 ,, 魏帅,, 冯宇霞, 钱伟,西南大学农学与生物科技学院/西南大学农业科学研究院,重庆 400716
,, 魏帅,, 冯宇霞, 钱伟,西南大学农学与生物科技学院/西南大学农业科学研究院,重庆 400716Creating a New-Type Brassica napus (AnArCnCo) with High Drought-resistance Employing Hexaploid (AnAnCnCnCoCo) as a Bridge
WAN HuaFang,, WEI Shuai,, FENG YuXia, QIAN Wei,College of Agronomy and Biotechnology, Southwest University/Academy of Agricultural Sciences, Southwest University, Chongqing 400716通讯作者:
责任编辑: 李莉
收稿日期:2019-10-5接受日期:2020-02-21网络出版日期:2020-08-16
| 基金资助: |
Received:2019-10-5Accepted:2020-02-21Online:2020-08-16
作者简介 About authors
万华方,E-mail:
魏帅,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1342KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
万华方, 魏帅, 冯宇霞, 钱伟. 以六倍体(AnAnCnCnCoCo)为桥梁创制抗旱新型甘蓝型油菜(AnArCnCo) [J]. 中国农业科学, 2020, 53(16): 3225-3234 doi:10.3864/j.issn.0578-1752.2020.16.003
WAN HuaFang, WEI Shuai, FENG YuXia, QIAN Wei.
0 引言
【研究意义】甘蓝型油菜(Brassica napus,2n=38,AACC)大约起源于7 500年前,是由白菜型油菜(Brassica rapa,2n=20,AA)和甘蓝(Brassica oleracea,2n=18,CC)杂交,并自然加倍形成的异源四倍体物种[1,2],是温带地区第二重要的油料作物[3]。干旱严重影响作物(包括油菜)的生长发育和经济产量[4]。随着菜籽油需求量的不断上升,油菜产业受到干旱的威胁日益严重[5],仅中国长江流域的油菜受秋旱影响就可减产20%左右[6]。因此,改良甘蓝型油菜的抗旱性是一个亟待解决的科学问题。然而,有限的原始亲本生态型、暂无其野生群体,以及“双低”品质选育过程引起的种质资源遗失,致甘蓝型油菜遗传背景狭窄[2,7-9],进而限制了甘蓝型油菜的抗旱育种研究[10]。利用其祖先种向甘蓝型油菜导入外源遗传成分是拓展其遗传背景的有效途径[11,12,13,14,15]。白菜型油菜A基因组的遗传渗入有利于拓展甘蓝型油菜的遗传背景[16]。白菜型油菜的农艺性状变异丰富[17,18,19],并具有耐迟播、生育期短、耐贫瘠、耐干旱等突出优点[20],是甘蓝型油菜遗传改良的重要资源[11,21-23]。因此,进行白菜型油菜抗旱性资源的鉴定,并以其为亲本合成具有优良抗旱性的新型甘蓝型油菜,对于提高油菜抗旱性能具有重要意义。【前人研究进展】利用亲本物种丰富的遗传变异拓宽遗传背景是改良现有甘蓝型油菜的一个重要方法[24]。RIPLEY等[25]利用甘蓝与甘蓝型油菜杂交,成功将甘蓝的自交不亲和性导入甘蓝型油菜。岳芳等[26]以六倍体(ArArAnAnCnCn)与甘蓝种间杂交,合成了新型甘蓝型油菜。钱伟等[27]以甘蓝型油菜和甘蓝为亲本,创建了六倍体(AnAnCnCnCoCo),并以其为桥梁,利用白菜型油菜丰富的遗传资源拓宽了现有甘蓝型油菜的遗传背景。MEI等[28]以六倍体(AnAnCnCnCoCo)作为桥梁,与甘蓝型油菜不断回交,获得了转移野生甘蓝菌核病抗性的新型甘蓝型油菜。李勤菲等[29]以六倍体(AnAnCnCnCoCo)为桥梁,将甘蓝型油菜亲本种的优良性状导入甘蓝型油菜中。【本研究切入点】甘蓝型油菜相对狭窄的遗传背景限制了其抗旱育种。耐干旱的白菜型油菜是重要的抗旱资源,但以其与甘蓝型油菜杂交、自交或回交选育新型甘蓝型油菜所需的周期长,效率低[29]。六倍体是拓宽甘蓝型油菜遗传背景的新材料[27,28,29],但以其为桥梁,转移白菜型油菜优良抗旱性的研究鲜有报道。【拟解决的关键问题】本研究通过PEG-6000溶液模拟干旱,评价白菜型油菜的抗旱性,并以六倍体材料为桥梁,以抗旱的白菜型油菜为抗性父本,创建具有优良抗旱性的新型甘蓝型油菜,为甘蓝型油菜抗旱育种提供优良的种质资源。1 材料与方法
1.1 试验材料
以11份六倍体作为母本(电子附表1)、68份白菜型油菜(电子附表2)作为父本,创建新型甘蓝型油菜。9份白菜型油菜、9份新型甘蓝型油菜和甘蓝型油菜中双11号(抗旱性好,种植广泛)为抗旱性分析材料(电子附表3),以中双11号为对照。
1.2 PEG-6000模拟干旱
参照SHANGGUAN等[30]方法,设0(CK)、100、150、200、250和300 g·L-1 6个PEG-6000质量浓度,对不同的9份白菜型油菜在萌发期进行模拟干旱处理;设0(CK)、100、150、200、250、300、350、400和600 g·L-1 PEG-6000质量浓度,对9份新型甘蓝型油菜和中双11号在萌发期进行模拟干旱处理。分别筛选用于白菜型油菜和甘蓝型油菜抗旱性鉴定的PEG-6000最适浓度。取直径9 cm的培养皿,铺垫双层滤纸,并以相应浓度的PEG-6000溶液5 mL浸润滤纸;取种子30粒均匀放置于上述浸润过的滤纸上,并盖上培养皿盖,置于组培室(25℃黑暗8 h/27℃光照16 h)培养,每天定时向培养皿中加入对应浓度的1 mL PEG-6000溶液,每个处理2个重复。以清水浸泡滤纸为对照。
1.3 检测指标
参照陈郡雯等[31]方法进行指标检测。每天定时观察种子萌发的粒数,测量胚根部位长度,以其为种子直径的一半为萌发标准。按照萌发的基本情况,以第3天的种子萌发数值计算萌发势和相对萌发势,以第8天的发芽数值计算萌发率和相对萌发率,并且在第8天随机选出10株,计算其相对根长和相对胚轴长。种子萌发抗旱指数(drought resistance index,DRI)=水分胁迫下种子萌发指数(plumule index of stress,PIS)/对照种子萌发指数(plumule index of control,PIC);萌发指数(plumule index,PI)=1nd2+ 0.75nd4+0.5nd6+0.25nd8(nd2、nd4、nd6、nd8相对应的是当天的种子萌发率),包括PIS和PIC类型;
萌发率(germination rate,GR)=正常萌发的种子数/供试种子的总数×100%;
相对萌发率(relaltive germination rate,RGR)=(处理萌发率/对照萌发率)×100%;
萌发势(sprout potential,SP)=达到高峰(第3天萌发的种子数)时正常萌发的种子数/供试种子的总数;
相对萌发势(relative sprout potential,RSP)=(处理萌发势/对照萌发势)×100%;
相对根长(relative root length,RRL)=(处理根长/对照根长)×100%;
相对胚轴长(relative stem length,RSL)=(处理胚轴长/对照胚轴长)×100%。
1.4 数据分析与处理
采用SAS v8.0软件进行数据分析。参照陈郡雯等[31]方法进行综合隶属函数值法分析,公式为R(Xi)= (Xi—Xmin)/(Xmax—Xmin)。反函数隶属函数值计算公式为R(Xi)=1—(Xi—Xmin)/(Xmax—Xmin),式中Xi指试验数据,Xmax、Xmin表示的是所对应数据的最大值和最小值。2 结果
2.1 新型甘蓝型油菜的鉴定
以11份六倍体与68份白菜型油菜杂交,获得124份新型甘蓝型油菜。在苗期(图1-Ⅰ、图1-Ⅱ和图1-Ⅲ),新型甘蓝型油菜的表型介于两亲本之间。体细胞观察结果表明,所利用的六倍体的体细胞染色体数目为56(图1-Ⅳ),而合成的新型甘蓝型油菜的体细胞染色体数目为38(图1-Ⅴ)。花粉育性结果表明,六倍体的花粉育性约90%(图1-Ⅵ),新型甘蓝型油菜的花粉育性约80%(图1-Ⅶ)。在苗期,随机选择59份长势优良的新型甘蓝型油菜、7份六倍体、10份白菜型油菜以及20份自然甘蓝型油菜提取DNA进行分子标记鉴定,并进行主成分分析。第一、第二和第三主成分的贡献率分别为13.92%、11.59%和9.09%。主成分分析将所有的供试材料分成3类(图1-Ⅷ),即白菜型油菜、六倍体、甘蓝型油菜(包括新型甘蓝型油菜和自然甘蓝型油菜),但新型甘蓝型油菜与自然甘蓝型油菜明显分开,表明以六倍体为桥梁创建新型甘蓝型油菜切实可行。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1新型甘蓝型油菜的鉴定
Ⅰ:六倍体(AnAnCnCnCoCo)的苗期表型;Ⅱ:新型甘蓝型油菜的苗期表型;Ⅲ:白菜型油菜的苗期表型;Ⅳ:六倍体体细胞染色体;Ⅴ:新型甘蓝型油菜体细胞染色体;Ⅵ:六倍体花分育性;Ⅶ:新型甘蓝型油菜花粉育性;Ⅷ:主成分分析,其中“圆形”为白菜型油菜;“红色方块”为六倍体;“绿色菱形”为新型甘蓝型油菜;“蓝色三角形”为自然甘蓝型油菜
Fig. 1Identification of new-type B. napus
Ⅰ: Seedling phenotype of hexaploid (AnAnCnCnCoCo); Ⅱ: Seedling phenotype of the new-type B. napus; Ⅲ: Seedling phenotype of B. rapa; Ⅳ: Chrosomes of hexaploid; Ⅴ: Chrosomes of new-type B. napus; Ⅵ: Pollen stainability of hexaploid; Ⅶ: Pollen stainability of new-type B. napus; Ⅷ: Principal component analysis result, “round” refers to B. rapa; “red square” refers to a hexaploid; “green diamond” refers to new-type B. napus; “blue triangle” refers to natural B. napus
2.2 筛选合适的PEG-6000浓度
依据染色体数目、花粉育性,从59份新型甘蓝型油菜中选取9份进行模拟抗旱研究(图2),在不同浓度PEG-6000溶液的处理下,油菜种子萌发均受到不同程度的抑制,并且随着浓度梯度增加,平均根长随之降低。当PEG-6000浓度达到250 g·L-1时,9份新型甘蓝型油菜与中双11号的平均根长为对照的40.2%(图2-Ⅰ),根长相对值变异系数最大,表明不同材料的平均根长有显著性差异,250 g·L-1的处理浓度为新型甘蓝型油菜模拟干旱的最适处理浓度(图2-Ⅱ)。当PEG-6000浓度达到200 g·L-1时,白菜型油菜的平均根长为对照的30.4%(图2-Ⅲ),白菜型油菜根长相对值变异系数最大,表明不同材料的平均根长有显著性差异,200 g·L-1的处理浓度为白菜型油菜模拟干旱的最适处理浓度(图2-Ⅳ)。图2
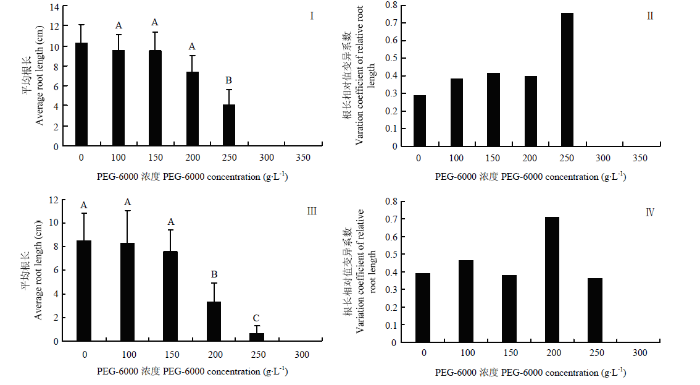 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2油菜在不同浓度PEG-6000条件下根长的变化
Ⅰ:不同浓度PEG-6000处理下新型甘蓝型油菜的平均根长;Ⅱ:不同浓度PEG-6000处理下新型甘蓝型油菜根长的相对值变异系数;Ⅲ:不同浓度PEG-6000处理下白菜型油菜的平均根长;Ⅳ:不同浓度PEG-6000处理下白菜型油菜根长的相对值变异系数。不同大写字母表示差异达到极显著水平(P<0.01)
Fig. 2Root length of rapeseed treated by PEG-6000 solution with different concentration
Ⅰ: Average root length of new-type B. napus treated by PEG-6000 solution with different concentration;Ⅱ: Variation coefficient of relative root length of the new-type B. napus treated by PEG-6000 solution with different concentration;Ⅲ: Average root length of B. rapa treated by PEG-6000 solution with different concentration; Ⅳ: Variation coefficient of relative root length of B. rapa treated by PEG-6000 solution with different concentration. Different letters indicate significant difference at P<0.01
2.3 油菜萌发期抗旱性相关指标测定及主成分分析
使用250 g·L-1的PEG-6000溶液对新型甘蓝型油菜和中双11号进行处理,用200 g·L-1的PEG-6000溶液对白菜型油菜进行处理,统计相对萌发势、相对萌发率、相对根长、相对胚轴长以及萌发抗旱指数,构成主成分分析的原始矩阵(表1)。Table 1
表1
表1油菜萌发期抗旱性相关指标主成分分析原始矩阵
Table 1
| 编号 Code | 类型 Type | 萌发抗旱指数 DRI | 相对萌发率 RGR | 相对萌发势 RSP | 相对根长 RRL | 相对胚轴长 RSL |
|---|---|---|---|---|---|---|
| 9X006 | 新型甘蓝型油菜New-type B. napus | 0.262 | 0.067 | 0.069 | 0.209 | 0.106 |
| 9X002 | 新型甘蓝型油菜New-type B. napus | 0.543 | 0.067 | 0.000 | 0.690 | 0.133 |
| 9X025 | 新型甘蓝型油菜New-type B. napus | 0.798 | 0.300 | 0.296 | 0.718 | 0.135 |
| 9X016 | 新型甘蓝型油菜New-type B. napus | 0.392 | 0.600 | 0.615 | 0.319 | 0.173 |
| 9X018 | 新型甘蓝型油菜New-type B. napus | 0.824 | 0.900 | 0.827 | 0.692 | 0.249 |
| 9X111 | 新型甘蓝型油菜New-type B. napus | 0.483 | 0.633 | 0.393 | 0.748 | 0.162 |
| 9X028 | 新型甘蓝型油菜New-type B. napus | 0.591 | 0.933 | 0.933 | 0.156 | 0.109 |
| 9X015 | 新型甘蓝型油菜New-type B. napus | 0.637 | 0.600 | 0.533 | 0.228 | 0.151 |
| 9X026 | 新型甘蓝型油菜New-type B. napus | 0.380 | 0.700 | 0.633 | 0.122 | 0.109 |
| 中双11 ZS11 | 甘蓝型油菜B. napus | 0.800 | 0.867 | 0.700 | 0.185 | 0.117 |
| 8M664 | 白菜型油菜B. rapa | 0.800 | 0.830 | 0.860 | 0.690 | 0.210 |
| 8M681 | 白菜型油菜B. rapa | 0.670 | 0. 830 | 0.800 | 0.920 | 0.410 |
| 8M625 | 白菜型油菜B. rapa | 0.530 | 0.570 | 0.530 | 0.310 | 0.260 |
| 8M623 | 白菜型油菜B. rapa | 0.520 | 0.900 | 0.830 | 0.610 | 0.500 |
| 8M684 | 白菜型油菜B. rapa | 0.290 | 0.470 | 0.410 | 0.560 | 0.130 |
| 8M624 | 白菜型油菜B. rapa | 0.370 | 0.700 | 0.500 | 0.510 | 0.210 |
| 8M693 | 白菜型油菜B. rapa | 0.450 | 0.730 | 0.730 | 0.690 | 0.300 |
| 8M191 | 白菜型油菜B. rapa | 0.300 | 0.600 | 0.480 | 0.500 | 0.810 |
| 8M655 | 白菜型油菜B. rapa | 0.420 | 0.730 | 0.660 | 0.690 | 0.250 |
新窗口打开|下载CSV
使用250 g·L-1的PEG-6000溶液对新型甘蓝型油菜进行模拟干旱处理,选用5个指标的相对值进行主成分分析(表2),第一、第二、第三主成分的贡献率分别为48.62%、35.47%和11.93%。以200 g·L-1的PEG-6000溶液对白菜型油菜进行模拟干旱处理,对5个指标进行主成分分析。第一、第二、第三主成分的贡献率分别为71.61%、19.01%和6.43%。由此可见,第一、第二和第三主成分所包含的信息集中反映了甘蓝型油菜和白菜型油菜萌发期抗旱性的各项指标在干旱胁迫环境下的相互关系,由此建立起来的评价体系可用于甘蓝型油菜和白菜型油菜萌发期的抗旱能力评价。
Table 2
表2
表2模拟干旱相关指标的主成分分析
Table 2
| 主成分 Principal component | 类别 Category | 特征值 Eigen value | 贡献率Contribution ratio (%) | 累计贡献率Accumulated contribution ratio (%) | 抗旱指标特征向量Eigen vector of measured indicators | ||||
|---|---|---|---|---|---|---|---|---|---|
| 萌发抗旱指数DRI | 相对萌发率RGR | 相对萌发势RSP | 相对根长RRL | 相对胚轴长RSL | |||||
| PC1 | 新型甘蓝型油菜 New-type B. napus | 2.431 | 48.620 | 48.620 | 0.416 | 0.597 | 0.589 | -0.024 | 0.350 |
| PC2 | 新型甘蓝型油菜 New-type B. napus | 1.774 | 35.470 | 84.090 | 0.332 | -0.224 | -0.274 | 0.720 | 0.497 |
| PC3 | 新型甘蓝型油菜 New-type B. napus | 0.597 | 11.930 | 96.020 | -0.784 | 0.083 | 0.110 | 0.011 | 0.605 |
| PC1 | 白菜型油菜 B. rapa | 3.580 | 71.610 | 71.600 | 0.459 | 0.512 | 0.520 | 0.468 | 0.194 |
| PC2 | 白菜型油菜 B. rapa | 0.950 | 19.010 | 90.600 | -0.313 | 0.08 | -0.073 | -0.038 | 0.946 |
| PC3 | 白菜型油菜 B. rapa | 0.320 | 6.430 | 97.000 | 0.599 | -0.001 | 0.103 | -0.775 | 0.175 |
新窗口打开|下载CSV
2.4 油菜萌发期抗旱性隶属函数分析
运用综合隶属函数法对新型甘蓝型油菜与白菜型油菜萌发期模拟抗旱检测指标进行分析,并以5个抗旱指标的平均值为指标评价抗旱性(表3)。结果表明,白菜型油菜8M664、8M655、8M693相比其他白菜型油菜,均具有更好的抗旱性。新型甘蓝型油菜9X018、9X015、9X016的抗旱性优于中双11号,9X111的抗旱性基本与中双11号持平,其他供试材料的抗旱性均低于中双11号。Table 3
表3
表3模拟干旱相关指标的隶属函数值
Table 3
| 编号 Code | 类型 Type | 萌发抗旱指数 DRI | 相对萌发率 RGR | 相对萌发势RSP | 相对根长 RRL | 相对胚轴长 RSL | 均值 Means |
|---|---|---|---|---|---|---|---|
| 9X006 | 新型甘蓝型油菜New-type B. napus | 1.00 | 0.00 | 0.01 | 0.73 | 0.20 | 0.388 |
| 9X002 | 新型甘蓝型油菜New-type B. napus | 0.78 | 0.09 | 0.00 | 1.00 | 0.19 | 0.412 |
| 9X025 | 新型甘蓝型油菜New-type B. napus | 1.00 | 0.25 | 0.24 | 0.88 | 0.00 | 0.474 |
| 9X026 | 新型甘蓝型油菜New-type B. napus | 0.46 | 1.00 | 0.89 | 0.02 | 0.00 | 0.474 |
| 9X028 | 新型甘蓝型油菜New-type B. napus | 0.58 | 1.00 | 1.00 | 0.05 | 0.00 | 0.526 |
| 9X111 | 新型甘蓝型油菜New-type B. napus | 0.55 | 0.80 | 0.39 | 1.00 | 0.00 | 0.548 |
| 中双11 ZS11 | 甘蓝型油菜B. napus | 0.91 | 1.00 | 0.78 | 0.09 | 0.00 | 0.556 |
| 9X016 | 新型甘蓝型油菜New-type B. napus | 0.50 | 0.96 | 1.00 | 0.33 | 0.00 | 0.558 |
| 9X015 | 新型甘蓝型油菜New-type B. napus | 1.00 | 0.92 | 0.78 | 0.15 | 0.00 | 0.570 |
| 9X018 | 新型甘蓝型油菜New-type B. napus | 0.88 | 1.00 | 0.89 | 0.68 | 0.00 | 0.690 |
| 8M623 | 白菜型油菜B. rapa | 0.07 | 1.00 | 0.83 | 0.28 | 0.00 | 0.436 |
| 8M191 | 白菜型油菜B. rapa | 0.00 | 0.61 | 0.35 | 0.40 | 1.00 | 0.472 |
| 8M624 | 白菜型油菜B. rapa | 0.31 | 1.00 | 0.59 | 0.62 | 0.00 | 0.504 |
| 8M684 | 白菜型油菜B. rapa | 0.37 | 0.78 | 0.66 | 1.00 | 0.00 | 0.562 |
| 8M625 | 白菜型油菜B. rapa | 0.88 | 1.00 | 0.89 | 0.17 | 0.00 | 0.588 |
| 8M681 | 白菜型油菜B. rapa | 0.51 | 0.83 | 0.76 | 1.00 | 0.00 | 0.620 |
| 8M655 | 白菜型油菜B. rapa | 0.35 | 1.00 | 0.84 | 0.91 | 0.00 | 0.620 |
| 8M693 | 白菜型油菜B. rapa | 0.36 | 1.00 | 1.00 | 0.90 | 0.00 | 0.652 |
| 8M664 | 白菜型油菜B. rapa | 0.90 | 0.96 | 1.00 | 0.73 | 0.00 | 0.721 |
新窗口打开|下载CSV
通过对新型甘蓝型油菜9X111、9X016、9X015和9X018及其父本白菜型油菜8M693、8M664、8M655和8M664的隶属函数值进行相关性分析(表4),发现9X111与其父本8M693呈现正相关(r= 0.672),9X016与其父本8M664呈现显著正相关(r=0.864,P<0.05),9X015与其父本8M655呈现正相关(r=0.372),9X018与其父本8M664呈现显著正相关(r=0.991,P<0.01)。结果表明,以六倍体为桥梁,可将白菜型油菜的优良抗旱性导入新型甘蓝型油菜,进而获得抗旱性提高的甘蓝型油菜。
Table 4
表4
表4模拟干旱隶属函数值的相关性分析
Table 4
| 编号Code | 9X006 | 9X002 | 9X025 | 9X026 | 9X028 | 9X111 | 中双11 ZS 11 | 9X016 | 9X015 | 9X018 |
|---|---|---|---|---|---|---|---|---|---|---|
| 8M623 | -0.686 | -0.583 | -0.336 | 0.874* | 0.833* | 0.354 | 0.610 | 0.909* | 0.522 | 0.666 |
| 8M191 | -0.631 | -0.497 | -0.821* | -0.274 | -0.362 | -0.475 | -0.563 | -0.398 | -0.665 | -0.734 |
| 8M624 | -0.309 | -0.129 | 0.081 | 0.692 | 0.651 | 0.764 | 0.560 | 0.791 | 0.503 | 0.789 |
| 8M684 | 0.023 | 0.289 | 0.398 | 0.311 | 0.300 | 0.912* | 0.216 | 0.539 | 0.201 | 0.695 |
| 8M625 | -0.151 | -0.317 | 0.122 | 0.923** | 0.948** | 0.265 | 0.992** | 0.891* | 0.986** | 0.872* |
| 8M681 | 0.064 | 0.28 | 0.455 | 0.400 | 0.402 | 0.906* | 0.341 | 0.622 | 0.347* | 0.793 |
| 8M655 | -0.232 | -0.005 | 0.177 | 0.569 | 0.547 | 0.805 | 0.409 | 0.748 | 0.372 | 0.775 |
| 8M693 | -0.291 | -0.089 | 0.122 | 0.624 | 0.611 | 0.721 | 0.440 | 0.805 | 0.404 | 0.784 |
| 8M664 | 0.067 | 0.052 | 0.441 | 0.742 | 0.778 | 0.651 | 0.800 | 0.864* | 0.818* | 0.991** |
新窗口打开|下载CSV
3 讨论
甘蓝型油菜是由甘蓝和白菜型油菜杂交,经自然加倍而成。由于其栽培历史短,相对于其亲本种而言,甘蓝型油菜遗传背景较为狭窄[1]。利用甘蓝型油菜亲本种丰富的遗传变异扩展其遗传背景已有不少报道。甘蓝型油菜与甘蓝杂交后不断自交,合成的甘蓝型油菜遗传多样性更丰富[32]。QUAZI等[33]利用甘蓝与甘蓝型油菜杂交,成功将羽衣甘蓝的蚜虫抗性转移到甘蓝型油菜;DING等[34]通过人工合成甘蓝型油菜的途径,将甘蓝亲本的菌核病抗性导入甘蓝型油菜。甘蓝型油菜另一祖先种—白菜型油菜分别独立起源于欧洲和亚洲,具有丰富的遗传变异[18],具备耐贫瘠、耐干旱、抗寒性强、生育期短等特性,为甘蓝型油菜遗传改良提供了丰富的遗传资源[20]。甘蓝型油菜A亚基因组比C亚基因组变异更丰富,白菜型油菜A亚基因组成分对于甘蓝型油菜的遗传变异有很大的影响,其遗传渗入有利于改变甘蓝型油菜遗传背景[16]。利用白菜型油菜与甘蓝型油菜杂交创建新型甘蓝型油菜的策略已在甘蓝型油菜育种中得到应用[35],但此策略中仍需要进行胚挽救,且从后代中选择获得甘蓝型油菜的周期长,效率低。因此,利用其他策略转移白菜型油菜的优良性状到甘蓝型油菜具有重要意义。XIAO等[35]利用埃塞俄比亚芥(B. carinata,BBCC,2n=34)与白菜型油菜杂交,经染色体加倍获得六倍体AABBCC,再与甘蓝型油菜杂交获得五倍体AABCC,而后进行自交,可获得新型甘蓝型油菜。近些年,许多研究者提出了利用六倍体为桥梁改良甘蓝型油菜的方法。钱伟等[27]发明了一种以甘蓝型油菜与甘蓝杂交创建六倍体AnAnCnCnCoCo的方法,并以此六倍体材料为桥梁与大量白菜型油菜杂交,同时导入甘蓝和白菜型油菜的遗传成分,有效拓宽了现有甘蓝型油菜的遗传背景。随后有许多的研究者运用此方法,以六倍体材料为桥梁合成新型甘蓝型油菜。MEI等[28]、刘瑶等[36]利用抗菌核病的甘蓝与甘蓝型油菜杂交获得六倍体,利用此六倍体与甘蓝型油菜回交,创建了菌核病抗性改良的新型甘蓝型油菜。以此六倍体与白菜型油菜杂交,进一步拓宽了甘蓝型油菜遗传背景,且获得了具有菌核病抗性、含白菜型油菜遗传成分的新型甘蓝型油菜[29]。本研究利用的六倍体材料(AnAnCnCnCoCo),前期导入了甘蓝的遗传成分,有效增加了遗传多样性,且以此六倍体与具备优良抗旱性的白菜型油菜(ArAr)杂交,可以快速获得新型甘蓝型油菜AnArCnCo,避免了不断地自交或者回交过程,最大程度快速拓宽了甘蓝型油菜的抗旱性遗传背景,为甘蓝型油菜抗旱性育种提供了优良的种质资源。植物的抗旱性是由多个基因决定的遗传性状,受基因型和水分胁迫的双重制约[37,38,39]。卢坤等[40]以RNA-Seq技术在甘蓝型油菜叶片中鉴定了部分干旱胁迫应答基因。CHEN等[41]对盐胁迫响应的甘蓝型油菜叶和根进行比较转录组分析,发现分别有582个和438个转录因子在盐胁迫响应中受到差异调节。ZHANG等[42]鉴定了79个在甘蓝型油菜水分胁迫耐受中起作用的候选基因,其中8个基因与干旱耐受性相关。本研究将白菜的抗旱性进行了有效转移,创建了抗旱性新型甘蓝型油菜,可为甘蓝型油菜抗旱性育种提供优质的种质资源,但是对其分子基础仍不清楚,因此还有待更深入的研究。后续研究中,不仅要深入挖掘白菜型油菜产生抗旱性产生的关键基因,还要深入探讨白菜型油菜A亚基因组的导入对于甘蓝型油菜基因组结构、基因组表达特性产生的深远影响。
4 结论
以六倍体材料为桥梁,与抗旱的白菜型油菜杂交,创制、筛选获得了具有优良抗旱性的新型甘蓝型油菜,可为甘蓝型油菜抗旱育种提供优良的基因资源。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
DOI:10.1126/science.1253435URLPMID:25146293 [本文引用: 2]

Oilseed rape (Brassica napus L.) was formed ~7500 years ago by hybridization between B. rapa and B. oleracea, followed by chromosome doubling, a process known as allopolyploidy. Together with more ancient polyploidizations, this conferred an aggregate 72x genome multiplication since the origin of angiosperms and high gene content. We examined the B. napus genome and the consequences of its recent duplication. The constituent An and Cn subgenomes are engaged in subtle structural, functional, and epigenetic cross-talk, with abundant homeologous exchanges. Incipient gene loss and expression divergence have begun. Selection in B. napus oilseed types has accelerated the loss of glucosinolate genes, while preserving expansion of oil biosynthesis genes. These processes provide insights into allopolyploid evolution and its relationship with crop domestication and improvement.
[本文引用: 1]
URLPMID:16483835 [本文引用: 1]
URLPMID:20150489 [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2014.18.005URL [本文引用: 1]
【目的】针对长江流域特别是长江上游近几年春季降水偏少易导致干旱的特点,研究水分胁迫对油菜蕾薹期生理特性及农艺性状的影响,为揭示油菜蕾薹期抗旱机制及抗旱品种选育提供理论依据。【方法】采用盆栽的方式,在遮雨网室对两种抗旱性不同的油菜品种中双10号(抗旱性弱)和94005(抗旱性强)蕾薹期进行干旱胁迫,以各性状的抗旱系数研究油菜相关生理指标及农艺性状的变化,在此基础上筛选出适用于该区域油菜蕾薹期抗旱性鉴定的指标。【结果】随着干旱时间的延长,两种油菜的叶片净光合速率、气孔导度、蒸腾速率、胞间二氧化碳浓度、RWC、叶绿素含量、RuBP羧化酶活性、株高、茎粗、一次分枝数以及单株产量抗旱系数均呈下降趋势,下降幅度与胁迫时间成正相关。其中,气孔导度、蒸腾速率抗旱系数下降幅度与材料抗旱性成正相关,其余指标则与材料抗旱性成负相关。抗旱性弱的材料气孔限制值、水分利用率抗旱系数分别呈现出上升-下降与一直下降的趋势,抗旱性强的材料则均为上升趋势。POD、SOD、CAT活性、可溶性糖以及可溶性蛋白相对值随着干旱胁迫时间的延长先升高后下降,抗旱性强的材料增加幅度高于抗旱性弱的材料;细胞膜透性、丙二醛(MDA)含量和脯氨酸相对值随干旱胁迫时间的延长呈上升趋势。对干旱胁迫下的油菜蕾薹期和复水后的成熟期相关指标之间的关系进行分析表明,干旱胁迫下,产量与净光合速率、RWC、叶绿素含量和一次分枝数抗旱系数呈极显著正相关,与细胞膜透性、MDA含量抗旱系数呈显著负相关。主成分分析将单株产量、净光合速率、胞间二氧化碳浓度、气孔限制值、水分利用率、RWC、叶绿素、RuBP羧化酶、SOD、POD、CAT、细胞膜透性、MDA、可溶性糖、株高、茎粗及一次分枝数划分为第一主成分;将叶片气孔导度、蒸腾速率、脯氨酸、可溶性蛋白划分为第二主成分。【结论】第一主成分的各指标(单株产量、净光合速率、胞间二氧化碳浓度、气孔限制值、水分利用率、RWC、叶绿素、RuBP羧化酶、SOD、POD、CAT、细胞膜透性、MDA、可溶性糖、株高、茎粗及一次分枝数)变化与品种抗旱性密切相关,在油菜抗旱品种选育时可作为油菜蕾薹期抗旱性鉴定的主要指标,第二主成分的各指标(叶片气孔导度、蒸腾速率、脯氨酸、可溶性蛋白)则是次要鉴选指标。
DOI:10.3864/j.issn.0578-1752.2014.18.005URL [本文引用: 1]
【目的】针对长江流域特别是长江上游近几年春季降水偏少易导致干旱的特点,研究水分胁迫对油菜蕾薹期生理特性及农艺性状的影响,为揭示油菜蕾薹期抗旱机制及抗旱品种选育提供理论依据。【方法】采用盆栽的方式,在遮雨网室对两种抗旱性不同的油菜品种中双10号(抗旱性弱)和94005(抗旱性强)蕾薹期进行干旱胁迫,以各性状的抗旱系数研究油菜相关生理指标及农艺性状的变化,在此基础上筛选出适用于该区域油菜蕾薹期抗旱性鉴定的指标。【结果】随着干旱时间的延长,两种油菜的叶片净光合速率、气孔导度、蒸腾速率、胞间二氧化碳浓度、RWC、叶绿素含量、RuBP羧化酶活性、株高、茎粗、一次分枝数以及单株产量抗旱系数均呈下降趋势,下降幅度与胁迫时间成正相关。其中,气孔导度、蒸腾速率抗旱系数下降幅度与材料抗旱性成正相关,其余指标则与材料抗旱性成负相关。抗旱性弱的材料气孔限制值、水分利用率抗旱系数分别呈现出上升-下降与一直下降的趋势,抗旱性强的材料则均为上升趋势。POD、SOD、CAT活性、可溶性糖以及可溶性蛋白相对值随着干旱胁迫时间的延长先升高后下降,抗旱性强的材料增加幅度高于抗旱性弱的材料;细胞膜透性、丙二醛(MDA)含量和脯氨酸相对值随干旱胁迫时间的延长呈上升趋势。对干旱胁迫下的油菜蕾薹期和复水后的成熟期相关指标之间的关系进行分析表明,干旱胁迫下,产量与净光合速率、RWC、叶绿素含量和一次分枝数抗旱系数呈极显著正相关,与细胞膜透性、MDA含量抗旱系数呈显著负相关。主成分分析将单株产量、净光合速率、胞间二氧化碳浓度、气孔限制值、水分利用率、RWC、叶绿素、RuBP羧化酶、SOD、POD、CAT、细胞膜透性、MDA、可溶性糖、株高、茎粗及一次分枝数划分为第一主成分;将叶片气孔导度、蒸腾速率、脯氨酸、可溶性蛋白划分为第二主成分。【结论】第一主成分的各指标(单株产量、净光合速率、胞间二氧化碳浓度、气孔限制值、水分利用率、RWC、叶绿素、RuBP羧化酶、SOD、POD、CAT、细胞膜透性、MDA、可溶性糖、株高、茎粗及一次分枝数)变化与品种抗旱性密切相关,在油菜抗旱品种选育时可作为油菜蕾薹期抗旱性鉴定的主要指标,第二主成分的各指标(叶片气孔导度、蒸腾速率、脯氨酸、可溶性蛋白)则是次要鉴选指标。
URLPMID:24169668 [本文引用: 1]
URLPMID:20350303
URLPMID:21102622 [本文引用: 1]
URLPMID:21847624 [本文引用: 1]
URLPMID:16604336 [本文引用: 2]
[本文引用: 1]
DOI:10.1007/s00122-011-1765-7URL [本文引用: 1]
DOI:10.1007/s10681-011-0533-8URL [本文引用: 1]
Although strong intersubgenomic heterosis for seed production has been observed between "natural'' domesticated Brassica napus (rapeseed, AACC) and a new type of rapeseed into which subgenomic components of Brassica rapa (AA) have been introgressed, the molecular genetic mechanism of this intersubgenomic heterosis is not understood. In this study, a recombinant inbred line population of new type rapeseed derived from a cross between B. napus and B. rapa, together with a population from a backcross with the parental line of B. napus, was used to identify single-locus quantitative trait locus (QTL) and interacting QTL pairs for yield and nine yield-related traits. More than half of single-locus QTLs and interacting QTL pairs detected were involved with the novel alleles induced by the introgression of B. rapa. The alleles directly from B. rapa A genome played a secondary role in contributing to intersubgenomic heterosis. Allelic and nonallelic interactions of both novel alleles generated by B. rapa introgression and the alleles directly from B. rapa A genome contributed to the intersubgenomic heterosis between "natural'' domesticated rapeseed and new type rapeseed into which B. rapa had been introgressed. Six loci for fixed heterosis were identified and their possible applications are also discussed.
URLPMID:28703467 [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1007/s00122-005-1967-yURLPMID:15806345 [本文引用: 2]
Amplified fragment length polymorphism (AFLP) markers were employed to assess the genetic diversity amongst two large collections of Brassica rapa accessions. Collection A consisted of 161 B. rapa accessions representing different morphotypes among the cultivated B. rapa, including traditional and modern cultivars and breeding materials from geographical locations from all over the world and two Brassica napus accessions. Collection B consisted of 96 accessions, representing mainly leafy vegetable types cultivated in China. On the basis of the AFLP data obtained, we constructed phenetic trees using MEGA 2.1: software. The level of polymorphism was very high, and it was evident that the amount of genetic variation present within the groups was often comparable to the variation between the different cultivar groups. Cluster analysis revealed groups, often with low bootstrap values, which coincided with cultivar groups. The most interesting information revealed by the phenetic trees was that different morphotypes are often more related to other morphotypes from the same region (East Asia vs. Europe) than to similar morphotypes from different regions, suggesting either an independent origin and or a long and separate domestication and breeding history in both regions.
URLPMID:24714366 [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/jac.12107URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1007/s00122-015-2459-3URLPMID:25628163 [本文引用: 3]
KEY MESSAGE: Sclerotinia resistance was transferred into rapeseed from a wild relative of Brassica oleracea (B. incana) using hexaploids derived from crosses between B. incana and rapeseed as a bridge. A high level of resistance against Sclerotinia sclerotiorum has been documented in wild Brassica oleracea, but not in cultivated rapeseed (Brassica napus). To transfer sclerotinia resistance from a wild relative into rapeseed, a strategy was proposed using hexaploids (AACCCC) derived from crosses between the wild B. oleracea-related B. incana genotype 'C01' and the Chinese rapeseed variety 'Zhongshuang 9' as a bridge. Progenies (BC1F1) generated by backcrossing the hexaploid to 'Zhongshuang 9' could be generated with a high crossability (average 18.3 seeds per pod). Seventy-three individuals in BC1F1 were firstly screened for resistance with five molecular markers linked to the major resistance QTL on chromosome C09 in 'C01', and 11 individuals harboring resistance loci were selected to develop vegetative clones. Of these, five exhibited significantly higher resistance than 'Zhongshuang 9' and the most resistant individual was chosen to develop the BC1F2 progeny. Finally, five individual genotypes with nearly twofold higher resistance than 'Zhongshuang 9' were found among 100 BC1F2 individuals by using marker-assisted selection and resistance evaluation. Hereof, one rapeseed-type individual with 38 chromosomes and good self-fertility (15.0 +/- 3.56 seeds/pod) was identified. Our results indicate that the proposed strategy is effective for transferring sclerotinia resistance from a wild relative of B. oleracea into rapeseed.
DOI:10.3864/j.issn.0578-1752.2017.01.011URL [本文引用: 4]
【目的】利用亲本种遗传资源是改良甘蓝型油菜的重要手段。以甘蓝型油菜与菌核病抗性甘蓝杂交合成的六倍体为桥梁,与大量的白菜型油菜杂交,合成杂种,探索改良甘蓝型油菜菌核病抗性的策略。【方法】采用菌核病抗病甘蓝(C01)与甘蓝型油菜(中双9号)杂交合成六倍体,通过分析六倍体的育性、菌病抗性和减数分裂行为来分析其作为桥梁材料转移菌核病抗性的可能性;将六倍体与110份白菜型油菜杂交,通过考察杂种发育和可交配性来分析六倍体与白菜型油菜杂交的可行性;通过鉴定杂种的苗期表型特征、自交结实率及离体茎秆的菌核病抗性来分析杂种在改良甘蓝型油菜遗传背景上的利用潜力。【结果】该六倍体的花粉育性为90.6%—92.7%,自交结实率为3—7粒/角果;菌核病抗性显著高于对照品种(中双9号);处于减数分裂后期I的花粉母细胞中,68.80%(86/125)的染色体分离比为28﹕28。110份春性、冬性和半冬性的白菜型油菜与六倍体杂交,授粉15 d后的胚珠发育正常,并且都能收获成熟种子,平均可交配性为(4.25±3.91)粒/角果。尽管不同基因型之间的可交配性存在显著差异,但是不同生态型的白菜型油菜与六倍体杂交的可交配性无显著差异(半冬性:(4.35±3.77)粒/角果,春性:(4.34±4.51)粒/角果,冬性:(4.01±3.43)粒/角果;P=0.44)。六倍体作为母本或者父本与白菜型油菜杂交都能结籽,而且没有显著性差异(六倍体为母本:平均结实率为4.27粒/角果;六倍体为父本:平均结实率为3.95粒/角果;P=0.69)。六倍体与白菜型油菜杂交创建的杂种,苗期形态似甘蓝型油菜,但是表型变异丰富;杂种都能自交结籽,平均自交结实率为(7.72±4.45)粒/角果;来自不同生态型的白菜型油菜与六倍体合成的杂种自交结实率无显著性差异(冬性白菜型油菜合成的杂种平均自交结实率:(8.07±3.43)粒/角果,半冬性:(7.88±4.64)粒/角果,春性:(6.41±3.00)粒/角果,P=0.95)。经过两年的离体茎秆菌核病抗性鉴定,6份杂种的菌核病发病程度两年均显著低于中双9号(P<0.05)。【结论】以六倍体为桥梁能有效地将甘蓝型油菜亲本种的优良性状导入到甘蓝型油菜中。
DOI:10.3864/j.issn.0578-1752.2017.01.011URL [本文引用: 4]
【目的】利用亲本种遗传资源是改良甘蓝型油菜的重要手段。以甘蓝型油菜与菌核病抗性甘蓝杂交合成的六倍体为桥梁,与大量的白菜型油菜杂交,合成杂种,探索改良甘蓝型油菜菌核病抗性的策略。【方法】采用菌核病抗病甘蓝(C01)与甘蓝型油菜(中双9号)杂交合成六倍体,通过分析六倍体的育性、菌病抗性和减数分裂行为来分析其作为桥梁材料转移菌核病抗性的可能性;将六倍体与110份白菜型油菜杂交,通过考察杂种发育和可交配性来分析六倍体与白菜型油菜杂交的可行性;通过鉴定杂种的苗期表型特征、自交结实率及离体茎秆的菌核病抗性来分析杂种在改良甘蓝型油菜遗传背景上的利用潜力。【结果】该六倍体的花粉育性为90.6%—92.7%,自交结实率为3—7粒/角果;菌核病抗性显著高于对照品种(中双9号);处于减数分裂后期I的花粉母细胞中,68.80%(86/125)的染色体分离比为28﹕28。110份春性、冬性和半冬性的白菜型油菜与六倍体杂交,授粉15 d后的胚珠发育正常,并且都能收获成熟种子,平均可交配性为(4.25±3.91)粒/角果。尽管不同基因型之间的可交配性存在显著差异,但是不同生态型的白菜型油菜与六倍体杂交的可交配性无显著差异(半冬性:(4.35±3.77)粒/角果,春性:(4.34±4.51)粒/角果,冬性:(4.01±3.43)粒/角果;P=0.44)。六倍体作为母本或者父本与白菜型油菜杂交都能结籽,而且没有显著性差异(六倍体为母本:平均结实率为4.27粒/角果;六倍体为父本:平均结实率为3.95粒/角果;P=0.69)。六倍体与白菜型油菜杂交创建的杂种,苗期形态似甘蓝型油菜,但是表型变异丰富;杂种都能自交结籽,平均自交结实率为(7.72±4.45)粒/角果;来自不同生态型的白菜型油菜与六倍体合成的杂种自交结实率无显著性差异(冬性白菜型油菜合成的杂种平均自交结实率:(8.07±3.43)粒/角果,半冬性:(7.88±4.64)粒/角果,春性:(6.41±3.00)粒/角果,P=0.95)。经过两年的离体茎秆菌核病抗性鉴定,6份杂种的菌核病发病程度两年均显著低于中双9号(P<0.05)。【结论】以六倍体为桥梁能有效地将甘蓝型油菜亲本种的优良性状导入到甘蓝型油菜中。
[本文引用: 1]
URLPMID:20394281 [本文引用: 2]
URLPMID:20394281 [本文引用: 2]
DOI:10.1007/s11032-014-0153-9URL [本文引用: 1]
DOI:10.1007/BF00303970URL [本文引用: 1]
DOI:10.1007/s10722-013-9978-zURL [本文引用: 1]
Sclerotinia stem rot is one of the most serious diseases in rapeseed (Brassica napus) due to the lack of resistance sources. A high level of resistance was reported in Brassica oleracea cytodeme, one of parental species of rapeseed. In this study, a panel of 55 resynthesized lines of B. napus (RS lines) derived from seven wild and two cultivated types of B. oleracea was evaluated for Sclerotinia resistance over 2 years. Relative to 'Zhongyou 821', a cultivar of B. napus with partial resistance against S. sclerotiorum, RS lines exhibited stronger stem resistance. Although the resistant level of RS lines was lower than that of corresponding parental B. oleracea, a moderate correlation between resistance of RS line and corresponding parental B. oleracea type was found both for leaf (r = 0.74, P = 0.02) and stem (r = 0.69, P = 0.04). Our data suggests that the RS lines are important resources to improve Sclerotinia resistance of current rapeseed. A breeding strategy is discussed to enhance the Sclerotinia resistance of rapeseed by using B. oleracea.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLPMID:25336153 [本文引用: 1]
DOI:10.1093/jxb/erl164URLPMID:17075077 [本文引用: 1]
Plants respond to survive under water-deficit conditions via a series of physiological, cellular, and molecular processes culminating in stress tolerance. Many drought-inducible genes with various functions have been identified by molecular and genomic analyses in Arabidopsis, rice, and other plants, including a number of transcription factors that regulate stress-inducible gene expression. The products of stress-inducible genes function both in the initial stress response and in establishing plant stress tolerance. In this short review, recent progress resulting from analysis of gene expression during the drought-stress response in plants as well as in elucidating the functions of genes implicated in the stress response and/or stress tolerance are summarized. A description is also provided of how various genes involved in stress tolerance were applied in genetic engineering of dehydration stress tolerance in transgenic Arabidopsis plants.
URLPMID:12972040 [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2015.04.02URL [本文引用: 1]
【目的】利用RNA Sequencing(RNA-Seq)技术比较2种不同生长条件下甘蓝型油菜苗期叶片转录组,鉴定油菜叶片干旱胁迫应答相关基因,从转录组水平揭示油菜适应干旱胁迫环境的分子机制。【方法】提取正常生长(ZY)和自然失水处理(ZY8D)的六叶期甘蓝型油菜中油821的叶片总RNA,以Illumina Hiseq 2000平台进行RNA-Seq分析。利用NGSQCTookit v2.3.3去除低质量和包含模糊碱基的reads。以甘蓝型油菜亲本物种白菜染色体v1.5和甘蓝Scaffold v1.0为参考序列,采用TopHat2-Cufflinks-Cuffmerge-Cuffdiff标准流程进行差异表达基因(differential expressed genes,DEGs)筛选。对上调和下调DEGs分别采用Cytoscape v3.1.0中的BiNGO和KOBAS2.0进行基因本体(gene ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)代谢途径富集分析。选择上调和下调DEGs各3个,以实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)验证RNA-Seq结果的可靠性。【结果】过滤低质量reads后,ZY和ZY8D分别保留了26 192 312和28 378 899对高质量reads用于DEGs筛选,其中86.6%和85.8%的reads能准确比对到参考序列上,说明RNA-Seq结果和参考序列可靠。DEGs鉴定结果表明3 657个基因受干旱胁迫诱导差异表达,其中上调表达基因1 431个,下调表达基因2 226个。GO富集分析发现上调表达基因主要与非生物胁迫响应和化学刺激响应相关,其中,参与水分胁迫响应和脱落酸(abscisic acid,ABA)刺激响应的基因分别有127和141个,而下调表达基因与植物病原菌防御、蛋白激酶活性和水杨酸(salicylic acid,SA)刺激相关。KEGG富集分析表明上调表达基因主要富集于苯丙烷和类胡萝卜素的生物合成及淀粉与蔗糖代谢途径,而下调表达基因主要富集于植物-病原菌互作和植物激素ABA、SA和茉莉酸(jasmonic acid,JA)信号转导途径。qRT-PCR检测6个DEGs的表达模式与RNA-Seq分析结果一致,证实了RNA-Seq结果的可靠性。【结论】RNA-Seq分析鉴定出3 657个甘蓝型油菜叶片干旱胁迫应答基因。GO和KEGG代谢途径分析明确了差异表达基因富集的分子功能与代谢途径。
DOI:10.3864/j.issn.0578-1752.2015.04.02URL [本文引用: 1]
【目的】利用RNA Sequencing(RNA-Seq)技术比较2种不同生长条件下甘蓝型油菜苗期叶片转录组,鉴定油菜叶片干旱胁迫应答相关基因,从转录组水平揭示油菜适应干旱胁迫环境的分子机制。【方法】提取正常生长(ZY)和自然失水处理(ZY8D)的六叶期甘蓝型油菜中油821的叶片总RNA,以Illumina Hiseq 2000平台进行RNA-Seq分析。利用NGSQCTookit v2.3.3去除低质量和包含模糊碱基的reads。以甘蓝型油菜亲本物种白菜染色体v1.5和甘蓝Scaffold v1.0为参考序列,采用TopHat2-Cufflinks-Cuffmerge-Cuffdiff标准流程进行差异表达基因(differential expressed genes,DEGs)筛选。对上调和下调DEGs分别采用Cytoscape v3.1.0中的BiNGO和KOBAS2.0进行基因本体(gene ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)代谢途径富集分析。选择上调和下调DEGs各3个,以实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)验证RNA-Seq结果的可靠性。【结果】过滤低质量reads后,ZY和ZY8D分别保留了26 192 312和28 378 899对高质量reads用于DEGs筛选,其中86.6%和85.8%的reads能准确比对到参考序列上,说明RNA-Seq结果和参考序列可靠。DEGs鉴定结果表明3 657个基因受干旱胁迫诱导差异表达,其中上调表达基因1 431个,下调表达基因2 226个。GO富集分析发现上调表达基因主要与非生物胁迫响应和化学刺激响应相关,其中,参与水分胁迫响应和脱落酸(abscisic acid,ABA)刺激响应的基因分别有127和141个,而下调表达基因与植物病原菌防御、蛋白激酶活性和水杨酸(salicylic acid,SA)刺激相关。KEGG富集分析表明上调表达基因主要富集于苯丙烷和类胡萝卜素的生物合成及淀粉与蔗糖代谢途径,而下调表达基因主要富集于植物-病原菌互作和植物激素ABA、SA和茉莉酸(jasmonic acid,JA)信号转导途径。qRT-PCR检测6个DEGs的表达模式与RNA-Seq分析结果一致,证实了RNA-Seq结果的可靠性。【结论】RNA-Seq分析鉴定出3 657个甘蓝型油菜叶片干旱胁迫应答基因。GO和KEGG代谢途径分析明确了差异表达基因富集的分子功能与代谢途径。
DOI:10.1093/abbs/gmp113URLPMID:20119627 [本文引用: 1]
High salinity and drought are the major abiotic stresses that adversely affect plant growth and agricultural productivity. To investigate genes that are involved in response to abiotic stresses in Brassica napus, a comprehensive survey of genes induced by high-salinity and drought stresses was done by macroarray analysis. In total, 536 clones were identified to be putative high-salinity-or drought-responsive genes. Among them, 172 and 288 clones are detected to be putative high-salinity- and drought-inducible genes, whereas 141 and 189 are candidates for high-salinity- and drought-suppressed genes, respectively. The functional classification of these genes are indicated that belonged to gene families encoding metabolic enzymes, regulatory factors, components of signal transduction, hormone responses, some abiotic stresses-related proteins, and other processes related to growth and development of B. napus. From the upregulated candidate genes, some interested genes were further demonstrated to be high-salinity- or/and drought-induced expression by real-time quantitative RT-PCR analysis. The experimental results also revealed that some genes may function in abscisic acid-dependent signaling pathway related to drought or salinity stress. Collectively, the data presented in this study will facilitate the understanding of molecular mechanism of B. napus in response to high-salinity and drought stresses, and also provide us the basis of effective genetic engineering strategies for improving stress tolerance of B. napus.
URLPMID:26640475 [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}