Functional Characterization of a Maize Plasma Membrane Intrinsic Protein ZmPIP2;6 Responses to Osmotic, Salt and Drought Stress
ZHOU Lian, XIONG YuHan, HONG XiangDe, ZHOU Jing, LIU ChaoXian, WANG JiuGuang, WANG GuoQiang, CAI YiLin,Maize Research Institute, College of Agronomy and Biotechnology, Southwest University, Chongqing 400715
Abstract 【Objective】Plasma membrane Intrinsic Proteins (PIPs) exist widely in the membrane system of plant cells, which are essential to water transport and water balance in plant. The aim of this study is to explore the function of ZmPIP2;6 in plant water stress tolerance, and provide potential gene resources for new varieties of stress tolerance maize breeding. 【Method】Amino acid sequences of ZmPIP2;6 was analyzed and compared with other PIPs that involved in water stress tolerance. To verify the subcellular localization of ZmPIP2;6, ZmPIP2;6-GFP infusion vector was constructed and assessed using maize protoplasts isolated from leaves of maize seedlings. Tissues from root, stem, mature leaf, immature tassel, immature ear, endosperm and embryo of maize were isolated. Samples from root and leaf of maize were collected at different time after PEG or NaCl treatment. Total RNA were extracted, and expression pattern of ZmPIP2;6 in different tissues or under water stress condition was investigated by qRT-PCR. Transgenic Arabidopsis plants that overexpressed ZmPIP2;6 were generated and identified. The phenotype of ZmPIP2;6 overexpression transgenic Arabidopsis that tolerated to osmotic, salt or drought stress were monitored and primary root length and leaf water loss rate were measured. A number of stress responsive genes in ZmPIP2;6 overexpression Arabidopsis were detected under drought or salt condition.【Result】Analysis and comparison of the amino acid sequences showed that ZmPIP2;6 shared the same conserved structural domains and had a high degree of sequence similarity with other PIPs. The subcellular localization was assessed using maize protoplasts indicated ZmPIP2;6 was located on the plasma membrane. qRT-PCR result showed that ZmPIP2;6 was highly expressed in tassel. Treatment with PEG or NaCl resulted in induced expression of ZmPIP2;6 in root and leaf of maize. Overexpression of ZmPIP2;6 in transgenic Arabidopsis showed enhanced osmotic and salt stress tolerance in MS media plate and improved drought stress tolerance in soil condition compared to wild type. Expressions of related genes in the stress signaling pathway were changed in ZmPIP2;6 overexpression Arabidopsis under drought or salt condition. 【Conclusion】 Expression of ZmPIP2;6 was up-regulated under osmotic or salt stress condition. Overexpression of ZmPIP2;6 in Arabidopsis enhanced osmotic, salt and drought stress tolerance. A number of stress responsive genes in ZmPIP2;6 overexpression Arabidopsis were affected under salt or drought stress condition. These results indicated that ZmPIP2;6 might be involved in plant water stress responsive pathway. Keywords:maize (Zea mays L.);plasma membrane intrinsic proteins;osmotic stress;drought stress;salt stress
PDF (4900KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 周练, 熊雨涵, 洪祥德, 周京, 刘朝显, 王久光, 王国强, 蔡一林. 玉米质膜内在蛋白ZmPIP2;6响应渗透、盐和 干旱胁迫的功能鉴定[J]. 中国农业科学, 2020, 53(3): 461-473 doi:10.3864/j.issn.0578-1752.2020.03.001 ZHOU Lian, XIONG YuHan, HONG XiangDe, ZHOU Jing, LIU ChaoXian, WANG JiuGuang, WANG GuoQiang, CAI YiLin. Functional Characterization of a Maize Plasma Membrane Intrinsic Protein ZmPIP2;6 Responses to Osmotic, Salt and Drought Stress[J]. Scientia Acricultura Sinica, 2020, 53(3): 461-473 doi:10.3864/j.issn.0578-1752.2020.03.001
A:ZmPIP2;6在根、茎、成熟叶、未成熟雄穗、未成熟雌穗、胚和胚乳中的相对表达情况;B:ZmPIP2;6在含有10% PEG营养液处理后的三叶期玉米幼苗中的表达;C:ZmPIP2;6在含有100 mmol·L-1 NaCl的营养液处理后的三叶期玉米幼苗中的表达 Fig. 3Expression pattern of ZmPIP2;6 under normal and stress conditions
A: Relative expression levels of ZmPIP2;6 in root, stem, mature leaf, immature tassel, immature ear, endosperm and embryo; B: Trifoliolate maize seedlings were treated with 10% PEG in nutrient solution; C: Trifoliolate maize seedlings were treated with 100 mmol·L-1 NaCl in nutrient solution
A:野生型和ZmPIP2;6超表达转基因拟南芥植株在正常对照和干旱条件下生长14 d以及复水3 d后的表型。B:WT和ZmPIP2;6超表达转基因株系叶片水分散失率的检测 Fig. 6Phenotypes of ZmPIP2;6 overexpression transgenic lines under drought treatment and water loss rate
A. Phenotype of WT and ZmPIP2;6 overexpression transgenic Arabidopsis seedlings under normal and dehydration conditions for 14 days and re-watered for another 3 days. B. Determination of water loss from detached leaves of WT and ZmPIP2;6 overexpression transgenic lines
OBELL DB, ROBERTS MJ, SCHLENKERW, BRAUNN, LITTLE BB, REJESUS RM, HAMMER GL . Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest Science, 2014,344:516-519. [本文引用: 1]
YUC . China’s water crisis needs more than words Nature, 2011,470:307. [本文引用: 1]
KNEPPER MA . The aquaporin family of molecular water channels Proceedings of the National Academy of Sciences of the United States of America, 1994,91:6255-6258. [本文引用: 2]
TYERMAN SD, NIEMIETA CM, BRAMLEYH . Plant aquaporins: Multifunctional water and solute channels with expanding roles Plant, Cell & Environment, 2002,25:173-194. [本文引用: 2]
CHAUMONTF, BARRIEUF, WOJCIKE, CHRISPEELS MJ, JUNGR . Aquaporins constitute a large and highly divergent protein family in maize Plant Physiology, 2001,125:1206-1215. [本文引用: 3]
HACHEZC, BESSERERA, CHEVALIER AS, CHAUMONTF . Insights into plant plasma membrane aquaporin trafficking Trends in Plant Science, 2013,18:344-352. [本文引用: 1]
FETTERK, VAN WILDERV, MOSHELIONM, CHAUMONTF . Interactions between plasma membrane aquaporins modulate their water channel activity The Plant Cell, 2004,16:215-228. [本文引用: 2]
ZELAZNYE, BORST JW, MUYLAERTM, BATOKOH, HEMMINGA MA, CHAUMONTF . FRET imaging in living maize cells reveals that plasma membrane aquaporins interact to regulate their subcellular localization Proceedings of the National Academy of Sciences of the United States of America, 2007,104:12359-12364. [本文引用: 1]
VANDELEUR RK, MAYOG, SHELDEN MC, GILLIHAMM, KAISER BN, TYERMAN SD . The role of plasma membrane intrinsic protein aquaporins in water transport through roots: diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine Plant Physiology, 2009,149:445-460. [本文引用: 1]
ALLEVAK, MARQUEZM, VILLARREALN, MUTP, BUSTAMANTEC, BELLATIJ, MARTINEZG, CIVELLOM, AMODEOG . Cloning, functional characterization, and co-expression studies of a novel aquaporin (FaPIP2;1) of strawberry fruit Journal of Experimental Botany, 2010,61:3935-3945. [本文引用: 1]
CHENW, YINX, WANGL, TIANJ, YANGR, LIUD, YUZ, MAN, GAOJ . Involvement of rose aquaporin RhPIP1;1 in ethylene- regulated petal expansion through interaction with RhPIP2;1 Plant Molecular Biology, 2013,83:219-233. [本文引用: 1]
YANEFFA, SIGAUTL, MARQUEZM, ALLEVAK, PIETRASANTA LI, AMODEOG . Heteromerization of PIP aquaporins affects their intrinsic permeability Proceedings of the National Academy of Sciences of the United States of America, 2014,111:231-236. [本文引用: 1]
JOHANSONU, KARLSSONM, JOHANSSONI, GUSTAVSSONS, SJOVALLS, FRAYSSEL, WEIG AR, KJELLBOMP . The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiology, 2001,126:1358-1369. [本文引用: 1]
SAKURAIJ, ISHIKAWAF, YAMAGUCHIT, UEMURAM, MAESHIMAM . Identification of 33 rice aquaporin genes and analysis of their expression and function Plant & Cell Physiology, 2005,46:1568-1577. [本文引用: 1]
ZHANG DY, ALIZ, WANG CB, XUL, YI JX, XU ZL, LIU XQ, HE XL, HUANG YH, KHAN IA, TRETHOWAN RM, MA HX . Genome-wide sequence characterization and expression analysis of major intrinsic proteins in soybean (Glycine max L.). PLoS ONE, 2013,8:e56312. [本文引用: 1]
KOJIMAS, BOHNERA, VON WIRENN . Molecular mechanisms of urea transport in plants The Journal of Membrane Biology, 2006,212:83-91. [本文引用: 1]
FLEXASJ, RIBAS-CARBOM, HANSON DT, BOTAJ, OTTOB, CIFREJ, MCDOWELLN, MEDRANOH, KALDENHOFFR . Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo. The Plant Journal, 2006,48:427-439. [本文引用: 1]
HECKWOLFM, PATERD, HANSON DT, KALDENHOFFR . The Arabidopsis thaliana aquaporin AtPIP1;2 is a physiologically relevant CO(2) transport facilitator. The Plant Journal, 2011,67:795-804. [本文引用: 1]
ZHOUL, WANGC, LIUR, HANQ, VANDELEUR RK, DUJ, TYERMANS, SHOUH . Constitutive overexpression of soybean plasma membrane intrinsic protein GmPIP1;6 confers salt tolerance BMC Plant Biology, 2014,14:181. [本文引用: 5]
MA JF, TAMAIK, YAMAJIN, MITANIN, KONISHIS, KATSUHARAM, ISHIGUROM, MURATAY, YANOM . A silicon transporter in rice Nature, 2006,440:688-691. [本文引用: 1]
TAKANOJ, WADAM, LUDEWIGU, SCHAAFG, VON WIRENN, FUJIWARAT . The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. The Plant Cell, 2006,18:1498-1509. [本文引用: 1]
MA JF, YAMAJIN, MITANIN, TAMAIK, KONISHIS, FUJIWARAT, KATSUHARAM, YANOM . An efflux transporter of silicon in rice Nature, 2007,448:209-212. [本文引用: 1]
AFZALZ, HOWTON TC, SUNY, MUKHTAR MS . The roles of aquaporins in plant stress responses Journal of Developmental Biology, 2016,4:E9. [本文引用: 1]
AROCAR, PORCELR, RUIZ-LOZANOJ M. Regulation of root water uptake under abiotic stress conditions Journal of Experimental Botany, 2012,63:43-57. [本文引用: 2]
ZHUC, SCHRAUTD, HARTUNGW, SCHAFFNER AR . Differential responses of maize MIP genes to salt stress and ABA Journal of Experimental Botany, 2005,56:2971-2981. [本文引用: 2]
WANG WF, ZHANG SQ, CHAI YL, LIU JB . Transcription regulation of ZmPIPs in defferent young maize root zones responses to short term in different young maize root water stress and rewater Acta Botanica Boreali-Occidentalia Sinica, 2010,30:1959-1965. (in Chinese) [本文引用: 2]
ZHOUS, HUW, DENGX, MAZ, CHENL, HUANGC, WANGC, WANGJ, HEY, YANGG, HEG . Overexpression of the wheat aquaporin gene, TaAQP7, enhances drought tolerance in transgenic tobacco PLoS ONE, 2012,7:e52439. [本文引用: 2]
LIUC, FUKUMOTOT, MATSUMOTOT, GENAP, FRASCARIAD, KANEKOT, KATSUHARAM, ZHONGS, SUNX, ZHUY, IWASAKII, DINGX, CALAMITAG, KITAGAWAY . Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination Plant Physiology and Biochemistry, 2013,63:151-158.
HUW, YUANQ, WANGY, CAIR, DENGX, WANGJ, ZHOUS, CHENM, CHENL, HUANGC, MAZ, YANGG, HEG . Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco Plant & Cell Physiology, 2012,53:2127-2141.
SREEDHARANS, SHEKHAWAT UK, GANAPATHI TR . Transgenic banana plants overexpressing a native plasma membrane aquaporin MusaPIP1;2 display high tolerance levels to different abiotic stresses Plant Biotechnol Journal, 2013,11:942-952. [本文引用: 2]
ALAVILLIH, AWASTHI JP, ROUT GR, SAHOOL, LEE BH, PANDA SK . Overexpression of a barley aquaporin gene, HvPIP2;5 confers salt and osmotic stress tolerance in yeast and plants Frontiers in Plant Science, 2016,7:1566. [本文引用: 3]
ZHOUL, ZHOUJ, XIONGY, LIUC, WANGJ, WANGG, CAIY . Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS ONE, 2018,13:e0198639. [本文引用: 2]
MARTINEZ-BALLESTA MDELC, BASTIASE, ZHUC, SCHAFFNER AR, GONZALEZ-MOROB, GONZALEZ-MURUAC, CARVAJALM . Boric acid and salinity effects on maize roots. Response of aquaporins ZmPIP1 and ZmPIP2, and plasma membrane H +-ATPase, in relation to water and nutrient uptake Physiology Plant, 2008,132:479-490. [本文引用: 1]
NELSON BK, CAIX, NEBENFUHRA . A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. The Plant Journal, 2007,51:1126-1136. [本文引用: 1]
BARTR, CHERNM, PARK CJ, BARTLEYL, RONALD PC . A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts Plant Methods, 2006,2:13. [本文引用: 1]
CHAUMONTF, BARRIEUF, HERMAN EM, CHRISPEELS MJ . Characterization of a maize tonoplast aquaporin expressed in zones of cell division and elongation Plant Physiology, 1998,117:1143-1152. [本文引用: 1]
BOURSIACY, CHENS, LUU DT, SORIEULM, VAN DENDRIES N, MAURELC . Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiology, 2005,139:790-805. [本文引用: 1]
MARTINEZ-BALLESTA MC, APARICIOF, PALLASV, MARTINEZV, CARVAJALM . Influence of saline stress on root hydraulic conductance and PIP expression in Arabidopsis. Journal of Plant Physiology, 2003,160:689-697. [本文引用: 1]
MARULANDAA, AZCONR, CHAUMONTF, RUIZ-LOZANOJ M, AROCAR. Regulation of plasma membrane aquaporins by inoculation with a Bacillus megaterium strain in maize (Zea mays L.) plants under unstressed and salt-stressed conditions. Planta, 2010,232:533-543. [本文引用: 1]
MURIESB, FAIZEM, CARVAJALM , MARTINEZ-BALLESTA MDEL C. Identification and differential induction of the expression of aquaporins by salinity in broccoli plants Molecular BioSystems, 2011,7:1322-1335. [本文引用: 1]
SADEN, GEBRETSADIKM, SELIGMANNR, SCHWARTZA, WALLACHR, MOSHELIONM . The role of tobacco Aquaporin1 in improving water use efficiency, hydraulic conductivity, and yield production under salt stress Plant Physiology, 2010,152:245-254. [本文引用: 1]
AHARONR, SHAHAKY, WININGERS, BENDOVR, KAPULNIKY, GALILIG . Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress The Plant Cell, 2003,15:439-447. [本文引用: 2]
KATSUHARAM, KOSHIOK, SHIBASAKAM, HAYASHIY, HAYAKAWAT, KASAMOK . Over-expression of a barley aquaporin increased the shoot/root ratio and raised salt sensitivity in transgenic rice plants Plant & Cell Physiology, 2003,44:1378-1383. [本文引用: 2]
YAMAGUCHI-SHINOZAKIK, SHINOZAKIK . A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. The Plant Cell, 1994,6:251-264. [本文引用: 1]
SHIH, ISHITANIM, KIMC, ZHU JK . The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na +/H + antiporter. Proceedings of the National Academy of Sciences of the United States of America, 2000,97:6896-6901. [本文引用: 1]
LIUJ, ZHU JK . A calcium sensor homolog required for plant salt tolerance Science, 1998,280:1943-1945. [本文引用: 1]
HALFTERU, ISHITANIM, ZHU JK . The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium- inding protein SOS3. Proceedings of the National Academy of Sciences of the United States of America, 2000,97:3735-3740. [本文引用: 1]
QIU QS, GUOY, DIETRICH MA, SCHUMAKER KS, ZHU JK . Regulation of SOS1, a plasma membrane Na +/H + exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proceedings of the National Academy of Sciences of the United States of America, 2002,99:8436-8441. [本文引用: 1]
APSE MP, AHARON GS, SNEDDEN WA, BLUMWALDE . Salt tolerance conferred by overexpression of a vacuolar Na +/H + antiport in Arabidopsis. Science, 1999,285:1256-1258. [本文引用: 2]
QIU QS, GUOY, QUINTERO FJ, PARDO JM, SCHUMAKER KS, ZHU JK . Regulation of vacuolar Na +/H + exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. The Journal of Biological Chemistry, 2004,279:207-215. [本文引用: 1]
SHIH, LEE BH, WU SJ, ZHU JK . Overexpression of a plasma membrane Na +/H + antiporter gene improves salt tolerance in Arabidopsis thaliana. Nature Biotechnology, 2003,21:81-85. [本文引用: 1]
YANGQ, CHEN ZZ, ZHOU XF, YIN HB, LIX, XIN XF, HONG XH, ZHU JK, GONGZ . Overexpression of SOS (Salt Overly Sensitive) genes increases salt tolerance in transgenic Arabidopsis. Molecular Plant, 2009,2:22-31. [本文引用: 1]
ZHUOC, WANGT, GUOZ, LUS . Overexpression of MfPIP2-7 from Medicago falcata promotes cold tolerance and growth under NO3 (-) deficiency in transgenic tobacco plants. BMC Plant Biology, 2016,16:138. [本文引用: 1]
 ,西南大学农学与生物科技学院玉米研究所,重庆400715
,西南大学农学与生物科技学院玉米研究所,重庆400715
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT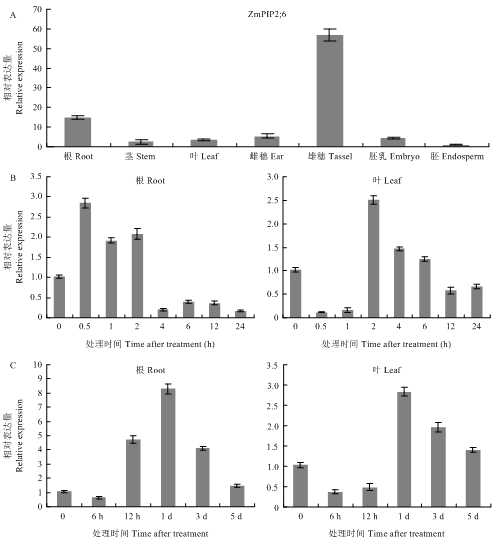 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}