率显著低于亲本;而拔节期和抽穗期,转Bt基因水稻倒1叶净光合速率、叶绿素含量和乙醇酸氧化酶活性显著高于亲本水稻(P<0.05),且拔节期转Bt水稻倒1叶气孔导度和胞间CO2浓度也显著高于亲本水稻(P<0.05);成熟期,转Bt基因与亲本水稻倒1叶光合特性无显著性差异(P>0.05)。【结论】与亲本水稻相比,外源Bt基因的插入对水稻叶片光合特性产生短暂影响,但这种影响不具有持续性。
URL [本文引用: 1]
对12个杂交水稻品种在两类生态条件下的光合特性及干物质积累进行了研究.结果表明:水稻
适应环境的能力主要由其自身因素决定,品种之间存在较大差异.品种间的净光合速率和叶
绿素含量差异均比生态区域间的差异更显著.产量与总干物质量和净光合速率呈显著正相关
,决定系数R2分别为0.584和0.590,与茎鞘物质输出率和茎鞘物质转换率相关不
显著.与低光强地区相比,高光强地区叶片厚度和干物质积累量增加,茎鞘物质输出率和茎
鞘物质转换率降低.在光照充足的条件下,产量中来自于后期光合积累物质的比例较大;在
光照不足的条件下,产量中来自于前期干物质积累及其转运的比例较大.
URL [本文引用: 1]
对12个杂交水稻品种在两类生态条件下的光合特性及干物质积累进行了研究.结果表明:水稻
适应环境的能力主要由其自身因素决定,品种之间存在较大差异.品种间的净光合速率和叶
绿素含量差异均比生态区域间的差异更显著.产量与总干物质量和净光合速率呈显著正相关
,决定系数R2分别为0.584和0.590,与茎鞘物质输出率和茎鞘物质转换率相关不
显著.与低光强地区相比,高光强地区叶片厚度和干物质积累量增加,茎鞘物质输出率和茎
鞘物质转换率降低.在光照充足的条件下,产量中来自于后期光合积累物质的比例较大;在
光照不足的条件下,产量中来自于前期干物质积累及其转运的比例较大.
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
对3个产量水平不同的水稻品种叶绿素含量变化规律进行了研究。结果表明:超高产品种沈农606和高产品种辽粳294在整个生育期内叶绿素含量都高于秋光,特别是在生育后期叶绿素含量下降迟缓,成熟时明显高于秋光,但其叶绿素a/b值始终低于秋光。沈农606和辽粳294齐穗后积累的干物质较多,且产量高于秋光,可能与它们具有较高的叶绿素含量和较低的叶绿素a/b.
URL [本文引用: 1]
对3个产量水平不同的水稻品种叶绿素含量变化规律进行了研究。结果表明:超高产品种沈农606和高产品种辽粳294在整个生育期内叶绿素含量都高于秋光,特别是在生育后期叶绿素含量下降迟缓,成熟时明显高于秋光,但其叶绿素a/b值始终低于秋光。沈农606和辽粳294齐穗后积累的干物质较多,且产量高于秋光,可能与它们具有较高的叶绿素含量和较低的叶绿素a/b.
URLPMID:15490102 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}