Physiological and Biochemical Characteristics of Low Temperature Vernalization of Germinating Seeds of Brassica rapa
XU ChunMei, ZOU Ya, LIU ZiGang,, MI WenBo, XU MingXia, DONG XiaoYun, CAO XiaoDong, ZHENG GuoQiang, FANG XinLingAgronomy College, Gansu Agricultural University/Gansu Rapeseed Engineering and Technology Research Center/Key Laboratory of Arid Land Crop Science in Gansu Province/Gansu Key Laboratory of Crop Improvement and Germplasm Enhancement, Lanzhou 730070
Abstract 【Objective】 To explore the possibility of low temperature through vernalization to the germination of Brassica rapa, and the phenotypic changes of seed physiology and biochemistry and plant set during vernalization, so as to provide theoretical basis for artificial breeding and accelerating breeding process of B. rapa. 【Method】 Three different temperature-sensitive of B. rapa were used as materials, the germinated seeds were placed in 4℃ of low temperature for vernalization treatment. During the vernalization process (0 d, 20 d, 30 d, 40 d, 50 d, 60 d) the physiological and biochemical indexes such as nitrate reductase, antioxidant enzyme activity, osmotic regulator and malondialdehyde content of the germinated seeds were measured. Simultaneously germinated seeds of each vernalization treatment, the growth period of the seed formation plant was observed, and the seed-setting performance of the plants were measured. 【Result】 With the increase of vernalization time, the vernalization rate (V), plant height (FPH), mature plant height (MPH) and primary branch number (PB), the number of pods per plant (SP), the length of pods (LS), the number of pods (SS) and the yield per plant (YP) of germinated seeds of B. rapa showed an increasing trend; the early stage of vernalization (0-40 d), the plant's seed-setting performance showed significant differences among different varieties. After the vernalization time increased (50-60 d), the seed-setting ability of different varieties was slightly difference, but they were not significantly difference. The results of regression analysis showed that the germination seeds of strong winter rapa Longyou 7 at 4℃ were completely vernalized (springing rate >95%) and need to be treated 76.9 d, Longyou 9 and Tianyou 4 were 54.0 d, 39.4 d, respectively. Correlation analysis showed that the vernalization rate was extremely significant positive correlated with plant height and seed-setting performance and other phenotypic traits. The correlation coefficient between vernalization rate with average first flower plant height and average mature plant height was 0.947 and 0.985, which indicated that vernalization degree of winter B. rapa significantly affected plant height and seed-setting performance. With the increase of low temperature vernalization time, the germinated seeds of winter B. rapa with nitrate reductase (NR), superoxide dismutase (SOD), peroxidase (POD), malondialdehyde (MDA), soluble protein (SP), Soluble sugar (SS) were increased first and then decreased, and the activity of catalase (CAT) decreased continuously. Compared with the control (the germinated seeds not treated at low temperature), the content of GA3 in the germinated seeds of Longyou 7 and Longyou 9 decreased significantly at the low temperature vernalization treatment, and the content of GA3 in the germinated seeds of Tianyou 4 was significantly higher than that of the control at 30 days. Compared with the control, the content of IAA in the winter rapeseed germinated in vernalization increased significantly (except for the treatment 40 days of Longyou9). Among them, the content of IAA in the seeds treated with vernalization for 50 days of Tianyou 4 increased by 197.0% compared with the control. The ABA content of the Longyou 7 was significantly increased compared with the control. 【Conclusion】 The germination seeds of winter Brassica rapa can be perceived the low temperature to complete the vernalization. The low temperature time required for vernalization of the variety depends on the winter strength. During the process vernalization of low temperature, the physiological and biochemical status is occurring some changes of the seed of winter B. rapa, and ultimately affect the growth and development of the plant and its seed setting performance. Keywords:Brassica rapa;germination seed;phenotypic characteristics;physiological and biochemical
PDF (1115KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 徐春梅, 邹娅, 刘自刚, 米文博, 徐明霞, 董小云, 曹小东, 郑国强, 方新玲. 白菜型冬油菜萌动种子低温春化的生理生化特征[J]. 中国农业科学, 2020, 53(5): 929-941 doi:10.3864/j.issn.0578-1752.2020.05.006 XU ChunMei, ZOU Ya, LIU ZiGang, MI WenBo, XU MingXia, DONG XiaoYun, CAO XiaoDong, ZHENG GuoQiang, FANG XinLing. Physiological and Biochemical Characteristics of Low Temperature Vernalization of Germinating Seeds of Brassica rapa[J]. Scientia Acricultura Sinica, 2020, 53(5): 929-941 doi:10.3864/j.issn.0578-1752.2020.05.006
a、b、c:春化处理30 d后移栽营养钵生长的植株;d、e、f:春化处理60 d后移栽营养钵生长的植株;1:对照(CK);2、3:春化 Fig. 1Effect of vernalization time on phenotype of B. Rapa
a, b, and c: Plants grown in a nutrient bowl after 30 days of vernalization; d, e, and f: Plants grown in a nutrient bowl after 60 days of vernalization; 1: CK; 2, 3: Vernalization
Table 2 表2 表2不同春化时间对白菜型冬油菜春化率的影响 Table 2Effect of vernalization time on the number of flowering plants of B. rapa (%)
时间Time (d)
陇油7号Longyou7
陇油9号Longyou9
天油4号Tianyou4
0(CK )
0.00
0.00
0.00
20
25.81
27.27
56.25
30
19.23
56.52
93.10
40
38.24
73.68
97.06
50
60.00
95.00
96.15
60
86.96
100.00
96.88
线性拟合方程Linear fitting equation
y =1.3567x-6.85(R2=0.8901)
y = 1.7954x-1.1007(R2=0.9785)
y =1.6169x+19.344(R2=0.7924)
春化率95%所需低温处理天数TTT (d)
76.9
54.0
39.4
TTT: The number of low temperature treatment days required for the vernalization rate of 95%. y is the vernalization rate in the table; x is the time required for the low temperature vernalization when the vernalization rate reaches y TTT:春化率95%所需低温处理天数。表中y为春化率;x为春化率达到y时所需低温春化的时间
Table 4 表4 表4不同春化时间对白菜型冬油菜结实性的影响 Table 4Effect of vernalization time on seed set of B. rapa
品种Variety
时间Time(d)
陇油7号Longyou7
陇油9号Longyou9
天油4号Tianyou4
PB
20
1.00±1.00b
2.00±1.00a
1.67±0.58b
30
0.67±0.58b
2.33±0.58a
2.00±1.00b
40
1.67±0.58ab
1.67±1.15a
1.33±0.58b
50
2.00±0ab
2.67±0.58a
2.33±0.58ab
60
3.00±1.00a
2.67±0.58a
3.33±0.58a
SP
20
37.00±8.19bc
40.3±4.73b
38.00±5.57b
30
30.33±3.21c
42.67±6.81b
42.67±4.04b
40
45.0±10.54abc
49.00±7.00ab
49.67±5.51ab
50
53.33±10.07ab
57.33±9.71a
53.67±13.05ab
60
57.33±10.07a
59.33±6.66a
60.67±10.60a
LS
20
4.38±0.47b
4.39±1.06b
4.97±0.46c
30
4.47±0.26b
3.83±0.26b
5.10±0.53bc
40
4.64±0.30b
4.83±0.58b
5.56±1.09abc
50
7.03±0.38a
6.34±.53a
6.20±0.45ab
60
6.91±0.25a
6.42±0.52a
6.66±0.21a
SS (mm)
20
12.33±3.48b
12.67±3.18b
14.11±2.14b
30
14.78±2.22b
12.78±9.22b
16.67±4.63a
40
16.89±3.66a
15.22±7.50a
19.44±3.37a
50
21.78±2.50a
24.22±4.17a
22.33±6.77a
60
21.33±1.15a
24.56±1.26a
23.33±0.58a
YP (g)
20
0.74±0.10b
0.78±0.18c
0.75±0.10b
30
0.67±0.09b
0.69±0.08c
0.92±0.13b
40
0.91±0.18b
0.97±0.17bc
1.21±0.09a
50
1.22±0.10a
1.14±0.22ab
1.25±0.24a
60
1.28±0.18a
1.32±0.17a
1.27±0.07a
PB: Number of primary branches; SP: Number of siliques per plant; LS: Lengths per silique; SS: Seeds per silique; YP: Yield per plant. Different lowercase letters indicate significant differences at the 0.05 level. The same as below PB:一次分枝数;SP:单株角果数;LS:角果长度;SS:角果粒数;YP:单株产量。不同小写字母表示在0.05水平存在显著差异。下同
Table 5 表5 表5不同冬性白菜型油菜春化率、表型特征之间的相关性 Table 5Correlation between vernalization rate and phenotypic traits of B. rapa
性状 Trait
春化率V
初花株期高FPH
成熟期株高MPH
一次分枝数PB
单株角果数SP
角果长度LS
角果粒数SS
单株产量YP
春化率V
1
初花期株高FPH
0.947**
1
成熟期株高MPH
0.985**
0.978**
1
一次分枝数PB
0.841**
0.862**
0.847**
1
单株角果数SP
0.876**
0.837**
0.854**
0.915**
1
角果长度LS
0.839**
0.798**
0.820**
0.867**
0.974**
1
角果粒数SS
0.881**
0.854**
0.877**
0.874**
0.972**
0.978**
1
单株产量YP
0.893**
0.858**
0.882**
0.870**
0.982**
0.979**
0.982**
1
V: Vernalize rate. **: Significantly correlated at 0.01 level (both sides); *: Significantly correlated at 0.05 level (both sides). The same as below V:春化率。**:在0.01水平(双侧)差异极显著;*:在0.05水平(双侧)差异显著。下同
ZHANG TG, LIY, DIAO ZH, SHI ZF, WANGJ, ZHENGS . Cloning and expression analysis of RbohC and RbohF genes in Brassica rapa L. Acta Botanica Boreali-Occidentalia Sinica, 2018,38(10):7-16. (in Chinese) [本文引用: 1]
SONGY, DOU LD, ZHANG HJ . Molecular and genetic mechanisms of control of floral induction in higher plants Plant Physiology Journal, 2014,50(10):1459-1468. (in Chinese) [本文引用: 1]
YU RF, SU TB, YU SC, ZHANG FL, YU YJ, ZHANG DS, ZHAO XY, WANG WH, LU GX . Effects of plumule-vernalization and seeding-vernalization on bolting and flowering times of different Chinese cabbage varieties China Vegetables, 2016(5):27-32. (in Chinese) [本文引用: 1]
YANG DG, SUN YX, IRFAN AR, LIU XM, LV JY, YU JP, GONGL, LIUZ, BAIB . Effects of low temperature stress on germination and physiological of maize seedling Journal of Northeast Agricultural University, 2018,49(5):4-11. (in Chinese) [本文引用: 1]
ZHANGX . Analysis of the vernalization process effect on DNA methylation and protein level in Brassica napus L. [D]. Zhengzhou: Zhengzhou University, 2013. ( in Chinese) [本文引用: 1]
ZHAOK . Effect of low temperature treated wheat germinaing seeds on growth period and yield under early sowing condition [D]. Yaan: Sichuan Agricultural University, 2017. ( in Chinese) [本文引用: 1]
JIANG XM, YU XH . Stimulatory effects of low temperature treatment of germinating seeds on flowerbud differentiation in Broccoli Journal of Plant Physiology and Molecular Biology, 2004(4):421-427. (in Chinese) [本文引用: 1]
TURANO, EKMEKCIY . Activities of photosystem II and antioxidant enzymesin chickpea (Cicer arietinum L.) cultivars exposed to chilling temperatures. Acta Physiology Plant, 2011,33:67-78. [本文引用: 1]
LI YQ, YAO ZP, ZHANGH, DAI PH . Effects of low temperature stress on physiological-biochemical indexes of three early spring ephemeral plants Xinjiang Agricultural Sciences, 2012,49(9):1608-1615. (in Chinese) [本文引用: 1]
CHENQ, YUAN JH, SUN WC, LIU ZG, ZHAO XW, FANGY, WU JY, LI XC, ZENG XC, MIC, PU YY, MAL, ZHAO YN, FANGY, XU YZ . Leaf photo synthetics and endogenous hormones of spring and winter Rapa ( Brassica rapa L.) under low temperature. Chinese Journal of Oil Crop Sciences, 2017,39(1):37-46. (in Chinese) [本文引用: 1]
YANGG, SUN WC, WANG LP, ZENG XC, LIU ZG, LI XC, FANGY, WU JY, WANG KY, HOU XF, QIANW, MAL, LIU HQ . equence variation of vernalization gene BrFLC in turnip rapes (Brassica rapa L.) Chinese Journal of Oil Crop Sciences, 2015,37(2):167-172. (in Chinese) [本文引用: 1]
YANG NN . Study on cold resistance mechanism and selection index evaluation system of winter tumip rape (Brassica napus L.) [D]. Lanzhou: Gansu Agricultural University, 2014. ( in Chinese) [本文引用: 1]
CHENK, LIUJ, ZHOU XT, JI RF, TONG YR, CHENT, YUANY . Determination of various endogenous hormones in flower buds of panax ginseng by UPLC-MS/MS Modern Chinese Medicine, 2018,20(6):77-82. (in Chinese) [本文引用: 1]
ESIMN, TIRYAKID, KARADAGOGLUO, ATICIO . Toxic effects of boron on growth and antioxidant system parameters of maize (Zea mays L.) roots. Toxicology and Industrial Health, 2013,29(9):800-805. [本文引用: 1]
HUW, HOU XL, SHI GJ . Vernalization characteristics and mechanism of plant vernalization Chinese Bulletin of Botany, 2004,21(1):26-36. (in Chinese) [本文引用: 1]
YANG GL, ZHANG XQ, XU CH, LA BC J,. Correlation analysis between economic characteristics and yield of rape Nianhe 18 Journal of Zhejiang Agricultural Sciences, 2019,60(2):186-188. (in Chinese) [本文引用: 1]
TIANH, DUAN MY, WANGL . Research progress of nitrate reductase function in plants Chinese Agricultural Science Bulletin, 2009,25(10):96-99. (in Chinese) [本文引用: 1]
GONG SZ . The influence of on nitric reductase active in maize seedling stage in low temperature Journal of Maize Sciences, 2003,11(2):73-74. (in Chinese) [本文引用: 1]
YAN LH, DAI HF, HEJ . The research on POD isozyme activity change of wheat after treatmention of vernalization devernalization or cold acclimation Chinese Agricultural Science Bulletin, 2014,30(9):99-104. (in Chinese) [本文引用: 1]
KAR RK, CHOUDHURI MA . Possible mechanisms of lighter induced chlorophyll degradation in senescing leaves of Hydrilla verticillata. Physiologia Plantarum, 1987,70(4):729-734. [本文引用: 1]
LI QY, WANG SS, ZHOU PL, LIU GL, ZENG WL, ZHOU DJ, ZHENGX, CHEN AG . Effects of low temperature stress on leaf shape and physiological characteristics in tobacco seedlings Chinese Tobacco Science, 2018,39(1):17-23. (in Chinese) [本文引用: 1]
HUANG WC, FAN YB, WANG YC . Low temperature stress and maize seedlings: Effect on antioxidant enzyme system and osmotic regulation Chinese Agricultural Science Bulletin, 2018,34(24):6-12. (in Chinese) [本文引用: 1]
YU LL, YUANX, SHAOQ, CHENX, CUI XY, WANG XJ . Effects of vernalization duration on activity of protective enzyme and content of soluble protein in leaves of peas Fujian Journal of Agricultural Sciences, 2016,31(5):460-464. (in Chinese) [本文引用: 1]
CHANG BW, ZHONGP, LIUJ, TANG ZH, GAO YB, YU HJ, GUOW . Effects of low temperature stress and gibberellin on seed germination and seeding physiological responses in peanut seedlings Acta Agronomica Sinica, 2019,45(1):118-130. (in Chinese) [本文引用: 1]
LI CY, XUW, LIU LW, YANGJ, ZHU XK, GUO WS . Changes of endogenous hormone contents and antioxidative enzyme activities in wheat leaves under low temperature stress at jointing stage Chinese Journal of Applied Ecology, 2015,26(7):2015-2022. [本文引用: 1]
ZHOU BY, LI YB, CHEN JZ, JI ZL, HU ZQ . Effects of low temperature stress and ABA on flower formation and endogenous hormone of litchi Acta Horticulturae Sinica, 2002,29(6):577-578. (in Chinese) [本文引用: 1]
WANGX, YUJ, YANGY, CANGJ, LI ZF . Changes of endogenous hormones of winter wheat varieties with different cold-resistances under low temperature Journal of Triticeae Crops, 2009,29(5):827-831. (in Chinese) [本文引用: 1]
 ,, 米文博, 徐明霞, 董小云, 曹小东, 郑国强, 方新玲甘肃农业大学农学院/甘肃省油菜工程与技术研究中心/甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室, 兰州 730070
,, 米文博, 徐明霞, 董小云, 曹小东, 郑国强, 方新玲甘肃农业大学农学院/甘肃省油菜工程与技术研究中心/甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室, 兰州 730070
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT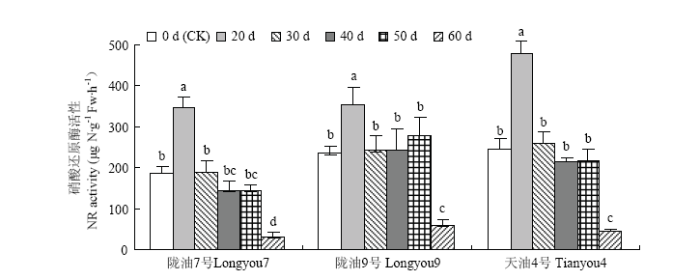 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT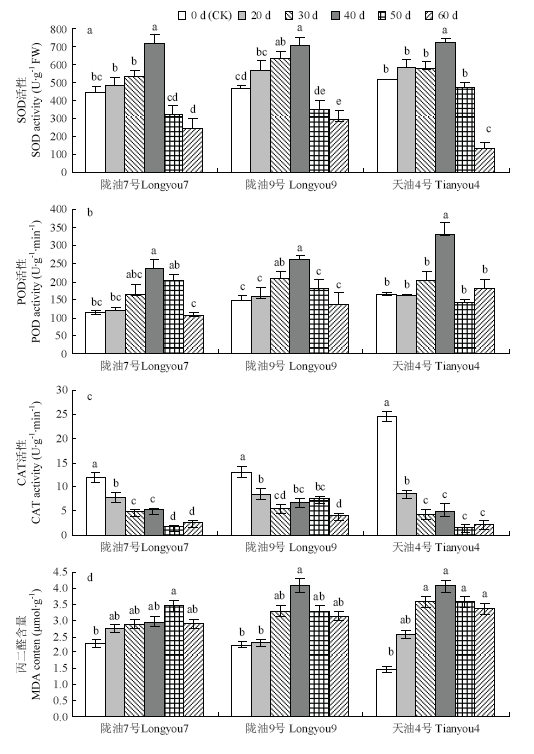 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT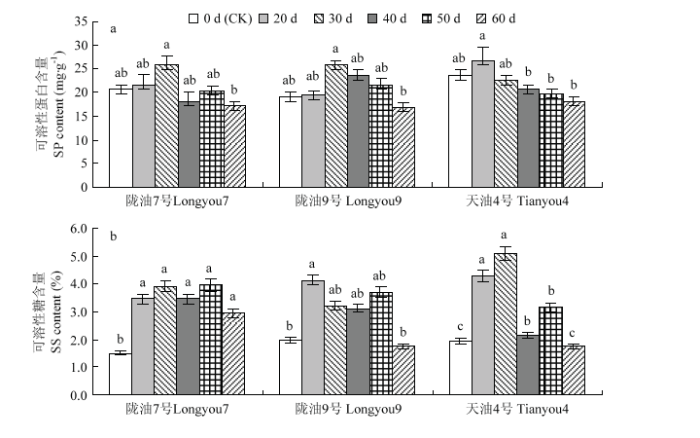 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT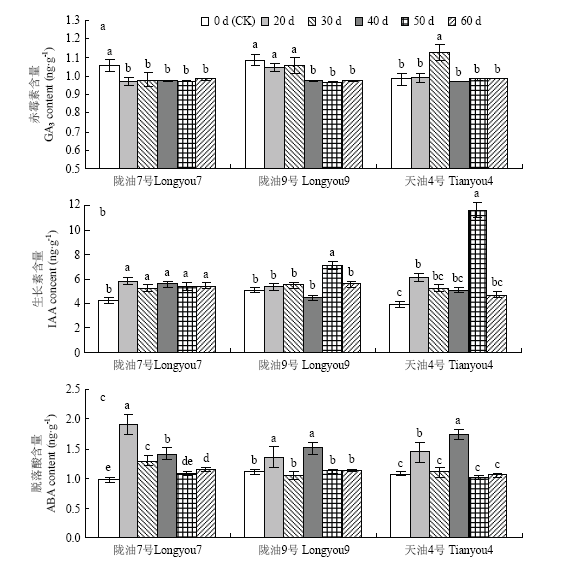 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}