ABA Metabolism and Signaling and Their Molecular Mechanism Regulating Seed Dormancy and Germination
SONG SongQuan1,4, LIU Jun2, XU HengHeng2, LIU Xu,3, HUANG Hui41 Institute of Botany, Chinese Academy of Sciences, Beijing 100093 2 Agro-Biological Gene Research Center, Guangdong Academy of Agricultural Sciences, Guangzhou 510640 3 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081 4 Key Laboratory of Research and Utilization of Ethnomedicinal Plant Resources of Hunan Province, Huaihua University/College of Biological and Food Engineering, Huaihua 418008, Hunan
Abstract Seed dormancy is an adaptive characteristic to environmental changes acquired by many plants during long-term phylogenetic development, and is an effective way regulating the optimal spatiotemporal distribution of seed germination and seedling formation, and is also a selective strategy for the successful reproduction and propagation in species. Phytohormonal regulation of seed dormancy and germination may be a highly conserved mechanism, of which abscisic acid (ABA) plays a master role in dormancy release and germination, and gibberellin (GA) functions as stimulating seed germination after dormancy is released. The role of ABA in seed dormancy and germination is mainly regulated by its metabolism (biosynthesis and catabolism) and signaling pathways. Therefore, in this paper, we mainly summarize the research progresses of ABA metabolism and signaling, the effects of ABA on seed development, dormancy and germination as well as the relationships between DOG1 (DELAY OF GERMINATION1, a specific gene involved in seed dormancy) and ABA signaling components. The researches showed that C40 epoxycarotenoid is a precursor, and zeaxanthin epoxidase and 9-cis-epoxycarotenoid dioxygenase are the principal regulatory enzymes in ABA biosynthesis. The ABA catabolism includes hydroxylation and conjugation with glucose. The hydroxylation of ABA at C-8' position is catalyzed by the CYP707A, which is an important step for ABA catabolism. In the core ABA signaling pathway, ABA binds to PYR/PYL/RCAR receptors and triggers a conformational change that allows receptor-ABA complex to bind to and inhibit type 2C protein phosphatase (PP2C) activity, which results in de-repression and activation of kinases such as sucrose non-fermenting1-related protein kinase 2 (SnRK2). These kinases then phosphorylate and activate transcription factors (TF), which bind to the target promoters and induce the expression of ABA response gene downstream. ABA accumulates in seeds during mid- and late-maturation stages, and ABA synthesized in zygotic tissues induces primary dormancy and promotes seed maturation. ABA content accumulated during development and preserved in dry seeds declines at the early stage of seed imbibition. ABA is a positive regulator of seed dormancy induction and maintenance, and is a negative regulator of seed germination. DOG1 expresses and functions during seed maturation, and its expression is regulated by alternative splicing and alternative polyadenylation. Antisense DOG1 is a repressor of seed dormancy, which negatively regulates DOG1 expression and seed dormancy by causing transcriptional interference and affecting transcription extension. Seed dormancy and germination are regulated not only by core ABA signaling pathway, but also by DOG1-AHG1 (ABA HYPERSENSITIVE GERMINATION1)/AHG3 pathway. DOG1 can bind to AHG1/AHG3 and cause seed dormancy by sequestrating those negative regulators of ABA signaling and increasing ABA sensitivity in seeds. Finally, we propose some scientific issues required for investigation further in the future. How do ABA 8'-hydroxylase, ABA glucosyltransferase and β-glucosidase and their genes respond to developmental and environmental changes to maintain the normal ABA levels in ABA catabolism? How do the important regulators in ABA physiology such as Ca 2+ or reactive oxygen species influence the core ABA signaling pathway? Which pathway is preferentially responded by PP2C, a downstream overlapping component of core ABA signaling pathway and DOG1-AHG1/AHG3 pathway, when it integrates physiological conditions or environmental signals, and how are these two pathways coordinated, and what new target components does PP2C have? This paper will provide a basis to further investigate the molecular mechanism regulating seed dormancy and germination by ABA. Keywords:abscisic acid;dormancy;dormancy gene DOG1;germination;metabolism;signaling
PDF (1356KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 宋松泉, 刘军, 徐恒恒, 刘旭, 黄荟. 脱落酸代谢与信号传递及其调控种子休眠与萌发的分子机制[J]. 中国农业科学, 2020, 53(5): 857-873 doi:10.3864/j.issn.0578-1752.2020.05.001 SONG SongQuan, LIU Jun, XU HengHeng, LIU Xu, HUANG Hui. ABA Metabolism and Signaling and Their Molecular Mechanism Regulating Seed Dormancy and Germination[J]. Scientia Acricultura Sinica, 2020, 53(5): 857-873 doi:10.3864/j.issn.0578-1752.2020.05.001
ABA是调控植物许多发育过程包括种子休眠、萌发和幼苗生长,以及控制许多非生物胁迫反应的关键因子[12,13,14]。研究结果证明,ABA在许多物种的种子休眠中起重要作用:外源ABA可以延迟或抑制种子和胚的萌发;在未成熟种子中,内源ABA维持种胚处于发育而不是萌芽过程;收获前的萌发也与种子中ABA含量较低有关;由ABA生物合成突变、转基因修饰或者化学抑制所引起的ABA缺乏的种子是非休眠的;ABA生物合成的化学抑制也引起一些休眠种子的萌发;ABA生物合成基因的过表达也会抑制和延迟种子萌发;在吸胀的最初几个小时,非休眠种子中的ABA含量比休眠种子下降更多[15,16,17]。ABA在种子休眠与萌发中的作用主要受ABA代谢(包括生物合成和分解代谢)和信号传递途径的调控[12-13,17-19],环境因子对种子休眠和萌发的影响也是通过ABA和GA起作用[20]。然而,许多证据表明GA主要是在种子休眠被解除后起促进萌发的作用,而不是参与解除种子休眠[17,20-21]。本文主要综述ABA代谢与信号传递的研究进展,ABA在种子发育、休眠与萌发中的作用,以及种子休眠基因DOG1(DELAY OF GERMINATION1)和ABA信号组分的关系;此外,我们提出了该领域需要进一步研究的问题,试图为深入研究ABA调控种子休眠与萌发的分子机理提供新的参考。
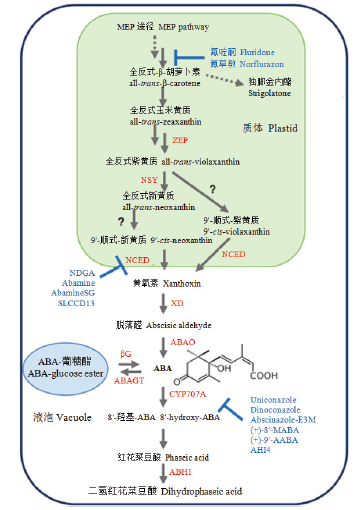
ABA前体是由甲基赤藓糖醇磷酸(MEP)途径合成的。酶用红色表示。ZEP:玉米黄质环氧化酶;NSY:新黄质合酶;NCED:9-顺式-环氧类胡萝卜素双加氧酶;XD:黄氧素脱氢酶;ABAO:脱落醛氧化酶;CYP707A:ABA 8'-羟化酶;ABH1:红花菜豆酸还原酶1;ABAGT:ABA葡糖基转移酶;βG:β-葡糖苷酶。酶的抑制剂用蓝色表示。(+)-9'-AABA:(+)-9'-乙炔-ABA;AHI4:ABA 8'-羟化酶抑制剂4;(+)-8'-MABA:(+)-8'-次甲基-ABA;NDGA:去甲二氢愈创木酸;SLCCD13:类倍半萜类胡萝卜素裂解双加氧酶抑制剂13 Fig. 1ABA biosynthetic and catabolic pathways (Modified from DEJONGHE et al.[13])
ABA precursor is synthesized from the methylerythritol phosphate (MEP) pathway. Enzymes are shown in red colour. ZEP: Zeaxanthin epoxidase; NSY: Neoxanthin synthase; NCED: 9-cis-epoxycarotenoid dioxygenase; XD: Xanthoxin dehydrogenase; ABAO: Abscisic aldehyde oxidase; CYP707A: ABA 8'-hydroxylase; ABH1: Phaseic acid reductase 1; ABAGT: ABA glucosyltransferase; βG: β-glucosidase. Enzyme inhibitors are shown in blue colour. (+)-9'-AABA: (+)-9'-acetylene-ABA; AHI4: ABA-8'-hydroxylase inhibitor 4; (+)-8'-MABA: (+)-8'-methylidyne-ABA; NDGA: Nordihydroguaiaretic acid; SLCCD13: Sesquiterpene-like carotenoid cleavage dioxygenase inhibitor 13
核心ABA信号传递组分主要由PYR/PYL/RCAR(pyrabactin resistance 1/pyrabactin resistance 1-like/ regulatory components of ABA receptor)蛋白、A组2C类蛋白磷酸酶(group A type 2C protein phosphatase,PP2C)、亚类Ⅲ蔗糖非发酵-1-相关蛋白激酶2(subclass Ⅲ sucrose nonfermenting-1-related protein kinase2,SnRK2)和ABF(ABA-responsive element (ABRE)- binding factor)/AREB(ABRE-binding protein)转录因子组成(图2和图3)[12-13,19,47-49]。ABA通过与PYR/PYL/ RCAR蛋白中高度保守的氨基酸进行直接的和水介导的接触,被结合进疏水的配体结合的ABA受体(结合)口袋中。结合口袋(binding pocket)含有类似于一只折叠的手的7个β折叠,以及1个大的和2个较小的α螺旋[50,51]。ABA的结合促进了包含β3和β4之间的一个门环(gate-loop)的构象变化,这种构象变化关闭结合口袋,形成与ABA的接触。除了PYR/PYL/RCAR12和PYR/PYL/RCAR13分别含有序列-SDLPA-和-SGFPA-外,在所有的PYR/PYL/ RCAR蛋白的门环中都含有序列-SGLPA-[13]。β5和β6含有不变的序列-HRL-,它们之间的第二个“门闩(latch)”环也发生构象改变;这种改变使受体-配体复合物对接和抑制PP2C。PP2C含有一个高度保守的、定位于A组专一识别环中的色氨酸残基,该残基能插入到由门环关闭所产生的小口袋中,并与ABA的酮基产生水介导的接触。这个水分子位于ABA、门的脯氨酸(-SGLPA-)、门闩的精氨酸(-HRL-)和PP2C的色氨酸锁之间的H-键网络中心(图2)[12,50,52]。
在ABA缺乏时,PYR/PYL(pyrabactin resistance 1/pyrabactin resistance 1-like)蛋白具有一个开放的门和门闩环的构型(分别为红色和绿色),它们位于ABA结合口袋的侧面。ABA的结合诱导门和门闩的关闭,依次产生相互作用的表面,使2C类蛋白磷酸酶(PP2C)对接到结合ABA的受体上。门中的一个保守的脯氨酸(对应于PYR1中的脯氨酸88残基,用蓝色表示)在对接位点与PP2C形成直接的接触,这解释了用PYR1P88观察到的PP2C结合的缺陷 Fig. 2ABA-induced changes in receptor conformation (From CUTLER et al.[12])
In the absence of ABA, PYR/PYL (pyrabactin resistance 1/pyrabactin resistance 1-like) proteins possess an open conformation of the gate and latch loops (red and green, respectively) that flank the ABA-binding pocket. Binding of ABA induces closure of the gate and latch, which in turn creates the interaction surface that recruits docking of type 2C protein phosphatases (PP2C) onto the ABA-bound receptors. A conserved proline in the gate (which corresponds to the residue to proline 88 in PYR1 and is shown in blue) forms a direct contact with the PP2C at the docking site, which explains the PP2C-binding defect observed with PYR1P88
在ABA感受和信号传递途径中(左),ABA受体(PYR/PYL/RCAR)与ABA不敏感1(ABI1)亚家族2C类蛋白磷酸酶(PP2C)包括ABI1、ABI2、ABA过敏感1(HAB1)和HAB2结合,并使PP2C失活,从而导致激酶例如蔗糖非发酵-1相关的蛋白激酶2(SnRK2)的去抑制和活化。这些激酶然后磷酸化和活化转录因子(TF),TF与靶启动子(Pro)结合,诱导下游的ABA反应基因。对于种子休眠的调节(右),DOG1与ABA过敏感萌发1(AHG1)和AHG3结合,PP2C主要在种子中起作用。DOG1被认为是通过束缚这些ABA信号传递的负调控因子和增加种子对ABA的敏感性而引起种子休眠 Fig. 3ABA signalling pathway and emerging model of seed dormancy regulated by DOG1 (Modified from NONOGAKI [19])
In the ABA perception and signaling pathway (left), ABA receptors (PYR/PYL/RCARs) bind to and inactivate the ABA INSENSITIVE1 (ABI1) subfamily protein phosphatases 2C (PP2Cs), including ABI1, ABI2, HYPERSENSITIVE TO ABA1 (HAB1) and HAB2, which results in de-repression and activation of kinases, such as sucrose nonfermenting1-related protein kinase 2 (SnRK2). These kinases then phosphorylate and activate transcription factors (TF), which bind to the target promoters (Pro), to induce ABA-responsive genes downstream. For seed dormancy regulation (right), DOG1 binds to ABA HYPERSENSITIVE GERMINATION1 (AHG1) and AHG3, PP2Cs primarily functioning in seeds. The DOG1 is thought to cause seed dormancy by sequestrating these negative regulators of ABA signaling and increasing ABA sensitivity in seeds
DOG1在种子成熟过程中表达,其表达受可变剪接(alternative splicing)[95,97]和可变多腺苷酸化(alternative polyadenylation)的调控[91,98]。DOG1的几个剪接变体(splicing variant)[95]可产生5种转录物变异体(transcript variant,α、β、γ、δ、ε)和3种不同的蛋白(图4-A)[97];其中DOG1-ε不是一个真正意义上的剪接变体,但是拟南芥发育种子中的主要形式[97];可变剪接可以构成不同功能的蛋白,包括它们的亚细胞定位和增加的休眠潜能。这3种蛋白(α、(β、γ、ε)、δ)都能被转运到细胞核[97],因此,可以认为DOG1是作为一种调控蛋白以同源二聚体的形式起作用[96,97]。过表达分析结果表明,这3种同源异构体可以诱导种子休眠,且共表达时更为稳定。因此,异源二聚体的形成不能解释DOG1蛋白具有更好的稳定性。对于DOG1的稳定性,同源异构体共表达的正作用机制尚不够清楚[91]。当酵母剪接体组分19号复合物相关蛋白1(nineteen complex- related protein 1)的拟南芥直系同源物(AtNTR1)被突变时,它引起DOG1中内含子保留和外显子跳跃(intron retention and exon skipping)的主要缺陷。这种DOG1剪接的错误能调控种子减少休眠,但这种表型不会由可变剪接本身引起,而可能是这种突变体中DOG1表达水平降低的结果[99]。研究已经发现,转录延伸的效率(transcription elongation efficiency)对于DOG1的表达和种子休眠是一种重要的因子[7,100-102],认为AtNTR1是在剪接位点控制着RNA聚合酶Ⅱ(PolⅡ)的作用,并可作为转录延伸的校正点(checkpoint)[99]。
A:DOG1的结构。顶部:具有外显子(E1、E2、E3)和内含子(I1、I2)的DOG1基因组DNA。可变剪接区域用粉红色和橙色作标记。表明dog1突变(dog1-3、dog1-4和dog1-5中的T-DNA,以及dog1-1中的单个碱基缺失(-C))的大致位置。中部:可变的DOG1转录物(α、β、γ、δ、ε)和相应的蛋白。注意DOG1-ε不是一个真正意义上的可变剪接产物。底部:可变多腺苷酸化的短DOG1(shDOG1,与DOG1-ε相同)和长DOG1(lgDOG1,包括DOG1-α、DOG1-β、DOG1-γ和DOG1-δ)转录物。转录起始(TSS)和终止(TTS)位点被表明。反义DOG1(asDOG1)的大致位置和方向用蓝色箭头标明。B:AsDOG1功能的可能机制。相对稳定的asDOG1 RNA可能以一种序列专一的方式或者通过它的二级结构作为一种调节RNA起作用,用于RNA介导的染色质重塑(右图,反式调节)。然而,等位基因专一的asDOG1的表达已经表明asDOG1在顺式调节中起作用(左图)。转录本身的“行为”而不是转录产物(RNA)发挥asDOG1的表达对DOG1表达和休眠的负面作用。反义表达可能引起转录干扰和影响转录延伸,这对DOG1表达和种子休眠是重要的;而转录介导的染色质重塑也是可能的。AS:可变剪接;APA:可变多腺苷酸化;Dist:远端;Prox:近段;Prot:蛋白;Tran:转录物 Fig. 4Regulation of DOG1 expression and function (From NONOGAKI[91])
A: Structures of the DOG1 gene. Top: DOG1 gDNA with exons (E1, E2, E3) and introns (I1, I2). Alternatively spliced regions are highlighted in pink and orange. Approximate positions of the dog1 mutations (T-DNAs in dog1-3, dog1-4, dog1-5 and a single-base deletion [-C] in dog1-1) are also indicated. Middle: Alternative DOG1 transcripts (α, β, γ, δ, ε) and the corresponding proteins. Note that DOG1-ε is not exactly an alternative splicing product. Bottom: Alternatively polyadenylated short DOG1 (shDOG1), which is identical to DOG1-ε and long (lgDOG1) transcripts, which comprises DOG1-α, -β, -γ and -δ. The transcriptional start (TSS) and termination (TTS) sites are indicated. Approximate position and the orientation of antisense DOG1 (asDOG1) are shown as a blue arrow. B: Possible mechanisms of asDOG1 function. Relatively stable asDOG1 RNA could function as a regulatory RNA, in a sequence-specific manner or through its secondary structure, for RNA-mediated chromatin remodeling (right panel, trans regulation). However, allele-specific asDOG1 expression has indicated that asDOG1 functions in cis (left panel). The “act” of transcription itself, rather than its product (RNA), exerts the negative effects of asDOG1 expression to DOG1 expression and dormancy. Antisense expression could cause transcriptional interference and affect transcription elongation, which is known to be important for DOG1 expression and seed dormancy while transcription-mediated chromatin remodeling is also possible. AS: Alternative splicing; APA: Alternative polyadenylation; Dist: Distal; Prox: Proximal; Prot: Protein; Tran: Transcription
SONG SQ. Seed dormancy//The Editorial Board of Agricultural Science for 10000 Selected Problems in Sciences, ed 10000 Selected Problems in Sciences. Beijing: Science Press, 2011: 31-35. (in Chinese) [本文引用: 1]
GUBLERF, MILLER AA, JACOBSEN JV . Dormancy release, ABA and pre-harvest sprouting Current Opinion in Plant Biology, 2005,8:183-187. [本文引用: 2]
NONOGAKIH . Seed dormancy and germination-emerging mechanism and new hypotheses Frontiers in Plant Science, 2014,e5:233. [本文引用: 3]
CORBINEAUF, XIAQ, BAILLYC, EI-MAAROUF-BOUTEAUH . Ethylene, a key factor in the regulation of seed dormancy Frontiers in Plant Science, 2014,5:539. [本文引用: 1]
KUCERAB, COHN MA, LEUBNER-METZGERG . Plant hormone interactions during seed dormancy release and germination Seed Science Research, 2005,15:281-307. [本文引用: 1]
XU HH, LIN, LIU SJ, WANG WQ, WANG WP, ZHANGH, CHENG HY, SONG SQ . Research progress in seed germination and its control Acta Agronomica Sinica, 2014,40:1141-1156. (in Chinese) [本文引用: 1]
CUTLER SR, RODRIGUEZ PL, FINKELSTEIN RR, ABRAMS SR . Abscisic acid: Emergence of a core signaling network Annual Review of Plant Biology, 2010,61:651-679. [本文引用: 13]
DEJONGHEW, OKAMOTOM, CUTLER SR . Small molecule probes of ABA biosynthesis and signaling Plant Cell and Physiology, 2018,59:1490-1499. [本文引用: 7]
VISHWAKARMAK, UPADHYAYN, KUMARN, YADAVG, SINGHJ, MISHRA RK, KUMARV, VERMAR, UPADHYAY RG, PANDEYM, SHARMAS . Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects Frontiers in Plant Science, 2017,8:161. [本文引用: 1]
GIANINETTIA, VERNIERIP . On the role of abscisic acid in seed dormancy of red rice Journal of Experimental Botany, 2007,58:3449-3462. [本文引用: 1]
MILLAR AA, JACOBSEN JV, ROSS JJ, HELLIWELL CA, POOLE AT, SCOFIELDG, REID JB, GUBLERF . Seed dormancy and ABA metabolism in Arabidopsis and barley: The role of ABA 8'-hydroxylase. The Plant Journal, 2006,45:942-954. [本文引用: 8]
NAMBARAE, OKAMOTOM, TATEMATSUK, YANOR, SEOM, KAMIYAY . Abscisic acid and the control of seed dormancy and germination Seed Science Research, 2010,20:55-67. [本文引用: 4]
NAMBARAE, MARION-POLLA . Abscisic acid biosynthesis and catabolism Annual Review of Plant Biology, 2005,56:165-185. [本文引用: 4]
NONOGAKIH . Seed germination and dormancy - The classic story, new puzzles, and evolution Journal of Integrative Plant Biology, 2019,61:541-563. [本文引用: 14]
HOLDSWORTHR, BENTSINKL, SOPPE WJ J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytologist, 2008,179:33-54. [本文引用: 2]
LINKIESA, LEUBNER-METZGERG . Beyond gibberellins and abscisic acid: How ethylene and jasmonates control seed germination Plant Cell Reports, 2012,31:253-270. [本文引用: 1]
NORTH HM, ALMEIDA AD, BOUTINJ-P, FREYA, TOA, BOTRANL, SOTTAB, MARION-POLLA . The Arabidopsis ABA-deficient mutant aba4 demonstrates that the major route for stress-induced ABA accumulation is via neoxanthin isomers. The Plant Journal, 2007,50:810-824. [本文引用: 2]
TAN BC, JOSEPH LM, DENG WT, LIUL, LI QB, CLINEK , McCARTY D R. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. The Plant Journal, 2003,35:44-56. [本文引用: 1]
CHENG WH, ENDOA, ZHOUL, PENNEYJ, CHEN HC, ARROYOA, LEONP, NAMBARAE, ASAMIT, SEOM, KOSHIBAT, SHEENJ . A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. The Plant Cell, 2002,14:2723-2743. [本文引用: 1]
GONZáLEZ-GUZMáM, APOSTOLOVAN, BELLéS JM, BARRERO JM, PIQUERASP, PONCE MR, MICOL JL, SERRANOR, RODRíGUEZP . The short-chain alcohol dehydrogenase ABA2 catalyzes the conversion of xanthoxin to abscisic aldehyde The Plant Cell, 2002,14:1833-1846. [本文引用: 1]
XIONGL, ISHITANIM, LEEH, ZHU JK . The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression. The Plant Cell, 2001,13:2063-2083. [本文引用: 1]
GAMBLE PE, MULLET JE . Inhibition of carotenoid accumulation and abscisic acid biosynthesis in fluridone-treated dark-grown barley European Journal of Biochemistry, 1986,160:117-121. [本文引用: 1]
CREELMAN RA, BELLE, MULLET JE . Involvement of a lipoxygenase-like enzyme in abscisic acid biosynthesis Plant Physiology, 1992,99:1258-1260. [本文引用: 1]
MéRIGOUTP, KéPèSF, PERRET AM, SATIAT-JEUNEMAITREB, MOREAUP . Effects of brefeldin A and nordihydroguaiaretic acid on endomembrane dynamics and lipid synthesis in plant cells FEBS Letters, 2002,518:88-92. [本文引用: 1]
HAN SY, KITAHATAN, SEKIMATAK, SAITOT, KOBAYASHIM, NAKASHIMAK, YAMAGUCHI-SHINOZAKIK, SHINOZAKIK, YOSHIDAS, ASAMIT . A novel inhibitor of 9-cis-epoxycarotenoid dioxygenase in abscisic acid biosynthesis in higher plants. Plant Physiology, 2004,135:1574-1582. [本文引用: 1]
KITAHATAN, HAN SY, NOJIN, SAITOT, KOBAYASHIM, NAKANOT, KUCHITSUK, SHINOZAKIK, YOSHIDAS, MATSUMOTOS . A 9-cis-epoxycarotenoid dioxygenase inhibitor for use in the elucidation of abscisic acid action mechanisms. Bioorganic and Medicinal Chemistry, 2006,14:5555-5561. [本文引用: 1]
BOYDJ, GAIY, NELSON KM, LUKIWSKIE, TALBOTJ, LOEWEN MK, OWENS, ZAHARIA LI, CUTLER AJ, ABRAMS SR, LOEWEN MC . Sesquiterpene-like inhibitors of a 9-cis- epoxycarotenoid dioxygenase regulating abscisic acid biosynthesis in higher plants. Bioorganic and Medicinal Chemistry, 2009,17:2902-2912. [本文引用: 1]
KUSHIROT, OKAMOTOM, NAKABAYASHIK, YAMAGISHIK, KITAMURAS, ASAMIT, HIRAIN, KOSHIBAT, KAMIYAY, NAMBARAE . The Arabidopsis cytochrome P450 CYP707A encodes ABA 8'-hydroxylases: Key enzymes in ABA catabolism. The EMBO Journal, 2004,23:1647-1656. [本文引用: 1]
SAITOS, HIRAIN, MATSUMOTOC, OHIGASHIH, OHTAD, SAKATAK, MIZUTANIM . Arabidopsis CYP707As encode (+)-abscisic acid 8'-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid. Plant Physiology, 2004,134:1439-1449. [本文引用: 1]
HANADAK, HASET, TOYODAT, SHIONZAKIK, OKAMOTOM . Origin and evolution of genes related to ABA metabolism and its signaling pathways Journal of Plant Research, 2011,124:455-465. [本文引用: 1]
OKAMOTOM, KUWAHARAA, SEOM, KUSHIROT, ASAMIT, HIRAIN, KAMIYAY, KOSHIBAT, NAMBARAE . CYP707A1 and CYP707A2, which encode ABA 8'-hydroxylases, are indispensable for a proper control of seed dormancy and germination in Arabidopsis Plant Physiology, 2006,141:97-107. [本文引用: 4]
OKAMOTOM, TANAKAY, ABRAMS SR, KAMIYAY, SEKIM, NAMBARAE . High humidity induces abscisic acid 8'-hydroxylase in stomata and vasculature to regulate local and systemic abscisic acid responses in Arabidopsis. Plant Physiology, 2009,149:825-834. [本文引用: 1]
KEPKAM, BENSON CL, GONUGUNTA VK, NELSON KM, CHRISTMANNA, GRILLE, ABRAMA SR . Action of natural abscisic acid precursors and catabolites on abscisic acid receptor complexes Plant Physiology, 2011,157:2108-2119. [本文引用: 1]
KITAHATAN, SAITOS, MIYAZAWAY, UMEZAWAT, SHIMADAY, MIN YK, MIZUTANIM, HIRAIN, SHINOZAKIK, YOSHIDAS . Chemical regulation of abscisic acid catabolism in plants by cytochrome P450 inhibitors Bioorganic and Medicinal Chemistry, 2005,13:4491-4498. [本文引用: 1]
SAITOS, OKAMOTOM, SHINODAS, KUSHIROT, KOSHIBAT, KAMIYAY, HIRAIN, TODOROKIY, SAKATAK, NAMBARAE, MIZUTANIM . A plant growth retardant, uniconazole, is a potent inhibitor of ABA catabolism in Arabidopsis. Bioscience Biotechnology and Biochemistry, 2006,70:1731-1739. [本文引用: 1]
RADEMACHERW . Growth retardants: Effects on gibberellin biosynthesis and other metabolic pathways Annual Review of Plant Physiology and Plant Molecular Biology, 2000,51:501-531. [本文引用: 1]
TAKEUCHIJ, OKAMOTOM, MEGAR, KANNOY, OHNISHIT, SEOM, TODOROKIY . Abscinazole-E3M, a practical inhibitor of abscisic acid 8'-hydroxylase for improving drought tolerance Scientific Reports, 2016,6:37060. [本文引用: 1]
BENSON CL, KEPKAM, WUNSCHELC, RAJAGOPALANN, NELSON KM, CHRISTMANNA, ABRAMS SR, GRILLE, LOEWENM . Abscisic acid analogs as chemical probes for dissection of abscisic acid responses in Arabidopsis thaliana. Phytochemistry, 2015,113:96-107. [本文引用: 1]
ARAKIY, MIYAWAKIA, MIYASHITAT, MIZUTANIM, HIRAIN, TODOROKIY . A new non-azole inhibitor of ABA 8'-hydroxylase: Effect of the hydroxyl group substituted for geminal methyl groups in the six-membered ring Bioorganic and Medicinal Chemistry Letters, 2006,16:3302-3305. [本文引用: 1]
MAY, SZOSTKIEWICZI, KORTEA, MOESD, YANGY, CHRISTMANNA, GRILLE . Regulators of PP2C phosphatase activity function as abscisic acid sensors Science, 2009,324:1064-1068. [本文引用: 3]
NISHIMURAN, TSUCHIYAW, MORESCO JJ, HAYASHIY, SATOHK, KAIWAN, IRISAT, KINOSHITAT, SCHROEDER JI, YATES JR, HIRAYAMAT, YAMAZAKIT . Control of seed dormancy and germination by DOG1-AHG1 PP2C phosphatase complex via binding to heme Nature Communication, 2018,9:2132. [本文引用: 10]
PARK SY, FUNGP, NISHIMURAN, JENSEN DR, FUJIIH, ZHAOY, LUMBAS, SANTIAGOJ, RODRIGUESA, CHOW TF, ALFRED SE, BONETTAD, FINKELSTEINR, PROVART NJ, DESVEAUXD, RODRIGUEZ PL, McCOURTP, ZHU JK, SCHROEDER JI, VOLKMAN BF, CUTLER SR . Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins Science, 2009,324:1068-1071. [本文引用: 3]
MELCHERK, NG LM, ZHOU XE, SOONF-F, XUY, SUINO-POWELL KM, PARK SY, WEINER JJ, FUJIIH, CHINNUSAMYV, KOVACHA, LIJ, WANGY, LIJ, PERTERSON FC, JENSEN DR, YONG EL, VOLKMAN BF, CUTLER SR, ZHU JK, XU HE. A gate-latch-lock mechanism for hormone signalling by abscisic acid receptors Nature, 2009,462:602-608. [本文引用: 2]
SANTIAGOJ, DUPEUXF, ROUNDA, ANTONIR, PARK SY, JAMINM, CUTLER SR, RODRIGUEZ PL, MáRQUEZ JA. The abscisic acid receptor PYR1 in complex with abscisic acid Nature, 2009,462:665-668. [本文引用: 1]
YINP, FANH, HAOQ, YUANX, WUD, PANGY, YANC, LIW, WANGJ, YANN . Structural insights into the mechanism of abscisic acid signaling by PYL proteins Nature Structural and Molecular Biology, 2009,16:1230-1236. [本文引用: 1]
FUJIIH, ZHU JK . Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress Proceedings of the National Academy of Sciences of the United States of America, 2009,106:8380-8385. [本文引用: 2]
SOON FF, NG LM, ZHOU XE, WEST GM, KOVACHA, TAN M HE, SUINO-POWELL KM, HEY, XUY, CHALMERS MJ, BRUNZELLE JS, ZHANGH, YANGH, JIANGH, LIJ, YONG EL, CUTLERS, ZHU JK, GRIFFIN PR, MELCHERK, XU HE. Molecular mimicry regulates ABA signaling by SnRK2 kinases and PP2C phosphatases Science, 2012,335:85-88. [本文引用: 1]
NAKAMURAS, LYNCH TJ, FINKELSTEIN RR . Physical interactions between ABA response loci of Arabidopsis. The Plant Journal, 2001,26:627-635. [本文引用: 1]
HAUSERF, WAADTR, SCHROEDER JI . Evolution of abscisic acid synthesis and signaling mechanisms Current Biology, 2011,21:R346-R355. [本文引用: 2]
BOWMAN JL, KOHCHIT, YAMATO KT, JENKINSJ, SHUS, ISHIZAKIK, YAMAOKAS, NISHIHAMAR, NAKAMURAY, BERGERF, ADAMC, AKI SS, ALTHOFFF, ARAKIT, ARTEAGA-VAZQUEZ MA, BALASUBRMANIANS, BARRYK, BAYERD, SCHMUTZJ. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell, 2017,171:287-304. [本文引用: 1]
TISCHER SV, WUNSCHELC, PAPACEKM, KLEIGREWEK, HOFMANNT, CHRISTMANNA, GRILLE . Combinatorial interaction network of abscisic acid receptors and coreceptors from Arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America, 2017,114:10280-10285. [本文引用: 3]
ANTONIR, GONZALEZ-GUZMANM, RODRIGUEZL, RODRIGUESA, PIZZIO GA, RODRIGUEZ PL . Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors Plant Physiology, 2012,158:970-980. [本文引用: 5]
NISHIMURAN, YOSHIDAT, KITAHATAN, ASAMIT, SHINOZAKIK, HIRAYAMAT . ABA-HYPERSENSITIVE GERMINATION1 encodes a protein phosphatase 2C, an essential component of abscisic acid signaling in Arabidopsis seed The Plant Journal, 2007,50:935-949. [本文引用: 2]
FUJITAY, NAKASHIMAK, YOSHIDAT, KATAGIRIT, KIDOKOROS, KANAMORIN, UMEZAWAT, FUJITAM, MARUYAMAK, ISHIYAMAK, KOBAYASHIM, NAKASONES, YAMADAK, ITOT, SHINOZAKIK . Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant and Cell Physiology, 2009,50:2123-2132. [本文引用: 1]
NAKASHIMAK, FUJITAY, KANAMORIN, KATAGIRIT, UMEZAWAT, KIDOKOROS, MARUYAMAK, YOSHIDAT, ISHIYAMAK, KOBAYASHIM, SHINOZAKIK, YAMAGUCHI- SHINOZAKIK . Three Arabidopsis SnRK2 protein kinases, SRK2D/ SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant and Cell Physiology, 2009,50:1345-1363.
UMEZAWAT, SUGIYAMAN, MIZOGUCHIM, HAYASHIS, MYOUGAF, YAMAGUCHI-SHINOZAKIK, ISHIHAMAY, HIRAYAMAT, SHINOZAKIK . Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proceedings of the National Academy of Sciences of the United states of America, 2009,106:17588-17593. [本文引用: 1]
KLINE KG, BARRETT-WILT GA, SUSSMAN MR. In planta changes in protein phosphorylation induced by the plant hormone abscisic acid Proceedings of the National Academy of Sciences of the United States of America, 2010,107:15986-15991. [本文引用: 1]
UMEZAWAT, SUGIYAMAN, TAKAHASHIF, ANDERSON JC, ISHIHAMAY, PECK SC, SHINOZAKIK. Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana Science Signaling, 2013, 6:1rs8. [本文引用: 1]
WANGP, XUEL, BATELLIG, LEES, HOU YJ, VAN OOSTENM J, ZHANGH, TAO WA, ZHU JK. Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action Proceedings of the National Academy of Sciences of the United states of America, 2013,110:11205-11210. [本文引用: 1]
DINGS, ZHANGB, QINF . Arabidopsis RZFP34/CHYR1, an ubiquitin E3 ligase, regulates stomatal movement and drought tolerance via SnRK2.6-mediated phosphorylation The Plant Cell, 2015,27:3228-3244. [本文引用: 1]
BELDA-PALAZONB, RODRIGUEZL, FERNANDEZ MA, CASTILLO MC, ANDERSON EA, GAOC, GONZALEZ- GUZMANM, PEIRATS-LLOBETM, ZHAOQ, DE WINNEN, GEVEERTK, DE JAEGERG, JIANGL, LEòNJ, MULLEN RT, RODRIGUEZ PL. FYVE1/FREE1 interacts with the PYL4 ABA receptor and mediates its delivery to the vacuolar degradation pathway The Plant Cell, 2016,28:2291-2311. [本文引用: 1]
WUQ, ZHANGX, PEIRATS-LLOBETM, BELDA-PALAZONB, WANGX, CUIS, YUX, RODRIGUEZ PL, ANC . Ubiquitin ligases RGLG1 and RGLG5 regulate abscisic acid signaling by controlling the turnover of phosphatase PP2CA The Plant Cell, 2016,28:2178-2196. [本文引用: 1]
YUF, WUY, XIEQ . Ubiquitin-proteasome system in ABA signaling: From perception to action Molecular Plant, 2016,9:21-33. [本文引用: 1]
ZHAOJ, ZHAOL, ZHANGM, ZAFARS, FANGJ, LIM, ZHANGW, LIX . Arabidopsis E3 ubiquitin ligases PUB22 and PUB23 negatively regulate drought tolerance by targeting ABA receptor PYL9 for degradation International Journal of Molecular Science, 2017,18:1841. [本文引用: 1]
KARSSEN CM, BRINKHORST-VAN DER SWAND L C, BREEKLAND AE, KOORNNEEFM. Induction of dormancy during seed development by endogenous abscisic acid: Studies on abscisic acid deficient genotypes of Arabidopsis thaliana (L.) Heynh Planta, 1983,157:158-165. [本文引用: 1]
FREYA, GODINB, BONNETM, SOTTAB, MARION-POLLA . Maternal synthesis of abscisic acid controls seed development and yield in Nicotiana plumbaginifolia. Planta, 2004,218:958-964. [本文引用: 1]
KOORNNEEFM, HANHART CJ, HILHORST HW M, KARSSEN CM. In vivo inhibition of seed development and reserve protein accumulation in recombinants of abscisic acid biosynthesis and responsiveness mutants in Arabidopsis thaliana. Plant Physiology, 1989,90:463-469. [本文引用: 1]
CADMAN CS, TOOROP PE, HILHORST HW M, FINCH- SAVAGEW E. Gene expression profiles of Arabidopsis Cvi seeds during dormancy cycling indicate a common underlying dormancy control mechanism. The Plant Journal, 2006,46:805-822. [本文引用: 1]
LEFEBVREV, NORTHH, FREYA, SOTTAB, SEOM, OKAMOTOM, NAMBARAE, MARION-POLLA . Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesised in the endosperm is involved in the induction of seed dormancy. The Plant Journal, 2006,45:309-319. [本文引用: 1]
NAKABAYASHIK, OKAMOTOM, KOSHIBAT, KAMIYAY, NAMBARAE . Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: Epigenetic and genetic regulation of transcription in seed. The Plant Journal, 2005,41:697-709. [本文引用: 1]
FINCH-SAVAGE WE, LEUBNER-METZGERG . Seed dormancy and the control of germination New Phytologist, 2006,171:501-523. [本文引用: 3]
GUBLERF, HUGHEST, WATERHOUSEP, JACOBSENJ . Regulation of dormancy in barley by blue light and after-ripening: Effects on abscisic acid and gibberellin metabolism Plant Physiology, 2008,147:886-896. [本文引用: 3]
JACOBSEN JV, BARRERO JM, HUGHEST, JULKOWSKAM, TAYLOR JM, XUQ, GUBLERF . Roles for blue light, jasmonate and nitric oxide in the regulation of dormancy and germination in wheat (Triticum aestivum L.) grain. Planta, 2013,238:121-138. [本文引用: 2]
LIUA, GAOF, KANNOY, JORDAN MC, KAMIYAY, SEOM, AYELE BT . Regulation of wheat seed dormancy by after-ripening is mediated by specific transcriptional switches that induce changes in seed hormone metabolism and signaling PLoS ONE, 2013,8:e56570. [本文引用: 1]
BARRERO JM, TALBOT MJ, WHITE RG, JACOBSEN JV, GUBLERF . Anatomical and transcriptomic studies of the coleorhiza reveal the importance of this tissue in regulating dormancy in barley Plant Physiology, 2009,150:1006-1021. [本文引用: 1]
LIUY, SHIL, YEN, LIUR, JIAW, ZHANGJ . Nitric oxide-induced rapid decrease of abscisic acid concentration is required in breaking seed dormancy in Arabidopsis. New Phytologist, 2009,183:1030-1042. [本文引用: 3]
PRESTONJ, TATEMATSUK, KANNOY, HOBOT, KIMURAM, JIKUMARUY, YANOR, KAMIYAY, NAMBARAE . Temporal expression patterns of hormone metabolism genes during imbibition of Arabidopsis thaliana seeds: A comparative study on dormant and non-dormant accessions. Plant and Cell Physiology, 2009,50:1786-1800. [本文引用: 2]
MATAKIADIST, ALBORESIA, JIKUMARUY, TATEMATSUK, PICHONO, RENOU JP, SOTTAB, KAMIYAY, NAMBARAE, TROUNG HN . The Arabidopsis abscisic acid catabolism gene CYP707A2 plays a key role in nitrate control of seed dormancy. Plant Physiology, 2009,149:949-960. [本文引用: 1]
TOHS, IMAMURAA, WATANABEA, NAKABAYASHIK, OKAMOTOM, JIKUMARUY, HANADAA, ASOY, ISHIYAMAK, TAMURAN, IUCHIS, KOBAYASHIM, YAMAGUCHIS, KAMIYAY, NAMBARAE, KAWAKAMIN . High temperature- induced ABA biosynthesis and its role in the inhibition of GA action in Arabidopsis seeds. Plant Physiology, 2008,146:1368-1385. [本文引用: 1]
BENECH-ARNOLD RL, GUALANON, LEYMARIEJ, C?MED, CORBINEAUF . Hypoxia interferes with ABA metabolism and increases ABA sensitivity in embryos of dormant barley grains Journal of Experimental Botany, 2006,57:1423-1430. [本文引用: 1]
TOOROP PE, BEWLEY JD, HILHORST HW M. Endo-β-isoforms are present in the endosperm and embryo of tomato seeds, but are not essentially linked to germination Planta, 1996,200:153-158. [本文引用: 1]
MüLLERK, TINTELNOTS, LEUBNER-METZGERG . Endosperm- limited Brassicaceae seed germination: Abscisic acid inhibits embryo-induced endosperm weakening of Lepidium sativum(cress) and endosperm rupture of cress and Arabidopsis thaliana. Plant and Cell Physiology, 2006,47:864-877. [本文引用: 1]
MüLLERK, CARSTENS AC, LINKIESA, TORRES MA, LEUBNER-METZGERG . The NADPH-oxidase AtrbohB plays a role in Arabidopsis seed after-ripening. New Phytologist, 2009,184:885-897. [本文引用: 1]
NONOGAKIH . Seed biology updates-highlights and new discoveries in seed dormancy and germination research Frontiers in Plant Science, 2017,8:524. [本文引用: 8]
NéEG, KRAMERK, NAKABAYASHIK, YUANB, XIANGY, MIATTONE, FINKEMEIERI, SOPPE WJ . DELAY OF GERMINATION1 requires PP2C phosphatases of the ABA signalling pathway to control seed dormancy Nature Communication, 2017,8:72. [本文引用: 8]
ALONSO-BLANCOC, BENTSINKL, HANHART CJ, VRIES HB D, KOORNNEEFM . Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana. Genetics, 2003,164:711-729. [本文引用: 1]
BENTSINKL, HANSONJ, HANHART CJ, BLANKESTIJN-DE VRIESH, COLTRANEC, KEIZERP, EL-LITHYM, ALONSO-BLANCOC, DE ANDRéS MT, REYMONDM, VAN EEUWIJKF, SMEEKENSS, KOORNNEEFM. Natural variation for seed dormancy in Arabidopsis is regulated by additive genetic and molecular pathways. Proceedings of the National Academy of Sciences of the United States of America , 2010,107:4264-4269. [本文引用: 1]
BENTSINKL, JOWETTJ, HANHART CJ, KOORNNEEFM . Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 2006,103:17042-17047. [本文引用: 5]
NAKABAYASHIK, BARTSCHM, XIANGY, MIATTONE, PELLENGAHRS, YANOR, SEOM, SOPPE WJ J. The time required for dormancy release in Arabidopsis is determined by DELAY OF GERMINATION1 protein levels in freshly harvested seeds. The Plant Cell, 2012,24:2826-2838. [本文引用: 6]
NAKABAYASHIK, BARTSCHM, DINGJ, SOPPE WJ . Seed dormancy in Arabidopsis requires self-binding ability of DOG1 protein and the presence of multiple isoforms generated by alternative splicing. PLoS Genetics, 2015,11:e1005737. [本文引用: 6]
CYREKM, FEDAKH, CIESIELSKIA, GUO YW, SLIWAA, BRZEZNIAKL, KRZYCZMONIKK, PIETRASZ, KACZANOWSKIS, LIUF, SWIEZEWSKIS . Seed dormancy in Arabidopsis is controlled by alternative polyadenylation of DOG1. Plant Physiology, 2016,170:947-955. [本文引用: 7]
DOLATAJ, GUOY, KO?OWERZOA, SMOLI?SKID, BRZY?EKG, JARMO?OWSKIA, ?WIE?EWSKIS. NTR1 is required for transcription elongation checkpoints at alternative exons in Arabidopsis. The EMBO Journal, 2015,34:544-558. [本文引用: 2]
LIUY, KOORNNEEFM, SOPPE WJ . The absence of histone H2B monoubiquitination in the Arabidopsis hub1 (rdo4) mutant reveals a role for chromatin remodeling in seed dormancy. The Plant Cell, 2007,19:433-444. [本文引用: 2]
LIUY, GEYERR, VAN ZANTENM, CARLESA, LIY, H?ROLDA, VAN NOCKERS, SOPPE WJ J. Identification of the Arabidopsis REDUCED DORMANCY 2 gene uncovers a role for the polymerase associated factor 1 complex in seed dormancy. PLoS ONE, 2011,6:e22241.
MORTENSEN SA, GRASSER KD . The seed dormancy defect of Arabidopsis mutants lacking the transcript elongation factor TFIIS is caused by reduced expression of the DOG1 gene. FEBS Letters, 2014,588:47-51. [本文引用: 2]
DI GIAMMARTINO DC, NISHIDAK, MANLEY JL . Mechanisms and consequences of alternative polyadenylation Molecular Cell, 2011,43:853-866. [本文引用: 1]
FEDAKH, PALUSINSKAM, KRZYCZMONIKK, BRZEZNIAKL, YATUSEVICHR, PIETRASZ, KACZANOWSKIS, SWIEZEWSKIS . Control of seed dormancy in Arabidopsis by a cis-acting non- coding antisense transcript. Proceedings of the National Academy of Sciences of the United States of America, 2016,113:E7846-E7855. [本文引用: 12]
LINS, ZHANGL, LUOW, ZHANGX . Characteristics of antisense transcript promoters and the regulation of their activity International Journal of Molecular Sciences, 2015,17:1-17. [本文引用: 1]
KORNIENKO AE, GUENZL PM, BARLOW DP, PAULER FM . Gene regulation by the act of long non-coding RNA transcription BMC Biology, 2013,11:59. [本文引用: 1]
PELECHANOV, STEINMETZ LM . Non-coding RNA gene regulation by antisense transcription Nature Review of Genetics, 2013,14:880-893. [本文引用: 4]
QUINN JJ, CHANG HY . Unique features of long non-coding RNA biogenesis and function Nature Review Genetics, 2016,17:47-62. [本文引用: 1]
SHEARWIN KE, CALLEN BP, EGAN JB . Transcriptional interference - a crash course Trends in Genetics, 2005,21:339-345. [本文引用: 1]
HONGAY CF, GRISAFI PL, GALITSKIT, FINK GR . Antisense transcription controls cell fate in Saccharomyces cerevisiae. Cell, 2006,127:735-745. [本文引用: 1]
NéEG, XIANGY, SOPPE WJ . The release of dormancy, a wake-up call for seeds to germinate Current Opinion in Plant Biology, 2016,35:8-14. [本文引用: 2]
YOSHIDAT, NISHIMURAN, KITAHATAN, KUROMORIT, ITOT, ASAMIT, SHINOZAKIK, HIRAYAMAT . ABA-HYPERSENSITIVE GERMINATION3 encodes a protein phosphatase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phosphatase 2Cs Plant Physiology, 2005,140:115-126. [本文引用: 1]
LIT, BONKOVSKY HL, GUOJ . Structural analysis of heme proteins: Implications for design and prediction BMC Structural Biology, 2011,11:13. [本文引用: 1]
OHKUMAK, LYON JL, ADDICOTT FT, SMITH OE . Abscisin II, an abscission-accelerating substance from young cotton fruit Science, 1963,142:1592-1593. [本文引用: 1]
WANG YG, FU FL, YU HQ, HUT, ZHANG YY, TAOY, ZHU JK, ZHAOY, LI WC . Interaction network of core ABA signaling components in maize Plant Molecular Biology, 2018,96:245-263. [本文引用: 1]
LIU SJ, SONG SH, WANG WQ, SONG SQ . De novo assembly and characterization of germinating lettuce seed transcriptome using Illumina paired-end sequencing Plant Physiology and Biochemistry, 2015,96:154-162. [本文引用: 1]
WANG WQ, SONG BY, DENG ZJ, WANGY, LIU SJ, M?LLERI M, SONG SQ . Proteomic analysis of Lactuca sativa seed germination and thermoinhibition by sampling of individual seeds at germination and removal of storage proteins by PEG fractionation. Plant Physiology, 2015,167:1332-1350. [本文引用: 1]
XU HH, LIU SJ, SONG SH, WANG WQ, M?LLERI X, SONG SQ . Proteome changes associated with dormancy release of Dongxiang wild rice seeds Journal of Plant Physiology, 2016,206:68-86. [本文引用: 1]
 ,3, 黄荟41
,3, 黄荟41 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT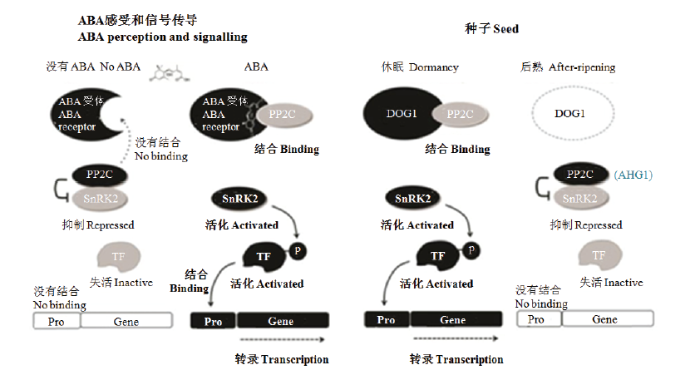 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}