 ,, 马红叶, 李荣飞, 杨仕品, 乔荣, 钟霈霖,贵州省农业科学院园艺研究所,贵阳 550006
,, 马红叶, 李荣飞, 杨仕品, 乔荣, 钟霈霖,贵州省农业科学院园艺研究所,贵阳 550006Metabolic Analysis of Aroma Components in Two Interspecific Hybrids from the Cross of F.ananassa Duch. and Fragaria nilgerrensis Schlecht.
WANG AiHua,, MA HongYe, LI RongFei, YANG ShiPin, QIAO Rong, ZHONG PeiLin,Institute of Horticulture, Guizhou Academy of Agricultural Sciences, Guiyang 550006通讯作者:
责任编辑: 赵伶俐
收稿日期:2020-05-27接受日期:2020-10-21网络出版日期:2021-03-01
| 基金资助: |
Received:2020-05-27Accepted:2020-10-21Online:2021-03-01
作者简介 About authors
王爱华,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1106KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王爱华, 马红叶, 李荣飞, 杨仕品, 乔荣, 钟霈霖. 凤梨草莓与黄毛草莓种间杂种果实香气成分的代谢谱分析[J]. 中国农业科学, 2021, 54(5): 1043-1054 doi:10.3864/j.issn.0578-1752.2021.05.015
WANG AiHua, MA HongYe, LI RongFei, YANG ShiPin, QIAO Rong, ZHONG PeiLin.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】香气作为草莓最重要的品质特征之一,备受消费者关注。近几十年来,育种家以提高农艺性能为目标,选育出了各种果实大、色泽艳丽、产量高的草莓新品种[1]。但是这些草莓新品种多为八倍体凤梨草莓(8×)的后代,遗传背景狭窄,香型单一,香气淡;野生草莓果实虽小,但蕴藏的香型丰富,香气浓,为了追求草莓香型的多样性,野生草莓受到草莓育种家的青睐,被认为是新型香味分子的重要供体[2]。因此,开展凤梨草莓与野生草莓种间杂种的香气研究对育种实践具有重要意义。【前人研究进展】前人在草莓果实风味方面进行了较多研究,如王玲等[3]分析了‘达赛莱克特’草莓果实挥发性物质的组成与含量特征,揭示了果实发育成熟过程中香气动态变化规律;DONG等[4]比较了凤梨草莓与森林草莓酯类的组成差异。目前,国内外有关草莓香气的研究主要集中在凤梨草莓和模式种森林草莓果实的香气成分鉴定、比较分析及果实发育过程中香气动态变化规律方面,而仅有2篇关于黄毛草莓香气成分的报道,STAUDT等[5]在黄毛草莓中检测到120种挥发性物质;ZHAO等[6]从3份黄毛草莓试材中鉴定出112种香气成分;有关黄毛草莓果实蜜桃香气特征成分的鉴定尚缺乏系统报道。常用的香气提取方法有溶剂辅助风味蒸发法(solvent-assisted flavor evaporation,SAFE)、液-液萃取法(liquid-liquid extraction,LLE)、超临界流体萃取法(supercritical fluid extraction method,SFE)和顶空固相微萃取法(headspace solid-phase microextraction,HS-SPME)等[7,8]。与传统的萃取方法相比,顶空固相微萃取无需或仅需很少的有机溶剂,无需复杂的样品预处理,快捷简便,可直接在GC-MS上分析。目前,顶空固相微萃取和气相色谱-质谱联用技术已成为国际公认的植物果实及食品香气提取方法[8,9],并广泛应用于草莓果实的香气成分分析鉴定中,如张运涛等[10]以日光温室栽培的‘甜查理’和‘章姬’草莓为试材,采用该技术测定了草莓果实发育过程中挥发物种类和质量分数的动态变化;曾祥国等[11]用该技术研究了‘晶玉’‘甜查理’‘晶瑶’‘章姬’和‘丰香’5个草莓品种果实挥发性物质成分的差异。香气成分测定技术的发展加速了草莓果实挥发性物质的鉴定,草莓属已有360多种挥发性物质被鉴定出来,这些物质包括酯类、醛类、酮类、醇类、萜烯类和呋喃酮类等[12,13,14],虽然个别种类通常以痕量出现,但可能对草莓的整体香气产生重大影响[2,15]。研究认为丁酸甲酯、丁酸乙酯、己酸甲酯、己酸乙酯等是草莓果味及青草味的来源[7],萜类是草莓花香味的来源[1,16],呋喃类赋予草莓典型的焦糖味[1],内酯类通常跟桃香味有关[17]。黄毛草莓(Fragaria nilgerrensis)为亚洲东部和东南部地区所特有的一种野生二倍体草莓[18],其种质中蕴藏着浓郁的蜜桃香气,是香气育种的理想材料,马鸿翔等[19]对黄毛草莓与凤梨草莓种间杂种进行了细胞遗传学分析;日本****NOGUCHI等[20,21]对中国云南的黄毛草莓与凤梨草莓开展种间杂交,育成了具有蜜桃香味的商业品种——‘桃熏’和‘久留米IH1号’等。【本研究切入点】尽管****们开展了凤梨草莓与黄毛草莓的种间杂交利用,但有关其种间杂种果实香气成分的代谢谱分析却鲜有报道。【拟解决的关键问题】本研究以亲本相同、遗传背景较一致,但蜜桃香气差异明显的凤梨草莓与黄毛草莓的种间杂种果实为试材,采用顶空固相微萃取和GC-MS技术,结合多元统计分析方法,分析其香气成分的代谢谱,为黄毛草莓蜜桃香气特征成分鉴定及利用野生草莓资源进行香气育种提供科学参考。1 材料与方法
1.1 供试材料
以凤梨草莓栽培品种‘红颜’(Benihoppe,2n= 8x)为母本,野生黄毛草莓(Fragaria nilgrrensis,2x)为父本杂交获得五倍体远缘杂交后代,五倍体匍匐茎茎尖经过染色体加倍后获得的十倍体成苗(母本)再与凤梨草莓栽培品种‘小白’(父本)杂交获得2个在蜜桃香气上存在明显差异的种间杂种PF(具黄毛草莓浓郁的蜜桃香气)和NF(无蜜桃香气)。PF和NF均种植于贵州省园艺研究所育种棚,于2018年2月18日采摘PF和NF完熟期各6个果实,进行果实性状比较及香气成分的代谢谱分析。1.2 果实性状比较
2个种间杂种PF和NF果实性状比较参考赵密珍[22]《草莓种质资源描述规范和数据标准》进行,平均单果重采用一级序果实平均值,果实硬度采用杭州托普仪器有限公司生产的GY-2型号水果硬度计,探头直径3.8 mm。1.3 香气成分的代谢谱分析
采用顶空固相微萃取法进行代谢物提取,参考贺书珍等[23]的方法进行,略有改动。进样前先将萃取头在气相色谱进样口老化1 h,老化温度为250℃,迅速取20 mg果肉匀浆于20 mL钳口样品瓶中,将老化好的萃取头插入样品瓶中顶空部位,40℃下恒温搅拌,平衡10 min后,萃取30 min,然后将萃取头抽出,插入安捷伦7890B GC/5977B MS联用仪(Agilent,USA),于250℃解析2 min,进行GC-MS分析。具体分析条件如下:萃取温度60℃;预热时间15 min;萃取时间30 min;解析时间4 min;分流模式Splitless Mode;隔垫吹扫流速3 mL?min-1;载气为Helium;色谱柱:DB-Wax(30 m×250 μm×0.25 μm);柱流速1 mL?min-1;柱箱升温程序:40℃保持4 min,以5℃?min-1升至245℃,保持5 min;前进样口温度 250℃;传输线温度250℃;离子源温度230℃;四极杆温度150℃;电离电压-70 eV;质量范围为20—500 aum;扫描模式Scan;溶剂延迟0 min。1.4 数据分析
使用ChromaTOF软件和NIST library(2 结果
2.1 2个种间杂种果实性状比较
凤梨草莓和黄毛草莓种间杂种PF和NF果实(图1)除蜜桃香气具有明显差异外,其他性状较一致,平均单果重分别为19.26和20.74 g,果面红色,果肉白色,果实硬度分别为0.96和1.03 kg?cm-2,髓心空洞小,果肉质地绵软,风味酸甜适中(表1)。Table 1
表1
表12个种间杂种PF和NF果实性状比较
Table 1
| 株系 Lines | 蜜桃香气 Honey peach aroma | 平均单果重 Mean fruit weight (g) | 果面颜色 Color of fruit | 果肉颜色 Flesh color | 果实硬度 Fruit firmness (kg?cm-2) | 髓心空洞 Fruit cavity size | 果肉质地 Fruit texture | 风味 Flavor |
|---|---|---|---|---|---|---|---|---|
| PF | 浓 Much | 19.26±0.81 | 红 Red | 白 White | 0.96±0.08 | 小 Small | 绵 Mealy | 酸甜适中 Sweet-Sour |
| NF | 无 None | 20.74±1.02 | 红 Red | 白 White | 1.03±0.19 | 小 Small | 绵 Mealy | 酸甜适中 Sweet-Sour |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12个种间杂种果实PF(左)和NF(右)
Fig. 1Fruits of two interspecific hybrids PF (Left) and NF (Right)
2.2 多元统计分析
总样本的PCA分析见图2,NF组主要分布在PC1左侧,PF组主要分布在PC1右侧,未见离散样本,说明在95%的置信区间NF组和PF组组间差异较大,组内差异较小,证实该数据质量好,分析模型稳健可靠。为了找到组间的差异代谢物质,在PCA模型基础上,建立了偏最小二乘法判别分析模型得分散点图(图3),并采用200次响应排序的方法对模型进行排序检验(图4),R2=0.74,Q2=-1.19,R2数据大于Q2数据,且Q2回归线与Y轴截距小于0,表明模型未“过拟合”,可根据VIP值分析筛选差异代谢物。图2
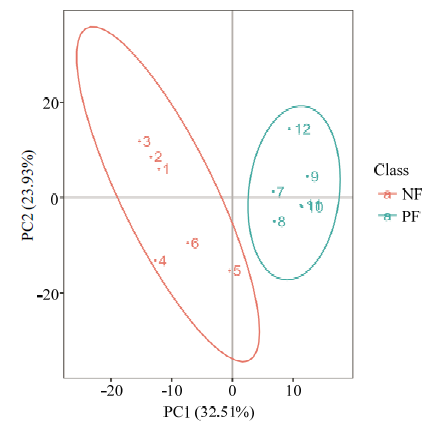 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2总样品的PCA得分图
Fig. 2PCA score scatter plots of total samples
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3比较组的PLS-DA得分图
R2Y:PLS-DA模型的解释率,Q2Y:PLS-DA模型的预测能力
Fig. 3PLS-DA score scatter plots of comparison group
R2Y: The interpretation rate of the PLS-DA model; Q2Y: The predictive ability of the PLS-DA model
图4
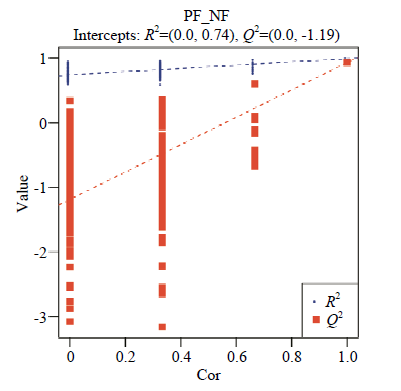 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4使用200次响应法对PLS-DA进行验证的点图
Cor:随机分组的Y与原始分组Y的相关性;Value:R2和Q2的得分
Fig. 4Validation plots of PLS-DA with 200 permutation
Cor: The correlation between the randomly grouped Y and the original group Y; Value: The score of R2 and Q2
2.3 总代谢物分析
2.3.1 种类分析 在供试的2组样品中,共检测到383种香气物质(图5),包括141种酯、41种醇、40种酮、36种烷烃、22种醛、17种烯烃、17种酸、13种内酯、10种环烷烃、7种醚、6种呋喃、5种苯酚、5种胺、3种糖和3种苯,其中酯类占总代谢物的36.81%,醇、酮和烷烃分别占10.70%、10.44%和9.40%。图5
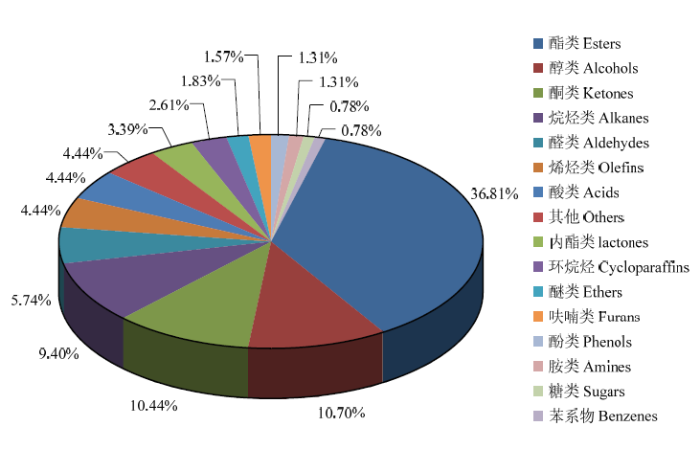 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5总代谢物种类分析
Fig. 5Analysis on category of the total metabolites
2.3.2 功能及分类注释 383种总代谢物中注释到KEGG pathyway的有16种(图6),占总代谢物的4.18%,通过Pathway分析16种代谢物参与的代谢通路有7条,包括氨基酸代谢、碳水化合物代谢、能量代谢、全局和概述地图、酯类代谢、萜类和聚酮类代谢及其他氨基酸代谢通路。lipid maps数据库分类注释的代谢物有31种(图7),占总代谢物的8.09%,包括含氧烃类化合物、烃类、脂肪醛、脂肪醇、脂肪酸和共轭物共5类。
图6
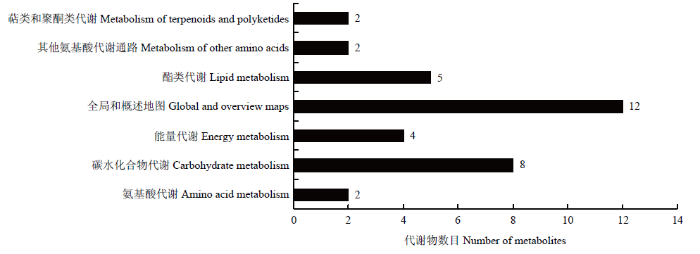 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6KEGG通路注释
Fig. 6KEGG pathway annotation
图7
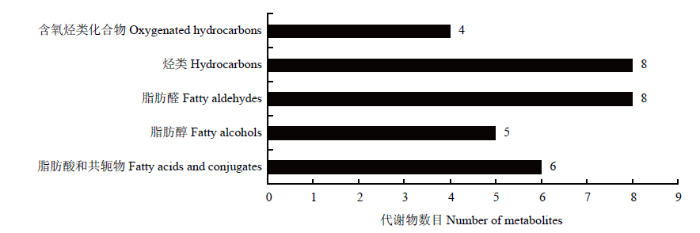 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7Lipid maps注释
Fig. 7Lipid maps annotation
2.4 差异代谢物分析
2.4.1 种类分析 从383种总代谢物中共筛选出67种差异代谢物,包括32种酯、7种内酯、6种醇、4种烯烃、4种酮、4种醛、2种酸,烷烃、糖、醚、环烷烃、呋喃、苯酚、苯和胺各1种,其中酯类占总差异代谢物的47.76%,内酯类和醇类分别占10.45%和8.96%(图8)。差异代谢物中,注释到KEGG数据库的有2种,为二甲硫醚和辛酸,分别在能量代谢和Global and overview maps代谢通路上。注释到LIPID数据库的有4种,为辛酸、辛醇、庚醛和5-庚烯-2-酮,分别属于脂肪酸和共轭物、脂肪醇、脂肪醛和含氧烃类化合物。图8
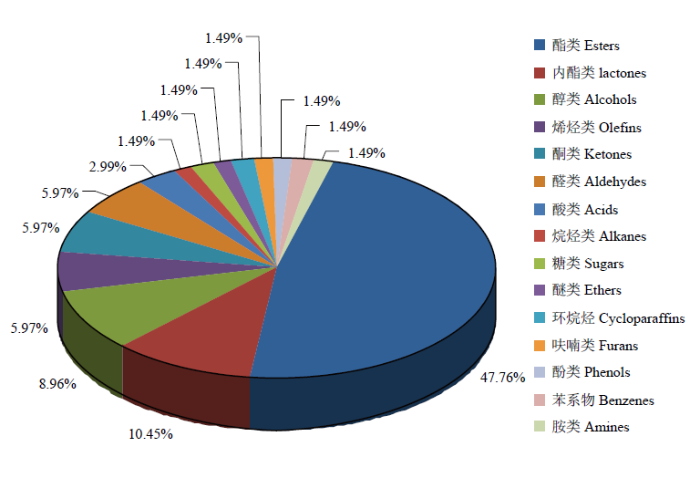 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8差异代谢物种类分析
Fig. 8Analysis on category of the significantly different metabolites
2.4.2 log2FC分析 PF相对于NF,67种差异代谢物中有58种上调,9种下调(表2)。上调幅度较大的差异代谢物为内酯类物质,log2FC排在前3名的依次是(Z)-7-癸烯-5-酸(5.60)、丁位十一内酯(5.33)、δ-癸内酯(5.30);下调幅度较大的差异代谢物为酯类物质,-log2FC排在前3名的依次是肉桂酸乙酯、亚硫酸(-7.19),2-乙基己基异己酯(-6.65)和3-羟基丁酸乙酯(-4.14)。
Table 2
表2
表2基于GC-MS的PF和NF果实中差异代谢物分析
Table 2
| 序号 No. | 化合物名称 Compound name | 保留时间 Retention time (min) | CAS号 CAS # | 相对含量 Relative content (%) | log2FC | P-value | VIP | |
|---|---|---|---|---|---|---|---|---|
| PF | NF | |||||||
| 酯类 Esters | ||||||||
| 1 | 3,7,11-三甲基-1,6,10-十二烷三烯-3-醇乙酸酯 Nerolidyl acetate | 31.46 | 2306-78-7 | 0.11 | 0.08 | 3.63 | 1.52E-02 | 1.54 |
| 2 | 己酸癸酯 Decyl hexanoate | 30.63 | 52363-43-6 | 2.16 | 0.47 | 3.57 | 7.62 E-03 | 2.84 |
| 3 | 异戊酸癸酯 Isovaleric acid, decyl ester | 27.27 | 72928-48-4 | 1.16 | 0.26 | 3.54 | 1.17 E-03 | 2.52 |
| 4 | 乙酸月桂酯 Lauryl acetate | 27.99 | 112-66-3 | 0.13 | 0.11 | 3.21 | 7.28 E-03 | 1.54 |
| 5 | 乙酸癸酯 Decyl acetate | 23.52 | 112-17-4 | 2.21 | 0.59 | 3.18 | 1.94E-03 | 2.63 |
| 6 | 1-Octen-1-ol, acetate | 20.06 | 2761-31-1 | 0.03 | 0.03 | 3.11 | 0.03 | 1.31 |
| 7 | 癸基辛酸酯 Decyl octanoate | 34.34 | 2306-89-0 | 0.01 | 0.01 | 3.01 | 0.02 | 1.39 |
| 8 | 环丙烷羧酸癸酯 Cyclopropanecarboxylic acid,decyl ester | 21.09 | - | 0.19 | 0.13 | 2.91 | 0.03 | 1.35 |
| 9 | 4-Hexen-1-ol, acetate | 14.24 | 72237-36-6 | 0.21 | 0.16 | 2.86 | 2.01E-03 | 1.55 |
| 10 | (Z)-7-十二碳烯-1-醇乙酸酯 cis-7-Dodecen-1-yl acetate | 28.62 | 14959-86-5 | 0.06 | 0.07 | 2.74 | 0.02 | 1.21 |
| 11 | (Z)-Dec-4-enyl isobutyl carbonate | 34.71 | - | 0.43 | 0.22 | 2.59 | 1.94E-03 | 1.83 |
| 12 | (Z)-Dec-4-en-1-yl 2-methylbutanoate | 27.61 | - | 2.35 | 1.22 | 2.57 | 3.45E-03 | 1.80 |
| 13 | 2-十二烯基乙酸酯trans-2-Dodecen-1-ol, acetate | 23.98 | 38363-23-4 | 9.59 | 2.90 | 2.49 | 9.12E-03 | 2.73 |
| 14 | 2-Butenoic acid, 2-methyl-, 2-methylpropyl ester | 26.78 | 66917-61-1 | 0.06 | 0.08 | 2.29 | 0.01 | 1.15 |
| 15 | Cyclobutanecarboxylic acid, 4-methylpentyl ester | 3.27 | - | 0.03 | 0.04 | 2.27 | 4.08E-04 | 1.30 |
| 序号 No. | 化合物名称 Compound name | 保留时间 Retention time (min) | CAS号 CAS # | 相对含量 Relative content (%) | log2FC | P-value | VIP | |
| PF | NF | |||||||
| 16 | (Z)-己-2-烯基乙酸酯 2-Hexen-1-ol, acetate, (Z)- | 14.73 | 56922-75-9 | 0.41 | 0.55 | 2.21 | 7.21 E-04 | 1.39 |
| 17 | 乙酸正壬酯 Acetic acid, nonyl ester | 21.13 | 143-13-5 | 0.05 | 0.08 | 2.16 | 0.01 | 1.05 |
| 18 | 3-乙酰氧基-3-羟基-2-甲基丙酸甲酯3-Acetoxy-3-hydroxy-2-methylpropionic acid, methyl ester | 25.99 | - | 0.42 | 0.60 | 2.05 | 5.61E-06 | 1.24 |
| 19 | 癸酸甲酯 Decanoic acid, methyl ester | 21.52 | 110-42-9 | 0.47 | 0.78 | 2.02 | 4.09E-03 | 1.06 |
| 20 | 2-四氢糠酸甲酯Methyl2-tetrahydrofuroate | 34.62 | 37443-42-8 | 0.08 | 0.12 | 2.01 | 1.70 E-03 | 1.11 |
| 21 | 乙酸丁香酚酯 Acetyl eugenol | 33.38 | 93-28-7 | 0.04 | 0.06 | 1.98 | 2.48 E-04 | 1.15 |
| 22 | 1,4-丁二醇二乙酸酯 1,4-Butanediol, diacetate | 25.64 | 628-67-1 | 0.57 | 0.28 | 1.96 | 0.02 | 2.00 |
| 23 | Fumaric acid, di(dec-4-enyl) ester | 30.99 | - | 7.07 | 4.77 | 1.87 | 0.04 | 2.39 |
| 24 | 醋酸辛酯 Acetic acid, octyl ester | 18.56 | 112-14-1 | 8.75 | 5.61 | 1.39 | 0.04 | 2.29 |
| 25 | (S)-3-羟基丁酸甲酯 Butanoic acid, 3-hydroxy-, methyl ester, (S)- | 18.65 | 53562-86-0 | 0.48 | 6.79 | -2.15 | 0.03 | 1.09 |
| 26 | 氨茴酸甲酯 Methyl anthranilate | 34.65 | 134-20-3 | 3.18E-03 | 0.06 | -2.24 | 0.01 | 1.13 |
| 27 | 2-甲氧基苯甲酸甲酯 Benzoic acid, 2-methoxy-, methyl ester | 31.53 | 606-45-1 | 0.02 | 0.46 | -2.54 | 6.47E-03 | 1.27 |
| 28 | 亚硫酸,环己基甲基十二烷基酯 Sulfurous acid, cyclohexylmethyl dodecyl ester | 30.71 | - | 0.04 | 0.57 | -2.79 | 0.04 | 1.18 |
| 29 | 丁酸乙酯 Butanoic acid, ethyl ester | 6.07 | 105-54-4 | 0.47 | 17.92 | -3.33 | 0.03 | 2.28 |
| 30 | 3-羟基丁酸乙酯 Butanoic acid, 3-hydroxy-, ethyl ester | 19.61 | 5405-41-4 | 0.08 | 9.93 | -4.14 | 0.02 | 2.53 |
| 31 | 亚硫酸,2-乙基己基异己酯 Sulfurous acid, 2-ethylhexyl isohexyl ester | 25.35 | - | 2.87E-03 | 0.72 | -6.65 | 0.03 | 2.75 |
| 32 | 肉桂酸乙酯 2-Propenoic acid, 3-phenyl-, ethyl ester | 32.72 | 103-36-6 | 7.23E-03 | 1.53 | -7.19 | 0.03 | 2.48 |
| 合计 Sum | 37.69 | 57.20 | ||||||
| 内酯类 Lactones | ||||||||
| 33 | (Z)-7-癸烯-5-酸 2H-Pyran-2-one, tetrahydro-6-(2-pentenyl)-, (Z)- | 35.01 | 25524-95-2 | 2.38 | 0.17 | 5.60 | 2.38E-05 | 3.55 |
| 34 | 丁位十一内酯 2H-Pyran-2-one, 6-hexyltetrahydro- | 37.94 | 710-04-3 | 0.55 | 0.08 | 5.33 | 2.61E-05 | 2.97 |
| 35 | δ-癸内酯2H-Pyran-2-one, tetrahydro-6-pentyl- | 33.89 | 705-86-2 | 12.53 | 1.01 | 5.30 | 1.33 E-04 | 3.44 |
| 36 | 2H-Pyran-2-one, 5,6-dihydro-6-propyl- | 34.59 | 16400-69-4 | 0.05 | 0.07 | 2.53 | 2.95 E-03 | 1.33 |
| 37 | 丁位己内酯2H-Pyran-2-one,tetrahydro-6-methyl- | 25.83 | 823-22-3 | 2.74 | 1.71 | 2.30 | 3.35 E-03 | 1.61 |
| 38 | 丁位辛内酯 2H-Pyran-2-one, tetrahydro-6-propyl- | 29.55 | 698-76-0 | 2.08 | 2.45 | 1.52 | 0.02 | 1.10 |
| 39 | 丙位壬内酯 2(3H)-Furanone, dihydro-5-pentyl- | 30.75 | 104-61-0 | 0.58 | 0.63 | 1.31 | 0.04 | 1.20 |
| 合计 Sum | 20.91 | 6.12 | ||||||
| 酮类 Ketones | ||||||||
| 40 | β-紫罗兰酮 3-Buten-2-one, 4-(2,6,6-trimethyl-1-cyclohexen-1-yl)- | 29.03 | 79-77-6 | 0.08 | 0.11 | 2.04 | 8.57 E-04 | 1.14 |
| 41 | 2-庚酮 2-Heptanone | 10.25 | 110-43-0 | 14.09 | 7.74 | 1.94 | 0.02 | 2.47 |
| 42 | 5-庚烯-2-酮 5-Hepten-2-one | 12.32 | 6714-00-7 | 0.64 | 0.84 | 1.93 | 1.67E-04 | 1.13 |
| 序号 No. | 化合物名称 Compound name | 保留时间 Retention time (min) | CAS号 CAS # | 相对含量 Relative content (%) | log2FC | P-value | VIP | |
| PF | NF | |||||||
| 43 | 3-壬烯-2-酮 3-Nonen-2-one | 19.41 | 14309-57-0 | 0.49 | 0.43 | 1.62 | 0.02 | 1.77 |
| 合计 Sum | 15.30 | 9.12 | ||||||
| 醇类 Alcohols | ||||||||
| 44 | 1-癸醇 1-Decanol | 25.45 | 112-30-1 | 0.89 | 0.25 | 4.50 | 2.23E-05 | 2.60 |
| 45 | 3-环戊基-1-丙醇3-Cyclopentyl-1-propanol | 34.09 | 767-05-5 | 0.04 | 0.04 | 3.07 | 0.02 | 1.24 |
| 46 | 反式-3-己烯-1-醇 3-Hexen-1-ol | 16.19 | 544-12-7 | 0.31 | 0.38 | 2.82 | 1.04E-03 | 2.14 |
| 47 | (Z)-4-癸烯-1-醇 (Z)-4-Decen-1-ol | 26.16 | 57074-37-0 | 1.74 | 0.66 | 2.39 | 8.64E-03 | 2.18 |
| 48 | 苯甲醇 Benzyl alcohol | 27.74 | 100-51-6 | 0.97 | 1.32 | 1.66 | 0.01 | 1.06 |
| 49 | 辛醇 1-Octanol | 20.71 | 111-87-5 | 5.91 | 4.86 | 1.52 | 0.02 | 1.37 |
| 合计 Sum | 9.86 | 7.51 | ||||||
| 醛类 Aldehydes | ||||||||
| 50 | 苯乙醛 Benzeneacetaldehyde | 22.43 | 122-78-1 | 6.74 | 9.22 | 2.20 | 9.46E-04 | 1.45 |
| 51 | 2-甲基丁醛 Butanal, 2-methyl- | 3.36 | 96-17-3 | 0.21 | 0.42 | 1.77 | 0.03 | 1.06 |
| 52 | 庚醛 Heptanal | 10.31 | 111-71-7 | 0.93 | 1.50 | 1.73 | 3.39E-04 | 1.05 |
| 53 | 糠醛 Furfural | 18.08 | 98-01-1 | 0.44 | 0.63 | 1.58 | 0.01 | 1.01 |
| 合计 Sum | 8.32 | 11.77 | ||||||
| 酸类 Acids | ||||||||
| 54 | 辛酸 Octanoic acid | 31.90 | 124-07-2 | 5.24 | 4.98 | 2.47 | 7.39E-03 | 1.37 |
| 55 | 异辛酸 Hexanoic acid, 2-ethyl- | 30.08 | 149-57-5 | 0.31 | 0.46 | 2.02 | 5.31E-03 | 1.09 |
| 合计 Sum | 5.55 | 5.44 | ||||||
| 烯烃类 Olefins | ||||||||
| 56 | 5-亚乙基-1-甲基-环庚烯 Cycloheptene, 5-ethylidene-1-methyl- | 25.41 | 15402-94-5 | 0.54 | 0.31 | 3.51 | 2.00E-04 | 1.97 |
| 57 | 4, 6(Z), 8(E)-大柱三烯 Megastigma-4, 6(Z), 8(E)-triene | 21.15 | 55497-53-5 | 0.13 | 0.16 | 2.05 | 5.10 E-04 | 1.17 |
| 58 | 7(E),9,13-大柱三烯 egastigma-7(E),9,13-triene | 17.82 | - | 0.31 | 0.40 | 1.98 | 9.91E-04 | 1.12 |
| 59 | Z-5-十九碳烯 Z-5-Nonadecene | 24.91 | - | 0.08 | 0.11 | 1.92 | 0.02 | 1.32 |
| 合计 Sum | 1.06 | 0.98 | ||||||
| 其他 Others | ||||||||
| 60 | N-甲基-叔丁基胺N-tert-Butylmethylamine | 2.36 | 14610-37-8 | 0.05 | 0.03 | 3.01 | 3.47E-05 | 1.76 |
| 61 | 1,5-二甲基-6-氧杂双环[3.1.0]己烷 1,5-Dimethyl-6-oxa-bicyclo[3.1.0]hexane | 4.90 | 82461-31-2 | 0.10 | 0.07 | 2.97 | 1.25E-05 | 1.75 |
| 62 | 4-烯丙基苯酚 Phenol, 4-(2-propenyl)- | 36.42 | 501-92-8 | 0.70 | 0.79 | 2.36 | 5.54E-04 | 1.34 |
| 63 | 顺式-1-乙基-2-甲基环戊烷 Cyclopentane, 1-ethyl-2-methyl-, cis- | 29.71 | 930-89-2 | 0.04 | 0.04 | 2.08 | 0.03 | 1.04 |
| 64 | 苯 Benzene | 3.81 | 71-43-2 | 0.13 | 0.15 | 2.05 | 1.47E-03 | 1.14 |
| 65 | 二甲基硫醚 Dimethyl sulfide | 2.04 | 75-18-3 | 0.15 | 0.27 | 1.75 | 5.32E-03 | 1.23 |
| 66 | 2-乙基呋喃 Furan, 2-ethyl- | 4.08 | 3208-16-0 | 0.12 | 0.33 | 1.48 | 0.01 | 1.13 |
| 67 | L-鼠李糖 Rhamnose | 22.38 | 3615-41-6 | 0.01 | 0.21 | -2.60 | 0.03 | 1.11 |
| 合计 Sum | 1.30 | 1.89 | ||||||
新窗口打开|下载CSV
2.4.3 相对含量分析 由表2可知,酯类是PF和NF相对含量排在第1的香气物质,酯类在PF(37.69%)中的相对含量明显低于NF(57.20%),PF中相对含量最大的酯是反-2-十二碳烯醇酯(9.59%),NF中相对含量最大的酯是丁酸乙酯(17.92%)。内酯类是PF中相对含量排在第2的香气物质,其在NF中排名第3,内酯类在PF(20.91%)中的相对含量大幅高于NF(6.12%),PF中相对含量最大的内酯是δ-癸内酯(12.53%),NF中相对含量最大的内酯是丁位辛内酯(2.45%)。酮类是PF中相对含量排在第3的香气物质(15.30%),其在NF中排名第2,酮类在PF(15.30%)中的相对含量略高于NF(9.12%),PF和NF中相对含量最大的酮均为2-庚酮,相对含量分别为14.09%和7.74%。醇类、醛类及酸类在PF和NF中含量相当,醇类化合物在PF和NF中的相对含量分别为9.86%和7.51%,醛类化合物在PF和NF中的相对含量分别为8.32%和11.77%,酸类化合物在PF和NF中的相对含量分别为5.55%和5.44%。在PF和NF中,醇类、醛类和酸类相对含量最大的代谢物一致,醇类物质为辛醇,相对含量分别为5.91%(PF)和4.86%(NF);醛类物质为苯乙醛,相对含量分别为6.74%(PF)和9.22%(NF);酸类物质为辛酸,相对含量分别为5.24%(PF)和4.98%(NF)。烯烃类和其他代谢物是PF和NF中相对含量最低的代谢物,烯烃类分别为1.06%(PF)和0.98%(NF),其他类代谢物分别为1.30%(PF)和1.89%(NF)。
3 讨论
3.1 关于PF和NF的香气成分
对草莓属植物特征香气成分的鉴定是成功进行香气育种的关键,NEGRI等[2]对麝香草莓(F. moschata)和森林草莓(F. vesca)的香气成分进行比较分析;王娟等[24]鉴定出8个凤梨草莓栽培品种(系)果实的关键香气成分。草莓的特征香气可能与关键香气成分在种类及含量相互比例方面的变化有关[25],草莓野生种往往比商业品种的香气更浓郁,香气种类也更加丰富[7,26];SCHWIETERMAN等[16]对35个不同的凤梨草莓栽培品种香气多样性的调查发现,即使在最具香味的商业品种中,也不超过81种香气成分,本研究从凤梨草莓与黄毛草莓的2个种间杂种中共鉴定出383种香气成分,远远超过凤梨草莓栽培品种的香气种类,该结果进一步证实了草莓野生资源在香气育种中的价值。本研究2个种间杂种的差异代谢物中,种类和相对含量最多是酯类,其次是内酯类,因此,酯类和内酯类可能是影响PF和NF果实香气的关键物质。3.2 酯类和内酯类对PF和NF果实香气的影响
酯类对草莓果香的贡献很大[4,12]。本研究对差异代谢物的分析表明,酯类在NF中的相对含量大幅高于PF,NF中相对含量较大的酯是丁酸乙酯和3-羟基丁酸乙酯。多数研究证明凤梨草莓与野生草莓在酯的种类上存在明显差异,各有其关键的特征酯类,如黄毛草莓以苯甲酸甲酯、乙酸苄酯、肉桂酸甲酯和肉桂酸乙酯等最为显著[4,5],氨茴酸甲酯是森林草莓(F. vesca)的特征酯类[1],而丁酸甲酯、丁酸乙酯、己酸乙酯和2-甲基丁酸甲酯是凤梨草莓最重要的酯类[27,28],本研究中存在于黄毛草莓的肉桂酸乙酯在2个种间杂种中相对含量较低,可能是在子代中发生了不同程度的缺失[29]。另一方面,肉桂酸乙酯虽然含量低,但PF相对于NF下调幅度最大,因此,肉桂酸乙酯可能与丁酸乙酯和3-羟基丁酸乙酯等一起构成NF的特征香气物质。ZHAO等[6]报道了黄毛草莓的2个内酯类特征香气成分,δ-癸内酯和丙位十二内酯(γ-Dodecalactone),并指出黄毛草莓的特异蜜桃香味可能与丙位十二内酯有关。草莓中桃香味相关的物质报道较多的是γ-癸内酯,其表达受FaFAD1的调节,内酯是一种挥发性脂肪酸衍生分子,具有“水果味”“甜”或“桃子”的香气[17]。与NF相比,本研究中PF显著富集了内酯类物质,上调幅度较大的7种内酯依次是(Z)-7-癸烯-5-酸、丁位十一内酯、2H-Pyran-2-one, 5,6-dihydro-6- propyl-、丁位己内酯、丁位辛内酯和丙位壬内酯,其中δ-癸内酯、(Z)-7-癸烯-5-酸、丁位己内酯含量较高,综合分析可知,这7种内酯类物质的显著增加可能是PF蜜桃香气形成的关键因素,经
3.3 酮类、醇类、醛类及酸类等对PF和NF果实香气的影响
本研究中酮类、醇类、醛类及酸类在PF和NF中相对含量相当,硫化物等其他化合物的相对含量最少,仅占1%左右。酮类物质对草莓香气形成也有重要作用,相对含量在PF中仅次于酯类和内酯类,在NF中仅次于酯类。2,5-二甲基-4-甲氧基-3(2H)-呋喃酮普遍存在于草莓属果实中,并具有焦糖味[7],醇类和酸类对味道的影响很小,往往产生令人不愉快的气味[1,28]。某些醛类对草莓的香气很重要,有助于产生青草味,并随果实成熟而减少[12,28]。一些硫化物即使浓度很低,但是对草莓的特征香气有重要作用[7]。野生草莓的浓郁香气能否成功导入栽培种中,在很大程度上取决于能否产生可育的种间杂种后代[2],本研究中五倍体经过染色体加倍后获得的十倍体再与八倍体栽培品种小白杂交,其后代畸形果较多(因九倍体不能结实,大部分种子发育不良所致),但也会产生少量可育的偶数倍种子,偶数倍种子的产生可能与异常有丝分裂如染色体单桥、双桥、多桥、多极分裂或2n配子的形成有关[30],类似现象在草莓上也有报道[31,32]。果香受基因型和环境的综合影响[4,33],是由多基因位点控制的复杂性状[34],代谢组学在草莓香气上的研究起步较晚,相关数据库有待完善。目前,草莓属植物中仅阐明了部分香气物质积累的生物合成途径,其内在的遗传规律及调控机制并不清楚[1]。因此,有必要结合其他组学数据,以快速、准确地了解这些物质的表达信息,并把这些数据作为准确的分子性状衡量指标,结合分子生物学手段,应用于调控基因位点及关键基因挖掘、功能解析乃至新品种的培育。
4 结论
在凤梨草莓与黄毛草莓的2个种间杂种PF和NF中检测到383种代谢物,其中筛选出67种差异代谢物。酯类是NF的主要香气物质,log2FC排在前3名的依次是肉桂酸乙酯、亚硫酸、2-乙基己基异己酯和3-羟基丁酸乙酯,相对含量最高是丁酸乙酯和3-羟基丁酸乙酯;PF显著上调了内酯类物质,log2FC排在前3名的依次是(Z)-7-癸烯-5-酸、丁位十一内酯和δ-癸内酯,相对含量最高的是δ-癸内酯和丁位己内酯。肉桂酸乙酯、丁酸乙酯和3-羟基丁酸乙酯等酯类可能是NF的关键香气物质,(Z)-7-癸烯-5-酸、丁位十一内酯和δ-癸内酯等内酯类可能是PF蜜桃香气形成的关键物质,这些内酯类物质可能会成为草莓蜜桃香味育种的新目标。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:29100102 [本文引用: 6]
URLPMID:25717332 [本文引用: 4]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.scienta.2012.11.001URL [本文引用: 4]
[本文引用: 2]
[本文引用: 2]
URLPMID:24115051 [本文引用: 5]
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/j.foodchem.2016.08.089URLPMID:27664623 [本文引用: 1]

Dates (Phoenix dactylifera L.) are distributed worldwide as major food complement providing a source of sugars and dietary fiber as well as macro- and micronutrients. Although phytochemical analyses of date fruit non-volatile metabolites have been reported, much less is known about the aroma given off by the fruit, which is critical for dissecting sensory properties and quality traits. Volatile constituents from 13 date varieties grown in Egypt were profiled using SPME-GCMS coupled to multivariate data analysis to explore date fruit aroma composition and investigate potential future uses by food industry. A total of 89 volatiles were identified where lipid-derived volatiles and phenylpropanoid derivatives were the major components of date fruit aroma. Multivariate data analyses revealed that 2,3-butanediol, hexanal, hexanol and cinnamaldehyde contributed the most to classification of different varieties. This study provides the most complete map of volatiles in Egyptian date fruit, with Siwi and Sheshi varieties exhibiting the most distinct aroma among studied date varieties.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17995662 [本文引用: 3]
DOI:10.1104/pp.16.00226URLPMID:26993618 [本文引用: 1]
Strawberries emit hundreds of different volatiles, but only a dozen, including the key compound HDMF [4-hydroxy-2,5-dimethyl-3(2H)-furanone] contribute to the flavor of the fruit. However, during ripening, a considerable amount of HDMF is metabolized to the flavorless HDMF beta-d-glucoside. Here, we functionally characterize nine ripening-related UGTs (UDP-glucosyltransferases) in Fragaria that function in the glucosylation of volatile metabolites by comprehensive biochemical analyses. Some UGTs showed a rather broad substrate tolerance and glucosylated a range of aroma compounds in vitro, whereas others had a more limited substrate spectrum. The allelic UGT71K3a and b proteins and to a lesser extent UGT73B24, UGT71W2, and UGT73B23 catalyzed the glucosylation of HDMF and its structural homolog 2(or 5)-ethyl-4-hydroxy-5(or 2)-methyl-3(2H)-furanone. Site-directed mutagenesis to introduce single K458R, D445E, D343E, and V383A mutations and a double G433A/I434V mutation led to enhanced HDMF glucosylation activity compared to the wild-type enzymes. In contrast, a single mutation in the center of the plant secondary product glycosyltransferase box (A389V) reduced the enzymatic activity. Down-regulation of UGT71K3 transcript expression in strawberry receptacles led to a significant reduction in the level of HDMF-glucoside and a smaller decline in HDMF-glucoside-malonate compared with the level in control fruits. These results provide the foundation for improvement of strawberry flavor and the biotechnological production of HDMF-glucoside.
[本文引用: 1]
DOI:10.1105/tpc.104.023895URLPMID:15522848 [本文引用: 1]
The blends of flavor compounds produced by fruits serve as biological perfumes used to attract living creatures, including humans. They include hundreds of metabolites and vary in their characteristic fruit flavor composition. The molecular mechanisms by which fruit flavor and aroma compounds are gained and lost during evolution and domestication are largely unknown. Here, we report on processes that may have been responsible for the evolution of diversity in strawberry (Fragaria spp) fruit flavor components. Whereas the terpenoid profile of cultivated strawberry species is dominated by the monoterpene linalool and the sesquiterpene nerolidol, fruit of wild strawberry species emit mainly olefinic monoterpenes and myrtenyl acetate, which are not found in the cultivated species. We used cDNA microarray analysis to identify the F. ananassa Nerolidol Synthase1 (FaNES1) gene in cultivated strawberry and showed that the recombinant FaNES1 enzyme produced in Escherichia coli cells is capable of generating both linalool and nerolidol when supplied with geranyl diphosphate (GPP) or farnesyl diphosphate (FPP), respectively. Characterization of additional genes that are very similar to FaNES1 from both the wild and cultivated strawberry species (FaNES2 and F. vesca NES1) showed that only FaNES1 is exclusively present and highly expressed in the fruit of cultivated (octaploid) varieties. It encodes a protein truncated at its N terminus. Green fluorescent protein localization experiments suggest that a change in subcellular localization led to the FaNES1 enzyme encountering both GPP and FPP, allowing it to produce linalool and nerolidol. Conversely, an insertional mutation affected the expression of a terpene synthase gene that differs from that in the cultivated species (termed F. ananassa Pinene Synthase). It encodes an enzyme capable of catalyzing the biosynthesis of the typical wild species monoterpenes, such as alpha-pinene and beta-myrcene, and caused the loss of these compounds in the cultivated strawberries. The loss of alpha-pinene also further influenced the fruit flavor profile because it was no longer available as a substrate for the production of the downstream compounds myrtenol and myrtenyl acetate. This phenomenon was demonstrated by cloning and characterizing a cytochrome P450 gene (Pinene Hydroxylase) that encodes the enzyme catalyzing the C10 hydroxylation of alpha-pinene to myrtenol. The findings shed light on the molecular evolutionary mechanisms resulting in different flavor profiles that are eventually selected for in domesticated species.
DOI:10.1371/journal.pone.0088446URLPMID:24523895 [本文引用: 2]
Fresh strawberries (Fragaria x ananassa) are valued for their characteristic red color, juicy texture, distinct aroma, and sweet fruity flavor. In this study, genetic and environmentally induced variation is exploited to capture biochemically diverse strawberry fruit for metabolite profiling and consumer rating. Analyses identify fruit attributes influencing hedonics and sensory perception of strawberry fruit using a psychophysics approach. Sweetness intensity, flavor intensity, and texture liking are dependent on sugar concentrations, specific volatile compounds, and fruit firmness, respectively. Overall liking is most greatly influenced by sweetness and strawberry flavor intensity, which are undermined by environmental pressures that reduce sucrose and total volatile content. The volatile profiles among commercial strawberry varieties are complex and distinct, but a list of perceptually impactful compounds from the larger mixture is better defined. Particular esters, terpenes, and furans have the most significant fits to strawberry flavor intensity. In total, thirty-one volatile compounds are found to be significantly correlated to strawberry flavor intensity, only one of them negatively. Further analysis identifies individual volatile compounds that have an enhancing effect on perceived sweetness intensity of fruit independent of sugar content. These findings allow for consumer influence in the breeding of more desirable fruits and vegetables. Also, this approach garners insights into fruit metabolomics, flavor chemistry, and a paradigm for enhancing liking of natural or processed products.
[本文引用: 2]
URLPMID:32003918 [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2019.13.010URL [本文引用: 1]
【目的】比较日中性草莓秋季和冬季果实挥发性成分,明确季节变化对挥发性成分构成及香气值的影响,为研究草莓香气形成和开发利用日中性品种提供理论依据。【方法】以日中性品种‘阿尔比’‘蒙特瑞’‘波特拉’和‘圣安德瑞斯’的秋季大拱棚栽培和冬季日光温室栽培的成熟果实为试材,采用顶空固相微萃取和气谱-质谱联用方法检测挥发性成分并进行定性、定量分析,计算挥发性成分的数量、含量、比例和香气值,通过主成分分析对秋、冬果实香气成分含量差异做进一步分析,并依据香气值>1的成分,利用聚类热图对供试样品进行分类。【结果】共检测到88种挥发性成分,计算了57种成分的香气值,其中有30种成分的香气值>1。季节变化对各类挥发性成分数量和总含量的影响因品种而异;冬季果实中酯类成分的含量及在总挥发性成分中的比例显著高于秋季果实,而萜烯类成分的比例则显著低于秋季果实。主成分分析表明,‘波特拉’果实的挥发性成分含量受季节变化影响最小,而‘圣安德瑞斯’受季节影响最大。所有挥发性成分中,丁酸乙酯的平均香气值最高,对果实香气有重要影响;与秋季果实相比,冬季果实乙酸丁酯和丁酸甲酯的香气值显著升高;香气值>1的萜烯类成分在冬季果实中的香气值之和均低于秋季果实。丁酸乙酯、丁酸丁酯、4-甲氧基-2,5-二甲基-3(2H )-呋喃酮(DMMF)、己酸乙酯、芳樟醇等成分在不同季节果实的香气差异中起重要作用。通过热图分析可以将供试样品分为3类,其中第I类样品包括‘阿尔比’秋季果实、冬季果实和‘蒙特瑞’秋季果实,它们的总香气值和丁酸乙酯香气值均高于其他样品。【结论】季节变化主要影响果实的酯类和萜烯类成分,冬季果实的酯类含量、比例显著高于秋季果实,而萜烯类的比例则显著低于秋季果实。4个品种中,‘波特拉’果实挥发性成分含量受季节影响最小,而‘阿尔比’果实香气最浓,香气受季节变化的影响最小,‘圣安德瑞斯’挥发性成分含量和香气均受影响最大。
DOI:10.3864/j.issn.0578-1752.2019.13.010URL [本文引用: 1]
【目的】比较日中性草莓秋季和冬季果实挥发性成分,明确季节变化对挥发性成分构成及香气值的影响,为研究草莓香气形成和开发利用日中性品种提供理论依据。【方法】以日中性品种‘阿尔比’‘蒙特瑞’‘波特拉’和‘圣安德瑞斯’的秋季大拱棚栽培和冬季日光温室栽培的成熟果实为试材,采用顶空固相微萃取和气谱-质谱联用方法检测挥发性成分并进行定性、定量分析,计算挥发性成分的数量、含量、比例和香气值,通过主成分分析对秋、冬果实香气成分含量差异做进一步分析,并依据香气值>1的成分,利用聚类热图对供试样品进行分类。【结果】共检测到88种挥发性成分,计算了57种成分的香气值,其中有30种成分的香气值>1。季节变化对各类挥发性成分数量和总含量的影响因品种而异;冬季果实中酯类成分的含量及在总挥发性成分中的比例显著高于秋季果实,而萜烯类成分的比例则显著低于秋季果实。主成分分析表明,‘波特拉’果实的挥发性成分含量受季节变化影响最小,而‘圣安德瑞斯’受季节影响最大。所有挥发性成分中,丁酸乙酯的平均香气值最高,对果实香气有重要影响;与秋季果实相比,冬季果实乙酸丁酯和丁酸甲酯的香气值显著升高;香气值>1的萜烯类成分在冬季果实中的香气值之和均低于秋季果实。丁酸乙酯、丁酸丁酯、4-甲氧基-2,5-二甲基-3(2H )-呋喃酮(DMMF)、己酸乙酯、芳樟醇等成分在不同季节果实的香气差异中起重要作用。通过热图分析可以将供试样品分为3类,其中第I类样品包括‘阿尔比’秋季果实、冬季果实和‘蒙特瑞’秋季果实,它们的总香气值和丁酸乙酯香气值均高于其他样品。【结论】季节变化主要影响果实的酯类和萜烯类成分,冬季果实的酯类含量、比例显著高于秋季果实,而萜烯类的比例则显著低于秋季果实。4个品种中,‘波特拉’果实挥发性成分含量受季节影响最小,而‘阿尔比’果实香气最浓,香气受季节变化的影响最小,‘圣安德瑞斯’挥发性成分含量和香气均受影响最大。
DOI:10.1007/s10722-006-9009-4URL [本文引用: 2]
Plant breeders are interested in strawberry species as donors of volatile compounds in breeding programmes because of the diversity and intensity of wild strawberry aroma. Therefore, the topic of this paper is the prospective analysis of four accessions of four wild strawberry accessions in comparison to a standard cultivar of Fragaria × ananassa Duch. by using human sensory, gas chromatography mass spectrometry (GCMS) and gas chromatography–olfactometry (GCO). The wild species have higher aroma intensities compared with the cultivated one. The flavour quality differs significantly. Semiquantitative GC analysis revealed that F. × ananassa cv. ‘Elsanta’ has the lowest content of volatile compounds whereas Fragaria moschata L. ‘Cotta’ has the highest. The aroma impressions, measured by GCO, support the findings of GCMS analyses. The nasal impact frequency (NIF)-profiles of the wild types are more manifold and of higher intensities than those of the cultivated F. × ananassa cv. ‘Elsanta’ which corresponds with the overall flavour impression when tasting the fresh fruits.
DOI:10.1007/BF01201770URL [本文引用: 1]
DOI:10.1021/jf960366oURL [本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.scienta.2010.03.015URL [本文引用: 1]
DOI:10.1016/j.scienta.2011.10.021URL [本文引用: 1]
DOI:10.21273/HORTSCI.35.6.1022URL [本文引用: 1]
DOI:10.1126/science.1112614URLPMID:16469919 [本文引用: 1]
Plants produce many volatile metabolites. A small subset of these compounds is sensed by animals and humans, and the volatile profiles are defining elements of the distinct flavors of individual foods. Flavor volatiles are derived from an array of nutrients, including amino acids, fatty acids, and carotenoids. In tomato, almost all of the important flavor-related volatiles are derived from essential nutrients. The predominance of volatiles derived from essential nutrients and health-promoting compounds suggests that these volatiles provide important information about the nutritional makeup of foods. Evidence supporting a relation between volatile perception and nutrient or health value will be reviewed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}