 ,1, 王登杰2, 雷仲仁1, 王海鸿,1
,1, 王登杰2, 雷仲仁1, 王海鸿,1Comparison of Life Tables for Experimental Populations of Individual- Rearing and Group-Rearing Frankliniella occidentalis
LI XinHua,1, WANG DengJie2, LEI ZhongRen1, WANG HaiHong,1通讯作者:
责任编辑: 岳梅
收稿日期:2020-05-19接受日期:2020-06-28网络出版日期:2021-03-01
| 基金资助: |
Received:2020-05-19Accepted:2020-06-28Online:2021-03-01
作者简介 About authors
李欣华,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (553KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李欣华, 王登杰, 雷仲仁, 王海鸿. 单头饲养和群体饲养的西花蓟马实验种群生命表比较[J]. 中国农业科学, 2021, 54(5): 959-968 doi:10.3864/j.issn.0578-1752.2021.05.008
LI XinHua, WANG DengJie, LEI ZhongRen, WANG HaiHong.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】西花蓟马(Frankliniella occidentalis)是一种世界性发生的,危害蔬菜、花卉及经济作物的重要害虫[1,2,3],起源于美国西部洛基山脉,2000年在中国云南省首次发现[4],并迅速传播,已造成了巨大的经济损失[4,5,6]。昆虫生命表是研究昆虫种群在一定生态条件下各阶段个体的存活数、死亡数和死亡原因的重要手段[7]。到目前为止,对西花蓟马生命表的研究均采用单头饲养(individual-rearing,IR)方式,包括寄主[8,9,10,11,12]、温度[13,14,15]、杀虫剂[16]、病原物[17]、生殖方式[18]、种群[19]以及复合因素[20,21]的影响。但事实上,在自然条件下西花蓟马常群聚发生[22]。单头饲养和群体饲养(group-rearing,GR)的昆虫生活史之间存在显著差异。因此,根据上述两种饲养方式分别建立生命表,对比生命表数据,可为西花蓟马生命表研究探索更可靠的方法,也为西花蓟马预测预报及防治提供更详实的理论依据。【前人研究进展】单头饲养的雄性太平洋甲虫蟑螂(Diploptera punctata)要比群体饲养(8—10头)的发育历期更长、成虫体型更大[23]。群体饲养(4—35头每组)与单头饲养的西南龟瓢虫(Propylea dissecta)在生长速度、产卵前期、成虫体型等方面有差异[24]。4龄松尺蠖(Bupalus piniarius)幼虫群体饲养时的生长速度快于单头饲养[25]。群体饲养的斑点木蝶(Pararge aegeria)比单头饲养的个体成虫体型小[26]、雌虫产卵量多[27]。因上述饲养方式的不同而产生生活史差异的现象在家蟋蟀(Acheta domesticus)[28]、西方玉米根虫(Diabrotica virgifera)[29]、黏虫(Mythimna separata)[30]、亚洲玉米螟(Ostrinia furnacalis)[31]、二点委夜蛾(Athetis lepigone)[32]、桃小食心虫(Carposina sasakii)[33]、玉米蛀茎夜蛾(Sesamia nonagrioides)[34]、梨豆夜蛾(Anticarsia gemmatalis)[35]、黄粉虫(Tenebrio molitor)[36]、冈比亚按蚊(Anopheles gambiae)[37]、玻里尼西亚斑蚊(Aedes polynesiensis)[38]、稻纵卷叶螟(Cnaphalocrocis medinalis)[39]、小地老虎(Agrotis ipsilon)[40]中也有发现。【本研究切入点】单头饲养西花蓟马的生命表得到了广泛的研究,但目前为止,还没有关于群体饲养西花蓟马的生命表研究。【拟解决的关键问题】通过设置相同饲养环境,比较单头饲养和群体饲养两种方式下的生物学特性和生命表参数,以期为西花蓟马种群动态监测及其综合治理决策提供依据。1 材料与方法
试验于2019—2020年在中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室完成。1.1 供试虫源
西花蓟马2018年采自北京昌平区辣椒(Capsicum annuum)田中。在中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室建立实验种群。饲养于三洋人工气候箱内(MLR-351H,SANYO Electric Co.,Ltd),环境温度(24±1)℃,光周期14 h﹕10 h(L﹕D),相对湿度(70±10)%。以菜豆(Phaseolus vulgaris)为寄主植物和产卵基质。供试全部虫源为10对西花蓟马在菜豆上产卵12 h后取得。1.2 单头饲养和群体饲养西花蓟马各虫态发育历期测定
试验在三洋人工气候箱中进行(气候条件设定同1.1)。将上述含卵寄主转移至双开口圆柱形玻璃罐(底部直径×高=10 cm×30 cm,本试验所用玻璃罐均为同一规格)中,玻璃罐两端用尼龙纱网(200目)封住保持透气,内部放置一张滤纸吸收罐内多余水分。每日在体视显微镜(SZX16,OLYMPUS)下观察卵的孵化情况,将新孵出的1龄若虫转移至新玻璃罐中并记录,作为试验虫源分别进行单头饲养和群体饲养。剩余同批孵化西花蓟马分别以上述两种饲养方式留作备用虫源。每日定时更换新鲜菜豆,观察记录单头饲养和群体饲养西花蓟马羽化前的发育与存活情况。试验虫源配对后,每日定时取出含卵寄主(菜豆)至新玻璃罐中,卵孵化后,将1龄若虫的数量记为产卵量[17]。1.2.1 单头饲养 将新孵出的1龄若虫分装至玻璃罐中,每罐1头,共40罐。待羽化后,在体视显微镜下鉴定成虫性别并记录,然后雌雄配对,若新羽化的试验虫源雌雄数量不对等,则使用备用虫源配对补齐。配对后,继续记录试验虫源的寿命、产卵量,直至其死亡。若试验虫源配偶先死亡,则随机从备用虫源中为其另选配偶进行配对。蛹期取40头备用虫源,测量其蛹重、蛹长、蛹宽,羽化后各取40头雌雄备用虫源分别测量体长、体宽。
1.2.2 群体饲养 所用方法参考文献[41,42]并加以改进,将新挑出的1龄若虫共40头[17]装入玻璃罐中,逐日观察,羽化后鉴定成虫性别,记录其产卵量和寿命。在饲养过程中,若所有雌虫全部死亡,则随机从备用虫源中选取与存活雄虫数量相等的雌性与其进行配对,记录剩余雄性试验虫源的寿命;若所有雄虫全部死亡,则随机从备用虫源中选取与存活雌虫数量相当的雄性进行配对,记录剩余雌性试验虫源的产卵量和寿命。试验虫源全部死亡后试验停止。其蛹重、蛹长、蛹宽,雌雄虫体长、体宽的测量方法同1.2.1。
1.3 数据分析
西花蓟马各阶段发育历期、雌雄虫寿命和雌成虫产卵量使用年龄-阶段两性生命表进行分析[43,44,45]。年龄-阶段特定存活率(age-stage specific survival rate,sxj)由每日记录的存活数计算得到,雌虫年龄-阶段繁殖力(age-stage fecundity,fxj)由每日记录的产卵量计算得到。计算得到以上数据后,根据公式分别算出,特定年龄存活率(age-specific survival rate,$ l_{x}=\sum_{j=1}^{k} s_{x j}$)、特定年龄繁殖力(age-specific fecundity,$ m_{x}=\frac{\sum_{j=1}^{k} s_{x j} f_{x j}}{\sum_{j=1}^{k} s_{x j}}$),年龄期望(life expectancy,$ e_{x j}=\sum_{i=x}^{\infty} \sum_{y=j}^{k} s_{i y}$ [46])。计算得出上述值后,再根据下列公式分别计算,内禀增长率(r,intrinsic rate of increase,$ \sum_{x=0}^{\infty} e^{-r(x+1)} l_{x} m_{x}=1$,初始年龄为0)、周限增长率(λ,finite rate of increase,λ=er)、总生殖率(GRR,gross reproductive rate,GRR=∑mx)、净增殖率(R0,net reproductive rate,$ R_{0}=\sum_{x=0}^{\infty} l_{x} m_{x}$)、平均世代时间(T,mean generation time,T=(ln R0)/r)等参数。其中x代表年龄,j代表阶段,k代表龄期数。s′iy代表一个x年龄j阶段的个体存活到i年龄y阶段的概率,假设sxj=1,计算方法依据CHI等[43]。
根据上述计算公式,原始数据可使用TWOSEX- MSChart[47](
体长、体宽、体重等数据使用SPSS 23.0计算平均值及标准误,采用配对t-test进行显著性分析。
2 结果
2.1 饲养方式对西花蓟马生长发育和繁殖的影响
单头饲养和群体饲养中每阶段各指标如表1所示。其中若虫期、单雌平均产卵量(P<0.001),总产卵前期、蛹重、雌成虫体长、雄成虫体长(P<0.01),蛹期、雄虫寿命(P<0.05)差异显著。单头饲养和群体饲养的卵期、雌成虫期、雄成虫期、雌成虫产卵前期、雌虫寿命、蛹长、蛹宽、雌成虫体宽、雄成虫体宽均无显著差异。单头饲养西花蓟马的若虫期(4.49 d)、蛹期(4.03 d)、雄虫寿命(22.82 d)、总产卵前期(11.37 d)显著长于群体饲养(3.05、3.32、18.64、10.00 d);单头饲养西花蓟马的蛹重(0.03 mg)、雌成虫体长(203.72 μm)、雄成虫体长(149.74 μm)、单雌产卵量(48粒)显著低于群体饲养(0.07 mg、288.81 μm、203.39 μm、133.39粒)。Table 1
Table 1The life history of individual-rearing and group-rearing F. occidentalis
| 指标 Index | 单头饲养 Individual-rearing | 群体饲养 Group-rearing | P | |||
|---|---|---|---|---|---|---|
| n | Mean±SE | n | Mean±SE | |||
| 卵期 Egg stage (d) | 40 | 3.00±0.05 | 40 | 3.03±0.03 | 0.660 | ns |
| 若虫期 Nymph stage (d) | 39 | 4.49±0.13 | 38 | 3.05±0.04 | <0.001 | *** |
| 蛹期 Pupal stage (d) | 34 | 4.03±0.23 | 36 | 3.32±0.15 | 0.021 | * |
| 雌成虫期 Female adult stage (d) | 19 | 20.44±1.42 | 18 | 21.89±1.52 | 0.542 | ns |
| 雄成虫期 Male adult stage (d) | 11 | 11.27±2.52 | 16 | 9.91±0.78 | 0.677 | ns |
| 单雌平均产卵量 Per female oviposition | 19 | 48.00±5.53 | 18 | 133.39±5.42 | <0.001 | *** |
| 雌成虫产卵前期 Adult preoviposition period (d) | 19 | 3.56±0.64 | 18 | 2.75±0.19 | 0.828 | ns |
| 总产卵前期 Total preoviposition period (d) | 30 | 11.37±0.39 | 34 | 10.00±0.005 | 0.003 | ** |
| 雌虫寿命 Female longevity (d) | 19 | 32.06±1.50 | 18 | 28.89±1.52 | 0.200 | ns |
| 雄虫寿命 Male longevity (d) | 11 | 22.82±2.59 | 16 | 18.64±0.88 | 0.047 | * |
| 蛹重 Pupal weight (mg) | 40 | 0.03±0.004 | 40 | 0.07±0.005 | 0.007 | ** |
| 蛹长 Pupal length (μm) | 40 | 170.24±5.17 | 40 | 181.84±4.38 | 0.219 | ns |
| 蛹宽 Pupal width (μm) | 40 | 41.91±3.19 | 40 | 47.32±1.15 | 0.167 | ns |
| 雌成虫体长 Adult length (♀) (μm) | 40 | 203.72±4.13 | 40 | 288.81±14.26 | 0.002 | ** |
| 雄成虫体长 Adult length (♂) (μm) | 40 | 149.74±3.85 | 40 | 203.39±6.77 | 0.002 | ** |
| 雌成虫体宽 Adult width (♀) (μm) | 40 | 42.52±0.81 | 40 | 48.58±2.18 | 0.072 | ns |
| 雄成虫体宽 Adult width (♂) (μm) | 40 | 27.17±2.04 | 40 | 27.35±1.55 | 0.947 | ns |
新窗口打开|下载CSV
2.2 饲养方式对西花蓟马存活率和繁殖率的影响
存活率曲线sxj描述的是个体存活到年龄x阶段j的可能性,可观察到重叠现象[49]。如图1所示,单头饲养若虫总历期(7 vs 4 d)、蛹总历期(8 vs 4 d)长于群体饲养。第8天时,单头饲养若虫已有72.5%化蛹,12.5%的个体仍为若虫,且未出现羽化个体,而群体饲养若虫已全部化蛹,且有20%的已羽化为成虫。第9天时,单头饲养若虫已全部化蛹,但无羽化个体,群体饲养37.5%个体已完成羽化。第10天时,单头饲养有7.5%的个体羽化,此时群体饲养个体已全部羽化。饲养至第14天时,单头饲养个体全部羽化。图1
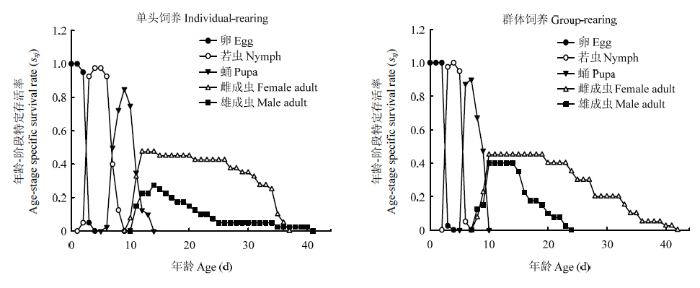 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1单头饲养和群体饲养西花蓟马的特定年龄-阶段特定存活率
Fig. 1Age-stage specific survival rate (sxj) of individual-rearing and group-rearing F. occidentalis
雌虫特定年龄繁殖力(fxj)、特定年龄存活率(lx)、特定年龄繁殖力(mx)、特定年龄繁殖值(lxmx)如图2所示。fxj是年龄x阶段j的单日平均产卵量[49],雌成虫位于生活史的第4阶段,故为fx,4。单头饲养于f14,4出现最大值4.26,群体饲养于f14,4出现最大值为15.56,但f12,4出现次高峰值,与峰值接近,其值为15.44。
图2
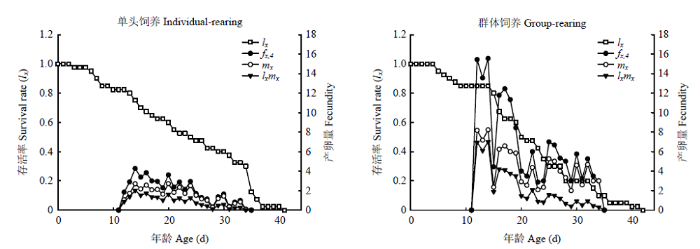 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2单头饲养和群体饲养西花蓟马的特定年龄存活率(lx)、雌虫特定年龄繁殖力(fx,4)、群体特定年龄繁殖力(mx)和特定年龄繁殖值(lxmx)
Fig. 2Age-specific survival rate (lx), female age-specific fecundity (fx,4),age-specific fecundity of total population (mx), and age-specific maternity (lxmx) of individual-rearing and group-rearing F. occidentalis
lx是年龄x的存活率;mx是年龄x时所有个体平均生产子代数;lxmx是lx与mx的乘积,是年龄x时所有存活个体的繁殖值[49]。从图2可以看出单头饲养的 lx值一直匀速下降,第35天,大部分蓟马的寿命达到最大值,曲线开始急剧下降;群体饲养的lx曲线在第16天开始急剧下降,此时试验虫源雄虫个体的数量开始锐减,在饲养至第23天时,试验虫源雄虫全部死亡。
特定年龄生命期望(exj)(图3)是指在年龄x阶段j个体的剩余存活时间,生命期望值会随着年龄增长逐渐降低[49],单头饲养的生命期望(25.7 vs 21.8 d)高于群体饲养。单头饲养雄虫的生命期望从第16天起出现反常并在第24天超过雌虫,原因为其中两头雄虫的寿命超过了平均寿命(22.82 d),分别为24、30 d,与之配对的雌虫均未产卵。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3单头饲养和群体饲养西花蓟马的特定年龄-阶段生命期望
Fig. 3Age-stage life expectancy (exj) of individual-rearing and group-rearing F. occidentalis
特定年龄-阶段繁殖值(vxj)(图4)是指在年龄x阶段j的个体对以后的种群贡献,初孵若虫的繁殖值(v01)等于周限增长率(λ)[49]。单头饲养的v01值为1.175,群体饲养的v01值为1.274。相对于单头饲养(v13,4= 18.36),群体饲养(v12,4=27.70)的繁殖值高峰较高,出现得更早。饲养至23 d时,群体饲养的试验虫源雄虫已全部死亡,使用试验虫源雌虫以雌雄比1﹕1进行配对后,第25天又出现一次峰值(v25,4=20.52)。
图4
新窗口打开|下载原图ZIP|生成PPT图4单头饲养和群体饲养西花蓟马的特定年龄-阶段繁殖值
Fig. 4Age-stage reproductive value (vxj) of individual-rearing and group-rearing F. occidentalis
2.3 饲养方式对西花蓟马后代种群参数的影响
单头饲养的内禀增长率(0.161 d-1)、周限增长率(1.175 d-1)、净增殖率(20.730)和总生殖率(35.699)均极显著低于群体饲养(0.242 d-1、1.274 d-1、60.499、102.342),而平均世代时间(18.70 d)极显著高于群体饲养(16.88 d)(表2)。Table 2
表2
表2单头饲养和群体饲养西花蓟马的种群参数
Table 2
| 种群参数Population parameter | 单头饲养Individual-rearing | 群体饲养Group-rearing | P | |
|---|---|---|---|---|
| 内禀增长率 Intrinsic rate of increase (r) (d-1) | 0.161±0.00004 | 0.242±0.00004 | <0.001 | *** |
| 周限增长率 Finite rate of increase (λ) (d-1) | 1.175±0.00005 | 1.274±0.00005 | <0.001 | *** |
| 净增殖率 Net reproductive rate (R0) | 20.730±0.01 | 60.499±0.03 | <0.001 | *** |
| 平均世代时间 Mean generation time (T) (d) | 18.70±0.003 | 16.88±0.0006 | <0.001 | *** |
| 总生殖率 Gross reproductive rate (GRR) | 35.699±0.02 | 102.342±0.04 | <0.001 | *** |
新窗口打开|下载CSV
3 讨论
单头饲养和群体饲养的西花蓟马在发育历期、成虫体长、蛹重、单雌平均产卵量、成虫寿命等方面存在显著差异。群体饲养的西花蓟马发育历期显著短于单头饲养个体,蛹重和成虫体长显著大于单头饲养个体。此种由于饲养密度不同而引起的发育历期差异的现象在秀丽线虫(Caenorhabditis elegans)上也有发现[50]。群体饲养较单头饲养昆虫发育时间短的现象在太平洋甲虫蟑螂[23]、珠腹珀蟋(Plebeiogryllus guttiventris)[51]、西南龟瓢虫[24]、亚洲玉米螟[31]中也有发现。
与单头饲养相比,群体饲养的昆虫发育历期短、蛹重较重可能与其幼虫期取食成功率较高有关[52,53,54]。例如,觅食中的黑腹果蝇(Drosophila melanogaster)幼虫会通过视觉信号和化学信号(气味等)追踪到正在进食的同种其他个体,减少觅食行为所花费的精力、时间[52]。同样,昆虫的胰岛素信号通路会对其营养摄取量进行反馈,营养摄取量多时该通路相关基因表达量升高,发育时间缩短,体尺增大。反之,发育时间增长,体尺减小[53]。在七叶树蝴蝶(Precis coenia)中,其类固醇激素和蜕皮激素同样可起到调节发育速度和体尺的作用[53]。对于西花蓟马,其发育历期、蛹重、体尺与取食成功率、营养摄取量的关系,以及内在的分子机制,仍有待进一步研究。
群体饲养西花蓟马的单雌平均产卵量显著高于单头饲养。类似的现象在螺旋粉虱(Aleurodicus rugioperculatus)上也有发现[55]。成虫体型和体重影响昆虫繁殖力[56,57,58,59]。埃及伊蚊(Aedes aegypti)在产卵最多时体型大的个体单雌平均产卵量、产卵次数显著高于体型小的个体[56]。斑点木蝶中体型较大的雌性个体体内拥有较多的成熟卵[57]。地中海实蝇(Ceratitis capitata)的雌体体型大小与最大日产卵量显著正相关[58,60]。日本黑蝇(Simulium kawamurae)的体型(翅长、头宽)与繁殖力成正比[59]。黑腹果蝇成虫体重低的个体产卵量少[61]。与单头饲养相比,群体饲养的西花蓟马产卵量较高,可能与其体型较大,繁殖力较强有关。
单头饲养西花蓟马的平均寿命长于群体饲养西花蓟马,类似的现象在秀丽线虫上也有发现[50]。大多数生物(包括昆虫在内)生存能量的分配会在繁殖和寿命间进行权衡,生殖能力强的个体寿命短[62]。当限制黑腹果蝇交配机会(切除雄性的外生殖器)时,雌虫寿命显著增长[63];限制其产卵(移除产卵基质)时,黑腹果蝇的寿命会延长[62,64]。与单头饲养相比,群体饲养的西花蓟马单雌平均产卵量较高、寿命较短,可能与此有关。
单头饲养方式会造成产卵延迟。单头隔离饲养的西花蓟马首次产卵时间比群体饲养平均推迟0.81 d。同样,在秀丽线虫中,单头饲养的群体首次产卵时间显著晚于群体饲养[50]。有报道指出,西花蓟马产卵时间的延迟会造成产卵量的下降[65],可能是因为卵母细胞发育不良或被吸收。单头饲养西花蓟马产卵量低于群体饲养可能与其产卵时间存在延迟有关。
单头饲养西花蓟马的内禀增长率、周限增长率、净增殖率和总生殖率显著小于群体饲养,平均世代时间显著长于群体饲养。由此可见,群体饲养西花蓟马种群扩增要显著优于单头饲养种群。若用单头饲养的方式建立生命表,对西花蓟马种群动态进行预测预报,很可能错过最佳防治时机。
4 结论
单头饲养和群体饲养的西花蓟马生命表参数存在显著差异。群体饲养西花蓟马生命表参数可能更符合田间种群发生动态,建议使用群体饲养的方式建立西花蓟马生命表。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:31536711 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29027089 [本文引用: 2]
URLPMID:22566175 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/jee/tov035URLPMID:26470222 [本文引用: 1]

Frankliniella occidentalis (Pergande) and Frankliniella intonsa (Trybom) are sympatric pests of many greenhouse and field crops in Korea. We compared the influence of constant (27.3 degrees C) and fluctuating temperatures (23.8-31.5 degrees C, with an average of 27.3 degrees C) on the life table characteristics of F. occidentalis and F. intonsa held at a photoperiod of 16:8 (L:D) h and 45+/-5% relative humidity. The development times of both F. occidentalis and F. intonsa were significantly affected by temperature fluctuation, species, and sex. The development time from egg to adult of F. intonsa was shorter than that for F. occidentalis at both constant and fluctuating temperatures. Survival of immature life stages was higher under fluctuating than constant temperature for both thrips species. The total and daily production of first instars was higher in F. intonsa (90.4 and 4.2 at constant temperature, and 95.7 and 3.9 at fluctuating temperatures) than that of F. occidentalis (58.7 and 3.3 at constant temperature, and 60.5 and 3.1 at fluctuating temperatures) under both constant and fluctuating temperatures. The percentage of female offspring was greater in F. intonsa (72.1-75.7%) than in F. occidentalis (57.4-58.7%) under both temperature regimes. The intrinsic rate of natural increase (rm) was higher at constant temperature than at fluctuating temperature for both thrips species. F. intonsa had a higher rm value (0.2146 and 0.2004) than did F. occidentalis (0.1808 and 0.1733), under both constant and fluctuating temperatures, respectively. The biological response of F. occidentalis and F. intonsa to constant and fluctuating temperature was found to be interspecifically different, and F. intonsa may have higher pest potential than F. occidentalis based on the life table parameters we are reporting first here.
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1093/jee/tov091URLPMID:26470219 [本文引用: 3]
We assessed effects of parental exposure to Beauveria bassiana on life history traits of subsequent generations of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Progeny from individuals that survived fungal exposure as second instars had significantly shorter egg stages, but longer prepupal development times than corresponding untreated controls. However, survivorship to adulthood of these progeny groups did not differ. Although fecundities of the parental types did not differ, the sex ratio of progeny from fungal-treated parents was male-biased, whereas sex ratio of progeny from untreated control parents was even. We calculated life table parameters for the progeny and found that all parameters, except for generation time, were significantly less for the progeny of fungal-treated parents than for progeny of untreated parents. The intrinsic rate of increase, finite rate of increase, net reproductive rate, mean generation time, and gross reproductive rate were 0.199 d(-1), 1.229 d(-1), 21.84, 15.48 d, and 27.273, respectively, for progeny of treated thrips, and 0.266 d(-1), 1.316 d(-1), 52.540, 14.92 d, and 70.64, respectively, for progeny of control thrips. Consequently, population projections demonstrated that offspring of parents exposed to B. bassiana would increase their population more slowly than those from untreated parents. These results demonstrate that B. bassiana has sublethal effects that reduce the reproductive success of F. occidentalis and these effects should be taken into account when evaluating its use in management programs for F. occidentalis.
DOI:10.1038/s41598-018-21689-zURLPMID:29463836 [本文引用: 1]
Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) is a serious pest that is capable of bisexual and arrhenotokous reproduction. In arrhenotokous reproduction, virgin females initially produce male offspring; later, when their sons are sexually mature, the mothers begin bisexual reproduction by carrying out oedipal mating with their sons. Because a virgin female produces many male offspring before oedipal mating occurs, multiple oedipal mating is common. In this study, we investigated the effect of multiple oedipal mating on the population growth of F. occidentalis by using the age-stage, two-sex life table theory. In the arrhenotokous cohorts, all unfertilized eggs developed into males. In the bisexual cohorts, the offspring sex ratio was significantly female biased with the mean number of female offspring and male offspring being 72.68 and 29.00, respectively. These were the same as the net reproductive rate of female offspring and male offspring. In arrhenotokous cohorts, the number of males available for oedipal mating significantly affected the production of female offspring. The number of female offspring increased as the number of sons available for oedipal mating increased. Correctly characterizing this unique type of reproduction will provide important information for predicting the timing of future outbreaks of F. occidentalis, as well as aiding in formulating successful management strategies against the species.
URLPMID:20388257 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/ee/20.6.1568URL [本文引用: 1]
DOI:10.1016/0022-1910(83)90040-9URL [本文引用: 2]
[本文引用: 2]
DOI:10.1046/j.1570-7458.2002.00998.xURL [本文引用: 1]
URLPMID:15861232 [本文引用: 1]
DOI:10.1111/eea.2006.118.issue-1URL [本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0212696URLPMID:30822329 [本文引用: 1]
The western corn rootworm, Diabrotica virgifera virgifera (LeConte) (Coleoptera: Chrysomelidae), is an economically important pest of corn in the northern United States. Some populations have developed resistance to management strategies including transgenic corn that produces insecticidal toxins derived from the bacterium Bacillus thuringiensis (Bt). Knowledge of insect dispersal is of critical importance for models of resistance evolution. Larval density affects survival in the field, and stress from crowding often affects facultative long-distance dispersal of adult insects. In this study, we used laboratory flight mills to characterize western corn rootworm flight performance as a function of larval rearing density. Larvae were reared under three densities and the resulting adult females were either allowed to fly voluntarily for 22 h or forced to fly specified durations. For both experiments we also measured lifetime fecundity following flight. The three rearing densities placed differential levels of stress on individuals, as evidenced by decreased survival to adulthood and decreased size of adults at greater rearing density. When larvae were reared under crowded conditions the resulting females were more likely to engage in flight activity, including long uninterrupted flights lasting >10 min, than those reared under low density conditions. Flight and egg production are both energy intensive processes. However, we found no evidence in either voluntary or forced flight experiments of a tradeoff between flight activity and female fecundity. The results suggest that females emerging from high density populations in cornfields are more likely to disperse and disperse farther than those emerging from low density populations. These results are important because they imply that variation in population density in the landscape will affect dispersal, which may in turn require computer models of resistance evolution to incorporate multiple dispersal rates arising from varying larval densities among fields.
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLPMID:30329061 [本文引用: 1]
DOI:10.14411/eje.2008.084URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41598-017-05041-5URLPMID:28684791 [本文引用: 1]
In this study, we first construct an age-stage, two-sex life table for onion maggot, Delia antiqua, grown on three host plants: onion, scallion, and garlic. We found that onion is the optimal host for this species and populations grown on onion have maximum fecundity, longest adult longevity and reproduction period, and the shortest immature developmental time. In contrast, the fecundity on other hosts was lower, particularly on garlic, but these crops can also serve as important secondary hosts for this pest. These data will be useful to the growers to develop specific integrated management programs for each of hosts. We also compared the demographic analyses of using individually-reared and group-reared methods. These two methods provided similar accurate outcomes for estimating insect population dynamics for this species. However, for gregarious species, using the individually-reared method to construct insect life tables produces inaccurate results, and researchers must use group-reared method for life table calculations. When studying large groups of insect, group-reared demographic analysis for age-stage, two-sex life table can also simplify statistical analysis, save considerable labor, and reduce experimental errors.
[本文引用: 2]
DOI:10.1093/jee/tow091URLPMID:27122495 [本文引用: 1]
The oriental fruit fly, Bactrocera dorsalis (Hendel), reduces the quantity and quality of many host fruits through the process of oviposition and larval feeding, and this insect has been considered a major insect pest in several Asian countries for decades. Using an earlier-developed, female-specific system that combines the toxicity of the ricin A chain (RTA) and the alternative RNA splicing property of doublesex ( Bddsx ), we show that transgenic male flies harboring the RTA-Bddsx transgene unevenly repress the pest population through inheritable effects. In age-stage, two-sex life-table analyses, high larval mortality and a delay in pupation were observed after introducing the transgene. The high male to female ratio in DsRed + flies demonstrates the lethal effect of ricin on females. The fitness of both the DsRed + - and DsRed - -transformed females was reduced as shown in the decrease of the net reproductive rate ( R0 ), intrinsic rate ( r ), and finite rate (lambda) values compared with the wild-type populations. The integrity of the RTA-Bddsx transgene remained in more than 80% of DsRed + males after ten generations, supporting the stable inheritance of the transgene. All of the data from this study support the proposed RTA-Bddsx SIT approach, which provides a species-specific and environmentally friendly method of suppressing, rather than eradiating, B. dorsalis.
DOI:10.1093/ee/17.1.26URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 5]
DOI:10.1371/journal.pgen.1006717URLPMID:28394895 [本文引用: 3]
Environmental conditions experienced during animal development are thought to have sustained impact on maturation and adult lifespan. Here we show that in the model organism C. elegans developmental rate and adult lifespan depend on larval population density, and that this effect is mediated by excreted small molecules. By using the time point of first egg laying as a marker for full maturity, we found that wildtype hermaphrodites raised under high density conditions developed significantly faster than animals raised in isolation. Population density-dependent acceleration of development (Pdda) was dramatically enhanced in fatty acid beta-oxidation mutants that are defective in the biosynthesis of ascarosides, small-molecule signals that induce developmental diapause. In contrast, Pdda is abolished by synthetic ascarosides and steroidal ligands of the nuclear hormone receptor DAF-12. We show that neither ascarosides nor any known steroid hormones are required for Pdda and that another chemical signal mediates this phenotype, in part via the nuclear hormone receptor NHR-8. Our results demonstrate that C. elegans development is regulated by a push-pull mechanism, based on two antagonistic chemical signals: chemosensation of ascarosides slows down development, whereas population-density dependent accumulation of a different chemical signal accelerates development. We further show that the effects of high larval population density persist through adulthood, as C. elegans larvae raised at high densities exhibit significantly reduced adult lifespan and respond differently to exogenous chemical signals compared to larvae raised at low densities, independent of density during adulthood. Our results demonstrate how inter-organismal signaling during development regulates reproductive maturation and longevity.
DOI:10.1007/BF01930414URL [本文引用: 1]
DOI:10.1371/journal.pone.0095495URLPMID:24740198 [本文引用: 2]
We quantified the extent and dynamics of social interactions among fruit fly larvae over time. Both a wild-type laboratory population and a recently-caught strain of larvae spontaneously formed social foraging groups. Levels of aggregation initially increased during larval development and then declined with the wandering stage before pupation. We show that larvae aggregated more on hard than soft food, and more at sites where we had previously broken the surface of the food. Groups of larvae initiated burrowing sooner than solitary individuals, indicating that one potential benefit of larval aggregations is an improved ability to dig and burrow into the food substrate. We also show that two closely related species, D. melanogaster and D. simulans, differ in their tendency to aggregate, which may reflect different evolutionary histories. Our protocol for quantifying social behavior in larvae uncovered robust social aggregations in this simple model, which is highly amenable to neurogenetic analyses, and can serve for future research into the mechanisms and evolution of social behavior.
URLPMID:12941617 [本文引用: 3]
DOI:10.1016/j.cois.2017.11.002URL [本文引用: 1]
DOI:10.1093/jee/tov299URLPMID:26494708 [本文引用: 1]
Fecundity is defined as the number of progeny produced by an organism. In our study, we used three different experimental settings to measure the fecundity of rugose spiraling whitefly (Aleurodicus rugioperculatus Martin), a recent invasive insect in Florida, on Strelitzia nicolai host plants. The adult longevity was significantly different among treatments, with females surviving significantly longer in groups in whole-plant cages and individual females in clip cages compared with individual females in whole-plant cages. Females laid 34.5 +/- 8.4, 156.4 +/- 18.3, and 225.2 +/- 18 eggs, on average, when kept individually in whole-plant cages, in a group in whole-plant cages, and individually in clip cages, respectively. In all treatments, there was a significant correlation between the longevity and fecundity of females. Males emerged earlier than females in all treatments. Virgin females produced only males in the F1, while mated females produced both males and females, which suggests an arrhenotokous parthenogenesis. The egg-to-adult survival of the whitefly was 29 +/- 4%, and the highest mortality occurred at the crawler stage when reared on S. nicolai. The results showed a significant effect of aggregation and cage setting on longevity and fecundity of this whitefly. To conclude, the experimental setting of the group fecundity treatment is more similar to what occurs in the landscape. Such a significant effect of cage setting and aggregation on the life-history parameters should be always taken into account when designing experiments or interpreting results.
DOI:10.1111/j.1948-7134.2010.00028.xURLPMID:20618648 [本文引用: 2]
The effects of dietary sugar and body size on the oviposition of Ae. aegypti were studied under laboratory conditions. In female mosquitoes provided with sugar, the start of maximum fecundity was significantly delayed and the oviposition period was longer than in females provided with water. The peak of oviposition was also delayed in sugar-fed females. Large females oviposited more eggs per day than small females at maximum fecundity and during eight days of observations. Large females also visited significantly more water-containing cups in their cages per day than small females at maximum fecundity. During the eight days of observations, large females and sugar-fed females visited more water-containing cups in their cages than water-fed small females. Both large females and sugar-fed females oviposited their eggs at sites higher above the water line than water-fed small females. These results suggested that large and sugar-fed female Ae. aegypti mosquitoes had more energy reserves and oviposited their eggs at higher sites, which would lead to a time lag in hatching.
DOI:10.1111/fec.2008.22.issue-3URL [本文引用: 2]
DOI:10.1093/aesa/92.2.278URL [本文引用: 2]
URLPMID:1495068 [本文引用: 2]
URLPMID:10747041 [本文引用: 1]
DOI:10.1016/j.exger.2006.10.009URL [本文引用: 1]
Abstract
The extension of life span by diet restriction in Drosophila has been argued to occur without limiting calories. Here we directly measure the calories assimilated by flies when maintained on full- and restricted-diets. We find that caloric intake is reduced on all diets that extend life span. Flies on low-yeast diet are long-lived and consume about half the calories of flies on high-yeast diets, regardless of the energetic content of the diet itself. Since caloric intake correlates with yeast concentration and thus with the intake of every metabolite in this dietary component, it is premature to conclude for Drosophila that calories do not explain extension of life span.DOI:10.1016/j.exger.2010.10.008URL [本文引用: 2]
Reproduction shortens lifespan in practically all organisms examined so far, but the underlying mechanisms remain largely unknown to date. Here I review what evolutionary and molecular biologists have learned about such "costs of reproduction" in the fruit fly (Drosophila melanogaster) since Maynard Smith's (1958) seminal discovery that sterile mutants in D. subobscura live substantially longer than fertile wildtype flies. Together with observations from the nematode worm (Caenorhabditis elegans) and other organisms, the data from Drosophila suggest that there are at least four general principles that underlie trade-offs between reproduction anti lifespan: (1) trade-offs between survival and reproduction are widespread; (2) the relationship between increased lifespan and decreased fecundity can be uncoupled under certain conditions; (3) while survival costs of reproduction might not necessarily be due to competitive resource allocation, we lack robust alternative explanations for their occurrence; and (4) physiological trade-offs between reproduction and longevity do not always translate into evolutionary genetic trade-offs. I conclude that despite much recent progress - our current understanding of the proximate basis of survival costs of reproduction remains very limited; much future work on the genetics and physiology of such trade-offs will be required to uncover their mechanistic basis. (C) 2010 Elsevier Inc.
DOI:10.1038/338760a0URL [本文引用: 1]
DOI:10.1016/0022-1910(87)90060-6URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}