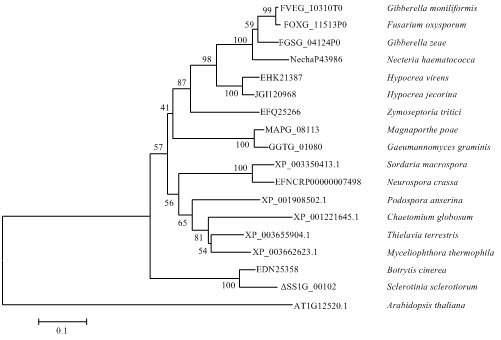
Abstract 【Objective】Sclerotinia stem rot is a kind of fungal disease caused by Sclerotinia sclerotiorum. The host range of S. sclerotiorum is wide, which seriously endangers the quality of many crops. The objective of this study is to enhance the resistance to stem rot by silencing the virulence genes of S. sclerotiorum in host via the host-induced gene silencing (HIGS) technology, and to provide new ideas for breeding of sclerotinia stem rot resistance.【Method】The gene encoding a copper chaperone for copper/zinc superoxide dismutase of S. sclerotiorum (SsCCS) was selected as the target gene, the sequences were analyzed by bioinformatics tools, and the phylogenetic tree was constructed using MEGA6.0 software. The specific interference fragment was selected for amplification after comparing the genome of Arabidopsis thaliana and S. sclerotiorum, respectively. HIGS vector containing the RNAi structure of SsCCS was transferred into wild type A. thaliana Col-0 mediated by Agrobacterium, and the stable HIGS-CCS transgenic A. thaliana lines were screened by DNA identification and labeling. The leaves of HIGS-CCS transgenic plants grown for 4-5 weeks were selected to analyze the resistance to sclerotinia stem rot according to the lesion area at 24 h after inoculated with S. sclerotiorum. The relative expression level of SsCCS during infecting was analyzed by qRT-PCR. The accumulation of H2O2 during the period of interaction between transgenic plants and S. sclerotiorum was detected by DAB staining at 6, 12 and 24 hpi.【Result】Bioinformatics analysis showed that the length of genome sequence of SsCCS (SS1G_00102) is 1 010 bp, while the length of its coding sequence (CDS) is 759 bp, encoding a protein with 253 amino acids, the molecular weight is 2 7176.96 Da, the isoelectric point (PI) is 5.04. SsCCS has 87% amino acid homology to BcCCS (EDN25358) while far to AtCCS (AT1G12520.1). By aligning with the genome of S. sclerotiorum and A. thaliana, a 314 bp specific interference fragment was selected and constructed the HIGS vector successfully and transformed into A. thaliana. The lesion of T1 and T2 generation transgenic lines was smaller than that of wild type A. thaliana 24 h after inoculated with strain 1980. From the T2 generation, three stably expressed T3 generation HIGS-CCS transgenic A. thaliana lines (HIGS-CCS-5, HIGS-CCS-8, HIGS-CCS-13) were obtained. Compared with wild type A. thaliana, the lesion area on the HIGS-CCS transgenic plants was reduced by 46% to 61% 24 h after inoculated with strain 1980. The expression of SsCCS was significantly reduced by 98% in the HIGS-CCS transgenic plants compared to that in wild type A. thaliana at 6 h after inoculated with S. sclerotiorum strain 1980. Furthermore, the accumulation of H2O2 was decreased in transgenic plants as revealed by DAB staining, indicating the ROS production was reduced in transgenic plants.【Conclusion】The resistance to stem rot can be significantly enhanced by silencing the SsCCS of S. sclerotiorum in A. thaliana via the HIGS. This study provides a reference for the resistance improvement to sclerotinia stem rot of host crops, such as rapeseed. Keywords:Arabidopsis thaliana;host-induced gene silencing (HIGS);Sclerotinia sclerotiorum; SsCCS;disease resistance;sclerotinia stem rot
PDF (1537KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 柴亚茹, 丁一娟, 周思钰, 杨文静, 闫宝琴, 远俊虎, 钱伟. HIGS-SsCCS转基因拟南芥的菌核病抗性鉴定[J]. 中国农业科学, 2020, 53(4): 761-770 doi:10.3864/j.issn.0578-1752.2020.04.008 CHAI YaRu, DING YiJuan, ZHOU SiYu, YANG WenJing, YAN BaoQin, YUAN JunHu, QIAN Wei. Identification of the Resistance to Sclerotinia Stem Rot in HIGS-SsCCS Transgenic Arabidopsis thaliana[J]. Scientia Acricultura Sinica, 2020, 53(4): 761-770 doi:10.3864/j.issn.0578-1752.2020.04.008
利用BioEdit软件将SsCCS与核盘菌以及拟南芥基因组序列进行比对,选择SsCCS的特异片段作为RNA干扰片段,利用Primer Premier 5设计特异引物,并引入相应的酶切位点(表1)。以核盘菌野生菌株1980的cDNA为模板,以SsCCS-SF/SsCCS-SR及SsCCS-AF/SsCCS-AR分别进行扩增,并将胶回收的目的片段连接到pGEMT-Easy载体,转化至大肠杆菌DH5α,挑选阳性单克隆送Invitrogen公司测序。提取测序正确T克隆质粒T-SsCCS-S及T-SsCCS-A,首先采用限制性内切酶EcoRI/EcoRV双酶切T-SsCCS-S,将该正向片段连入pCIT,获得中间载体p-ccs-1。随后采用PstI/BamHI双酶切T-SsCCS-A,将该片段反向连入p-ccs-1,获得SsCCS的干扰载体p-ccs。随后,用EcoRI和XhoI分别双酶切p-ccs及植物转化载体pBin35SRed3的质粒,将p-ccs载体中的主体结构“正向序列-内含子-反向序列”连接到pBin35SRed3的CaMV35S启动子与Nos终止子之间,获得沉默SsCCS的HIGS载体R-ccs。将重组质粒R-ccs转化至农杆菌GV3101,挑选阳性单克隆的菌液加入50%的甘油1﹕1等体积混合于2 mL离心管中,保存于-80℃。
Table 1 表1 表1SsCCS载体构建所用引物序列 Table 1Sequence information of primers used for vector construction of SsCCS
A:SsCCS沉默表达载体构建模式图The pattern of vector construction to silence SsCCS;B:SsCCS特异干扰片段扩增Amplification of specific interference fragment of SsCCS;C:p-ccs载体转化大肠杆菌DH5α菌液检测结果Colony PCR verification of Escherichia coli DH5α transformed with p-ccs vector;D:EcoRI和XhoI双酶切鉴定R-ccs Identification of R-ccs vector by double digestion with EcoRI/XhoI enzymes;E:R-ccs载体转化农杆菌GV3101的菌落PCR鉴定Colony PCR identification of Agrobacterium tumefaciens GV3101 transformed with R-ccs vector;M:D2000 Marker Fig. 2Construction of HIGS vector of SsCCS
M:D2000 Marker;WT:野生型拟南芥Col-0 Wild type A. thaliana Col-0 (Control);1—17:17株T1代转基因植株17 transformed A. thaliana lines of T1 generation Fig. 3PCR identification of transformed A. thaliana lines of T1 generation
Table 2 表2 表2转基因拟南芥分离比及接种核盘菌后病斑统计 Table 2Segregation and lesion analyses of transgenic and wild type A. thaliana lines after inoculated with S. sclerotiorum
编号 Code
T1代 T1 generation
T2代病斑面积 T2 generation lesion area (cm2)
阳性种子数 Positive
非阳性种子数 Negative
分离比 Segregation ratio
病斑面积 Lesion area (cm2)
WT
-
-
-
1.46±0.26a
1.02±0.36a
HIGS-CCS-13
73
27
3﹕1
0.69±0.2b
0.79±0.27b
HIGS-CCS-8
69
31
3﹕1
0.71±0.17b
0.48±0.21b
HIGS-CCS-5
78
22
3﹕1
0.89±0.13b
0.55±0.23b
HIGS-CCS-14
79
21
3﹕1
0.82±0.27b
0.93±0.27a
HIGS-CCS-7
92
8
15﹕1
0.77±0.18b
HIGS-CCS-18
69
31
3﹕1
0.91±0.53a
HIGS-CCS-22
85
25
3﹕1
1.11±0.08a
HIGS-CCS-3
71
29
3﹕1
1.11±0.25a
HIGS-CCS-19
44
56
1﹕1
1.12±0.11a
HIGS-CCS-4
56
44
1﹕1
1.15±0.13a
HIGS-CCS-17
100
0
-
1.27±0.28a
HIGS-CCS-9
78
22
3﹕1
1.37±0.18a
HIGS-CCS-11
95
5
15﹕1
1.21±0.24a
HIGS-CCS-2
70
30
3﹕1
0.91
HIGS-CCS-20
76
34
3﹕1
1.02
HIGS-CCS-1
89
11
15﹕1
1.13
HIGS-CCS-12
74
26
3﹕1
1.26
同列数据后不同小写字母表示0.05水平上差异显著Different lowercases after the data in the same column indicate significant differences at the 0.05 level
A:接种24 h后的病斑Disease phenotypic of transgenic and wild type A. thaliana at 24 h after inoculated with S. sclerotiorum;B:A中的病斑统计结果Lesion area at 24 hpi in A;“*”表示转基因拟南芥材料与对照(WT)相比在0.05水平上差异显著The transgenic group is significantly different from the control group (WT) at the 0.05 level Fig. 4Resistance identification of T3 generation transgenic A. thaliana lines
HIGS-CCS-5:HIGS-CCS-5转基因拟南芥株系叶片Transgenic A. thaliana line HIGS-CCS-5;WT:野生型拟南芥植株叶片Wild type of A. thaliana Fig. 5DAB staining assay of wild type and transgenic A. thaliana lines
以SsTubulin为参考基因,采用2-ΔΔCT法进行相对定量,数据表示3个生物学重复的平均值,误差线表示标准偏差(±SD) Fig. 6Relative expression level of SsCCS at 6 h after the wild type and transgenic A. thaliana lines inoculated with S. sclerotiorum strain 1980
2-ΔΔCT method was used to obtain relative quantification using SsTubulin as reference gene. Data indicate average from three biological replicates and error bars represent standard deviation (±SD)
BOLAND GJ, HALLR . Index of plant hosts of Sclerotinia sclerotiorum Canadian Journal of Plant Pathology, 1994,16(2):93-108. [本文引用: 1]
BOLTON MD, THOMMA BP, NELSON BD . Sclerotinia sclerotiorum(Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, 2006,7(1):1-16. [本文引用: 1]
ZHANGY, WANGX, CHANGX, SUNM, ZHANGY, LIW, LIY . Overexpression of germin-like protein GmGLP10 enhances resistance to Sclerotinia sclerotiorum in transgenic tobacco Biochemical and Biophysical Research Communications, 2018,497(1):160-166. [本文引用: 1]
ZHANG HS, HU YY, ZHANGN, WEI XJ, YANG WX, LIU DQ . Progress of host-induced gene silencing (HIGS) technology Journal of Agricultural Biotechnology, 2013,21(5):603-611. (in Chinese) [本文引用: 2]
ZHANG XJ, ZHANGY, HU SW . Progress on resistance mechanism of Sclerotinia sclerotiorum and genetic breeding program on disease resistant rapeseed. Molecular Plant Breeding, 2016,14(3):704-711. (in Chinese) [本文引用: 1]
FIREA, XU SQ, MONTGOMERY MK, KOSTAS SA, DRIVER SE, MELLO CC . Potent and specific genetic interference by double- stranded RNA in Caenorhabditis elegans Nature, 1998,391(6669):806-811. [本文引用: 1]
WIANNYF, ZERNICKA-GOETZM . Specific interference with gene function by double-stranded RNA in early mouse development Nature Cell Biology, 2000,2(2):70-75. [本文引用: 1]
SVOBODAP, STEINP, HAYASHIH, SCHULTZ RM . Selective reduction of dormant maternal mRNAs in mouse oocytes by RNA interference Development, 2000,127(19):4147-4156. [本文引用: 1]
NUNES CC, DEAN RA . Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies Molecular Plant Pathology, 2012,13(5):519-529. [本文引用: 1]
WANGM, WEIBERGA, LIN FM, THOMMA BP, HUANG HD, JINH . Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection Nature Plants, 2016,2(10):16151. [本文引用: 1]
NOWARAD, GAYA, LACOMMEC, SHAWJ, RIDOUTC, DOUCHKOVD, HENSELG, KUMLEHNJ, SCHWEIZERP . HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis The Plant Cell, 2010,22(9):3130-3141. [本文引用: 1]
PANWARV, MCCALLUMB, BAKKERENG . Endogenous silencing of Puccinia triticina pathogenicity genes through in planta-expressed sequences leads to suppression of rust diseases on wheat The Plant Journal, 2013,73(3):521-532. [本文引用: 1]
KOCHA, KUMARN, WEBERL, KELLERH, IMANIJ, KOGEL KH . Host-induced gene silencing of cytochrome P450 lanosterol C14 α-demethylase-encoding genes confers strong resistance to Fusarium species Proceedings of the National Academy of Sciences of the United States of America, 2013,110(48):19324-19329. [本文引用: 1]
ANDRADE CM, TINOCOM L P, RIETHA F, MAIAF C O, ARAG?OF J L, . Host-induced gene silencing in the necrotrophic fungal pathogen Sclerotinia sclerotiorum Plant Pathology, 2016,65(4):626-632. [本文引用: 2]
JIAO CX . Reactive oxygen species in plant stress and defense Journal of Dali University (Natural Science), 2006,5(10):73-76, 79. (in Chinese) [本文引用: 1]
IMLAY JA, LINNS . DNA damage and oxygen radical toxicity Science, 1988,240(4857):1302-1309. [本文引用: 1]
BYRDS, REINSD, DOETSCH PW . Effects of oxidative DNA damage on transcription by RNA polymerases Free Radical Biology and Medicine, 1990,9(1):47. [本文引用: 1]
XUL, CHENW . Random T-DNA mutagenesis identifies a Cu/Zn superoxide dismutase gene as a virulence factor of Sclerotinia sclerotiorum Molecular Plant-Microbe Interactions, 2013,26(4):431-441. [本文引用: 2]
BOWLERC, VAN CAMPW, VAN MONTAGUM, INZéD . Superoxide dismutase in plant Critical Reviews in Plant Science, 1994,13(3):199-218. [本文引用: 1]
ZOU GL, QIU MY, ZHUT . The history, current status and application prospects of superoxide dismutase research. Journal of Amino Acids, 1991(3):28-32. (in Chinese) [本文引用: 1]
VELUCHAMYS, WILLIAMSB, KIMK, DICKMAN MB . The CuZn superoxide dismutase from Sclerotinia sclerotiorum is involved with oxidative stress tolerance, virulence, and oxalate production Physiological and Molecular Plant Pathology, 2012,78:14-23. [本文引用: 2]
HIMELBLAUE, MIRAH, LIN SJ, CULOTTA VC, PE?ARRUBIAL, AMASINOR M . Identification of a functional homolog of the yeast copper homeostasis gene ATX1 from Arabidopsis Plant Physiology, 1998,117(4):1227-1234. [本文引用: 1]
SCHMIDT PJ, RAE TD, PUFAHL RA, HAMMAT, STRAINJ, O’HALLORANT V, CULOTTVV C . Multiple protein domains contribute to the action of the copper chaperone for superoxide dismutase The Journal of Biological Chemistry, 1999,274(34):23719-23725. [本文引用: 1]
LIY, CAO XL, ZHUY, YANG XM, ZHANG KN, XIAO ZY, WANGH, ZHAO JH, ZHANG LL, LI GB, ZHENG YP, FANJ, WANGJ, CHEN XQ, WU XJ, ZHAO JQ, DONG OX, CHEN XW, CHERNM, WANG WM . Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases New Phytologist, 2019,222(3):1507-1522. [本文引用: 1]
ZHOU YW . The role of chlorophyll synthetase for tocopherol synthesis in canola and Arabidopsis seeds [D]. Wuhan: Huazhong Agricultural University, 2017. (in Chinese) [本文引用: 1]
ZHANG LF, YANG ZX, ZHANG XW, LONG YX, ZHAOY . Cloning of the gene of pistil-regulated transcription factor in Capsella bursa-pastoris and its genetic transformation in Arabidopsis thaliana. Crop Research, 2018,32(3):202-207. (in Chinese) [本文引用: 1]
YUAN JH, DING YJ, YANG WJ, YAN BQ, CHAI YR, MEI JQ, QIANW . Identification of genes encoding secretory proteins related to the pathogenicity of Sclerotinia sclerotiorum using TRV-HIGS. Scientia Agricultura Sinica, 2019,52(23):4274-4284. (in Chinese) URL [本文引用: 1]
GUAN LM, SCANDALIOS JG . Hydrogen peroxide-mediated catalase gene expression in response to wounding Free Radical Biology and Medicine, 2000,28(8):1182-1190. [本文引用: 1]
CHEN XX, ZHANGT, YU QW, ZHOUW, AN YM, DU BH, GUO CH . Cloning and functional analysis of F-box protein gene MsFTL in alfalfa(Medicago sativa L.). Journal of Plant Genetic Resources, 2019,20(3):750-759. (in Chinese) [本文引用: 1]
WILLIAMSB, KABBAGEM, KIM HJ, BRITTR, DICKMAN MB . Tipping the balance:Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment PLoS Pathogens, 2011,7(6):e1002107. [本文引用: 1]
KABBAGEM, YARDENO, DICKMAN MB . Pathogenic attributes of Sclerotinia sclerotiorum: switching from a biotrophic to necrotrophic life style Plant Science, 2015,233:53-60. [本文引用: 1]
ZHANG MX, WANG QH, XUK, MENG YL, QUAN JL, SHAN WX . Production of dsRNA sequences in the host plant is not sufficient to initiate gene silencing in the colonizing oomycete pathogen Phytophthora parasitica PLoS ONE, 2011,6(11):e28114. [本文引用: 1]
WANGL, GEY, KANG YJ . Effect of copper on nuclear translocation of copper chaperone for superoxide dismutase-1 Experimental Biology and Medicine, 2016,241(14):1483-1488. [本文引用: 1]
ROLKEY, LIUS, QUIDDET, WILLIAMSONB, SCHOUTENA, WELTRING KM, SIEWERSV, TENBERGE KB, TUDZYNSKIB, TUDZYNSKIP . Functional analysis of H2O2-generating systems in Botrytis cinerea: The major Cu-Zn-superoxide dismutase (BCSOD1) contributes to virulence on French bean, whereas a glucose oxidase (BCGOD1) is dispensable Molecular Plant Pathology, 2004,5(1):17-27. [本文引用: 1]
 ,西南大学农学与生物科技学院,重庆 400715
,西南大学农学与生物科技学院,重庆 400715
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT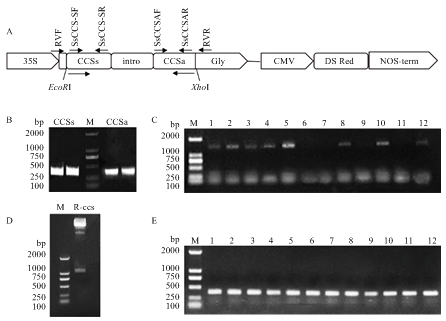 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT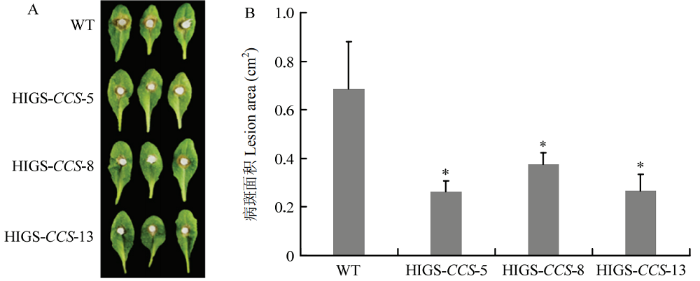 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT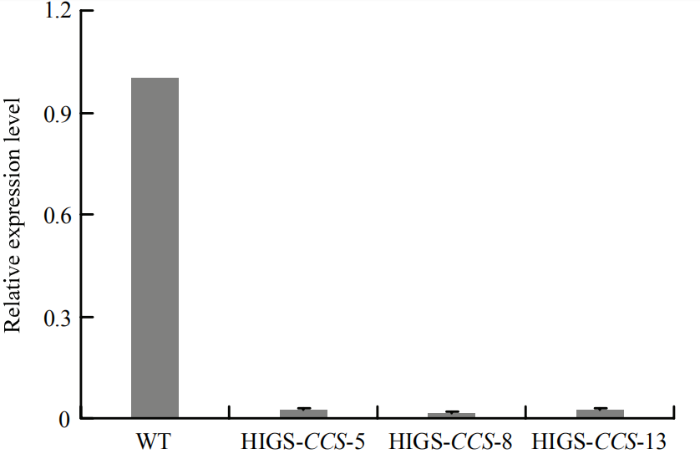 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}