Analysis of Differential Metabolites in Grains of Rice Cultivar Changbai 10 Under Salt Stress
ZHANG GuiYun,, ZHU JingWen, SUN MingFa, YAN GuoHong, LIU Kai, WAN BaiJie, DAI JinYing, ZHU GuoYong,Jiangsu Institute of Agricultural Sciences in Coastal Region, Yancheng 224001, Jiangsu
Abstract 【Objective】To reveal the influence of salt stress on metabolites and metabolism pathway in the rice seed of Changbai 10, understand the mechanism of its salt tolerance, and screen possible markers for salt tolerant rice varieties. 【Method】Gas chromatography-mass spectrometry (GC-MS) was used to analyze the changes of metabolites in the seeds (25 days after flower) of a saline tolerance japonica rice Changbai 10 under salt stress (4‰). The rice variety Yandao 8 with moderate salt tolerance was treated with the metabolite which was the highest improvement among the different metabolites to identify whether it could improve the salt tolerance. The treatment methods were as follows: Yandao 8 seeds were soaked in NaCl solution (5‰) containing the metabolite with different concentration gradient (0-400 mg·L-1), after 20 hours, the seeds were put in germination box for germination test.【Result】A total of 295 metabolites were detected in the samples. Principal component analysis (PCA) and orthogonal partial least-squares-discriminant analysis OPLS-DA were performed to visualize the metabolic difference among experimental groups. The results revealed a total of 71 metabolites were significantly changed (42 up-regulated and 29 down-regulated) under salt stress. The top ten up-regulated metabolites were 5-hydroxynorvaline (5.23 fold), Pentane-1,2,5-triol (4.10 fold), Cyanoalanine (2.97 fold), L-asparagine (2.91 fold), 2-hydroxybutanoic acid (2.86 fold), Oxoglutaric acid (2.74 fold), Pyruvic acid (2.50 fold), Ornithine (2.40 fold), Citrulline (2.07 fold), Proline (1.94 fold). And the top ten down-regulated metabolites were Erythrose (2.83 fold), Xylonolactone (2.49 fold), Aspartate (2.42 fold), Uridine 5’-monophosphate (2.37 fold), Cholesterone (1.90 fold), Hexaric acid (1.87 fold), D7-glucose (1.86 fold), 5,7-dihydroxy-4’-methoxyisoflavone (1.72 fold), Amidosulfonic acid (1.61 fold), 6-deoxygalactofuranose (1.61 fold). Among them, 15 differential metabolites were KEGG annotated and significantly(P<0.01) affected arginine biosynthesis, alanine, aspartate and glutamate metabolism, butanoate metabolism, carbon metabolism, lysine degration, biosynthesis of amino acids, glyoxylate and dicarboxylate metabolism, citrate cycle (TCA cycle) and so on. The germination rate of Yandao 8 was only 66.7% under salt stress (5‰), significantly lower than that with exogenous 5-hydroxynvaline treatment (74.3-90.3%). The result revealed that germination rate of the seeds with 200 mg·L-1 5-hydroxynvaline treatment was the highest.【Conclusion】Amino acid and organics acid were mainly up regulated, while sugar and lipid were mainly down regulated in salt treated group. The changed metabolism pathway of Arginine biosynthesis and TCA cycle might contribute to its high salt tolerance. 5-hydroxynvaline could effectively alleviate the inhibition of salt stress on rice seeds germination, and the optimal concentration is 200 mg·L-1. Keywords:salt stress;metabolome;differential metabolite;metabolic pathway;GC-MS;rice
PDF (3587KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 张桂云, 朱静雯, 孙明法, 严国红, 刘凯, 宛柏杰, 代金英, 朱国永. 盐胁迫条件下长白10号水稻籽粒中差异代谢物的分析[J]. 中国农业科学, 2021, 54(4): 675-683 doi:10.3864/j.issn.0578-1752.2021.04.001 ZHANG GuiYun, ZHU JingWen, SUN MingFa, YAN GuoHong, LIU Kai, WAN BaiJie, DAI JinYing, ZHU GuoYong. Analysis of Differential Metabolites in Grains of Rice Cultivar Changbai 10 Under Salt Stress[J]. Scientia Acricultura Sinica, 2021, 54(4): 675-683 doi:10.3864/j.issn.0578-1752.2021.04.001
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】水稻是重要的粮食作物之一,提高水稻产量和品质是确保粮食安全生产的关键问题。水稻生长发育进程中会面临各种胁迫因素的限制。近年来,各类极端气候不定时出现,土壤盐碱化问题呈日益加重的趋势。江苏沿海滩涂是省重要的土地资源,海岸泥沙淤积速度较快,每年增长面积达1 333 hm2以上。而水稻是沿海滩涂土壤改良利用的先锋作物,在盐胁迫下,体内发生了复杂的生理变化,为抵御盐害在细胞中积累大量的代谢产量,如叶片中脯氨酸含量会明显提高以调节受盐胁迫时的渗透平衡[1]。虽然基因组学、转录组学和蛋白质组学在植物逆境胁迫研究中取得很多进展,但它们很难涵盖重要的生命活性物质,细胞内许多生命活动发生在代谢物层面,如细胞信号释放、能量传递、细胞间通信等都受代谢物调控,因此代谢组学应运而生。【前人研究进展】与转录组学和蛋白质组学相比,代谢组学专门研究生物体由于基因修饰或外界环境改变引起代谢物的变化,是最接近表型的组学,也是目前组学研究领域热点之一。在干旱、高温及盐多重胁迫下,SUN等[2]利用核磁共振(nuclear magnetic resonance,NMR)技术发现玉米叶片中葡萄糖、果糖、苹果酸、柠檬酸、脯氨酸、丙氨酸、天冬氨酸、天冬酰胺酸及苏氨酸等多种代谢物发生显著变化,这些变化与TCA循环和核心代谢有关。GUO等[3]利用GC-MS对玉米幼苗在中性盐和碱性盐胁迫下代谢组的变化进行了系统分析,发现碱性盐使植物糖异生作用改变,同时抑制光合作用、氮代谢、糖酵解及多种氨基酸的合成。ZHANG等[4]利用代谢组学探究不同盐浓度对烟草植株的影响及烟草对盐胁迫的动态响应,发现盐胁迫造成三羧酸循环、谷氨酸介导的脯氨酸生物合成、莽草酸介导的次生代谢产物、核苷酸等代谢途径发生改变。WU等[5]利用GC-MS技术鉴定出野生和栽培大麦在盐胁迫下有82个差异代谢产物,其中多元醇只有在根系生长和耐盐性中发挥重要作用,野生大麦叶片中具有较高含量的叶绿素和渗透调节物质。GUO等[6]利用GC-MS分析中性盐和碱性盐胁迫对小麦代谢的影响,筛选出75个差异代谢物,主要为有机酸、氨基酸、糖、多元醇等,中性盐胁迫和碱性盐胁迫引起不同的代谢变化。碱性盐胁迫显著抑制光合速率、氮代谢、糖和氨基酸产量降低等。KIM等[7]对拟南芥盐胁迫处理0.5—72 h后进行代谢组研究,检测到氨基酸、糖、糖醇、有机酸和胺的极性代谢物等,发现短时间的盐胁迫处理会影响甲基化循环,苯甲素代谢途径和甜菜碱的生物合成,长时间的盐胁迫会诱导糖酵解、蔗糖代谢和抑制甲基化循环。【本研究切入点】前人利用代谢组学对不同植物进行了逆境胁迫研究,为植物耐盐生理机制提供了新的技术和思路[8,9]。但研究多集中在植物受到逆境胁迫后,叶片或者根中代谢产物和代谢途径发生哪些改变[10,11],且目前对水稻耐盐代谢组学的研究较少,特别是盐胁迫后对水稻籽粒代谢物的影响及代谢通路的分析仍鲜见报道。【拟解决的关键问题】本研究拟采用一耐盐碱性较好的水稻品种长白10号为研究对象,通过测定盐胁迫后对其籽粒中代谢产物的变化,探讨其应对盐胁迫的适应机制,筛选可能的可用于后续水稻品种耐盐性鉴定的标志物。
Fig. 1PCA (A) and OPLS-DA (B) scatter plots of salt stress treated and control samples
2.2 盐胁迫下籽粒中的差异代谢物
在盐胁迫处理组和对照组样品中共检测到295种代谢产物。OPLS-DA分析中,变量权重值(variable important in projection,VIP)大于1的代谢物被认为是差异代谢物,通过T检验,共筛选到存在显著差异的代谢物71个,其中,上调代谢物42个,下调代谢物29个。将所有显著差异代谢物表达量进行层次聚类(图2),可更直观地看出差异代谢物在盐胁迫组和对照组间的表达差异。其中上调代谢物的差异倍数都在1.9倍以上,最大的为5.2倍(5-羟基正缬氨酸,图3)。
A:发芽率;B:5-羟基正缬氨酸(200 mg·L-1)浸种(右图)与未处理对照(左图)在5‰盐胁迫下生长6 d对比图。**代表各处理间差异显著(P<0.01) Fig. 5Effect of exogenous 5-hydroxynvaline on seed germination under salt stress
A: Germination rate; B: Phenotypes of 5-hydroxynvaline (200 mg·L-1) presoaking seeds (right) and untreated control (left) under salt stress (5‰) for 6 days. ** represents significantly difference at P<0.01
DEMIRALT, TüRKANI. Exogenous glycinebetaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress Environmental and Experimental Botany, 2006,56(1):72-79. [本文引用: 1]
SUN CX, LI MQ, GAO XX, LIU LN, WU XF, ZHOU JH. Metabolic response of maize plants to multi-factorial abiotic stresses Plant Biology, 2016,18(1):120-129. [本文引用: 1]
GUOR, SHI LX, YAN CR, ZHONG XL, GU FX, LIUQ, XIAX, Li HR. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings BMC Plant Biology, 2017,17:41. [本文引用: 1]
ZHANG JT, ZHANGY, DU YY, CHEN SY, TANG HR. Dynamic metabonomic responses of tobacco (Nicotiana tabacum) plants to salt stress Journal of Proteome Research, 2011,10(4):1904-1914. [本文引用: 1]
WU DZ, CAI SG, CHEN MX, YE LZ, CHEN ZH, ZHANG HT, DAIF, WU FB, ZHANG GP. Tissue metabolic responses to salt stress in wild and cultivated barley PLoS ONE, 2013,8(1):e55431. [本文引用: 1]
GUOR, YANG ZZ, LIF, YAN CR, ZHONG XL, LIUQ, XIAX, LI HR, ZHAOL. Comparative metabolic responses and adaptive strategies of wheat (Triticum aestivum) to salt and alkali stress BMC Plant Biology, 2015,15:170. [本文引用: 1]
KIM JK, BAMBAT, HARADAK, FUKUSAKIE, KOBAYASHIA. Time-course metabolic profiling in Arabidopsis thaliana cell cultures after salt stress treatment Journal of Experimental Botany, 2006,58(3):415-424. [本文引用: 1]
NI JW, YANG XY, ZHANG HX, NI YY, WU HW, WEIQ. Metabolimics and its application in the crop research under abiotic stress World Forestry Research, 2014,27(5):11-17. (in Chinese) [本文引用: 1]
ZHANG XL, ZHANG RY. Metabolimics and its application in the crop research Letters in Biotechnology, 2018,29(3):446-450. (in Chinese) [本文引用: 1]
OBATAT, WITTS, LISECJ, PALACIOS-ROJASN, FLOREZ- SARASAI, YOUSFIS, ARAUS JL, CAIRNS JE, FERNIE AR. Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield Plant Physiology, 2015,169(4):2665-2683. [本文引用: 2]
DARKOE, GIERCZIKK, HUDAKO, FORGOP, PALM, TURKOSIE, KOVACSV, DULAIS, MAJLATHI, MOLNARI, JANDAT, MOLNAR-LANGM. Differing metabolic responses to salt stress in wheat-barley addition lines containing different 7H chromosomal fragments PLoS ONE, 2017,12(3):e0174170. [本文引用: 2]
WANGL. Metabolimics research in transgenic rice [D]. Beijing: Beijing University of Chemical Technology, 2013. (in Chinese) [本文引用: 1]
MUKHERJEES, SENGUPTAS, MUKHERJEEA, BASAKP, MAJUMDER AL. Abiotic stress regulates expression of galactinol synthase genes post-transcriptionally through intron retention in rice Planta, 2019,249(3):891-912. [本文引用: 1]
TANGY, BAOX, ZHIY, WUQ, GUOY, YINX, ZENGL, LIJ, ZHANGJ, HEW, LIUW, WANGQ, JIAC, LIZ, LIUK. Overexpression of a MYB family gene, OsMYB6, increases drought and salinity stress tolerance in transgenic rice Frontiers in Plant Science, 2019,10:168. [本文引用: 1]
ISLAM MO, KATOH, SHIMAS, TEZUKAD, MATSUIH, IMAIR. Functional identification of a rice trehalase gene involved in salt stress tolerance Gene, 2019,685:42-49. [本文引用: 2]
PAIVA A LS, PASSAIAG, LOBO A KM, JARIM-MESSEDERD, SILVEIRA J AG, MARGIS-PINHEIROM. Mitochondrial glutathione peroxidase (OsGPX3) has a crucial role in rice protection against salt stress Environmental and Experimental Botany, 2019,158:12-21. [本文引用: 1]
LIZ, FUX, TIANY, XUJ, GAOJ, WANGB, HANH, WANGL, ZHANGF, ZHUY, HUANGY, PENGR, YAOQ. Overexpression of a trypanothione synthetase gene from Trypanosoma cruzi, TcTrys, confers enhanced tolerance to multiple abiotic stresses in rice Gene, 2019,710:279-290. [本文引用: 1]
ZHANGA, LIUY, WANGF, LIT, CHENZ, KONGD, BIJ, ZHANGF, LUOX, WANGJ, TANGJ, YUX, LIUG, LUOL. Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene Molecular Breeding, 2019,39(3):47. [本文引用: 1]
HUANG XY, CHAO DY, GAO JP, ZHU MZ, SHIM, LIN HX. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control Genes Development, 2009,23(15):1805-1817. [本文引用: 1]
YANJ, LIPKA AE, SCHMELZ EA, BUCKLER ES, JANDERG. Accumulation of 5-hydroxynorvaline in maize (Zea mays) leaves is induced by insect feeding and abiotic stress Journal of Experimental Botany, 2014,66(2):593-602. [本文引用: 1]
YU LL, LIUY, XUF. The source of cyanide and its function in plants Chinese Bulletin of Life Sciences, 2019,31(2):12-18. (in Chinese) [本文引用: 1]
FORLANIG, BERTAZZINIM, CAGNANOG. Stress-driven increase in proline levels, and not proline levels themselves, correlates with the ability to withstand excess salt in a group of 17 Italian rice genotypes Plant Biology, 2019,21(2):336-342. [本文引用: 1]
GERONA M EB, DEOCAMPO MP, EGDANE JA, ISMAIL AM, DIONISIO-SESE ML. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage Rice Science, 2019,26(4):207-219. [本文引用: 1]
LIUJ, SHI DC. Photosynthesis, chlorophyll fluorescence, inorganic ion and organic acid accumulations of sunflower in responses to salt and salt-alkaline mixed stress Photosynthetica, 2010,48(1):127-134. [本文引用: 1]
ZHOU GY, ZHAI CJ, DENG XL, ZHANGJ, ZHANG ZL, DAI QG, CUI SY. Performance of yield, photosynthesis and grain quality of japonica rice cultivars under salinity stress in micro-plots Chinese Journal of Rice Science, 2018,32(2):146-154. (in Chinese) [本文引用: 1]
XIE LH, LUOJ, TANG SQ, CHENN, JIAO GA, SHAO GN, WEI XJ, HU PS. Proteins affect rice eating quality properties and its mechanism Chinese Journal of Rice Science, 2013,27(1):91-96. (in Chinese) [本文引用: 1]
AHMADP, ABDEL LATEF AA, HASHEMA, ABD_ALLAH EF, GUCELS, TRAN L SP. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea Frontiers in Plant Science, 2016,7:347. [本文引用: 1]
JOSEPH EA, RADHAKRISHNAN VV, MOHANAN KV. Variation in total polyamine content in some native rice cultivars of North Kerala, India in response to salinity stress Biotechnology, 2016,9(5):731-738. [本文引用: 1]
KHUSHBOOK, SHEKHAWAT GS. Nitric oxide improved salt stress tolerance by osmolyte accumulation and activation of antioxidant defense system in seedling of B. juncea (L.) Czern Vegetos, 2019,32:583-592. [本文引用: 1]
YASTREB TO, KOLUPAEV YE, KARPETS YV, DMITRIEV AP. Effect of nitric oxide donor on salt resistance of Arabidopsis jin1 mutants and wild-type plants Russian Journal of Plant Physiology, 2017,64(2):207-214. [本文引用: 1]
MARTINEZ-REYESI, DIEBOLD LP, KONGH, SCHIEBERM, HUANGH, HENSLEY CT, MEHTA MM, WANGT, SANTOS JH, WOYCHIKR, DUFOURE, SPELBRINK JN, WEINBERG SE, ZHAOY, DEBERARDINIS RJ, CHANDEL NS. TCA Cycle and mitochondrial membrane potential are necessary for diverse biological functions Molecular Cell, 2016,61(2):199-209. [本文引用: 1]
ZHANGY, BEARD K FM, SWARTC, BERGMANN S, KRAHNERT I, NIKOLOSKIZ, GRAFA, RATCLIFFE RG, SWEETLOVE LJ, FERNIE AR, OBATAT. Protein-protein interactions and metabolite channelling in the plant tricarboxylic acid cycle Nature Communications, 2017,8(1):15212. [本文引用: 1]
 ,, 朱静雯, 孙明法, 严国红, 刘凯, 宛柏杰, 代金英, 朱国永
,, 朱静雯, 孙明法, 严国红, 刘凯, 宛柏杰, 代金英, 朱国永

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT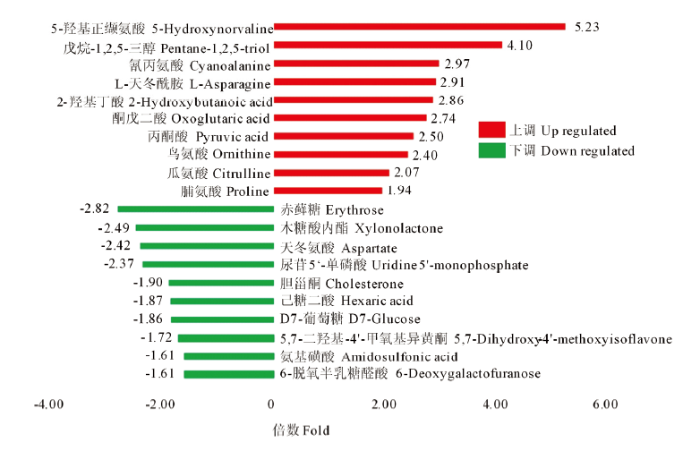 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT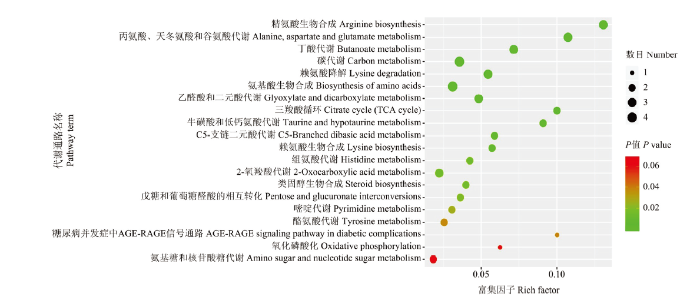 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT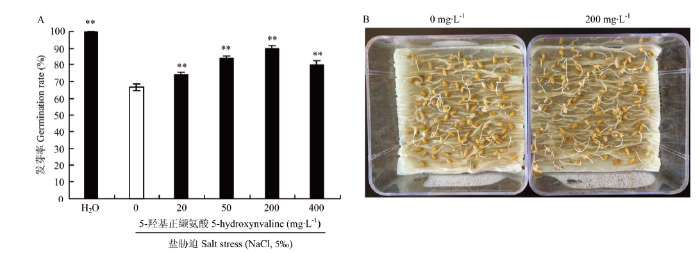 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}