Genetic Diversity and Phylogenetics of Malus baccata (L.) Borkh Revealed by Chloroplast DNA Variation
GAO Yuan, WANG DaJiang, WANG Kun,, CONG PeiHua,, ZHANG CaiXia, LI LianWen, PIAO JiChengResearch Institute of Pomology, Chinese Academy of Agricultural Sciences/Key Laboratory of Horticultural Crops Germplasm Resources Utilization, Ministry of Agriculture, Xingcheng 125100, Liaoning
Abstract 【Objective】Malus baccata (L.) Borkh is the most widely distributed native species in China. The non-coding region of chloroplast genome by maternal inheritance is suitable for the systematic study of lower taxonomic levels (such as families and genera). The non-coding regions of cpDNA of 215 germplasms were sequenced, and their genetic variation was analyzed. In this study, the genetic diversity of Malus baccata and the phylogenetic relationship among different populations were explored from the perspective of maternal inheritance, which provided a theoretical basis for origin and genetic evolution, collection and protection of Malus baccata germplasm resources in China.【Method】Four non-coding regions (trnH-psbA, trnS-trnG spacer + intron, trnT-5'trnL and 5'trnL-trnF) of 215 germplasms were amplified by four primers. After manually proofreading sequences obtained through forward and backward sequencing, MEGA 7.0 was used for sequence splicing and alignment, and based on genetic distance, the Neighbour-Joining phylogenetic tree was constructed among different populations of Malus baccata. DnaSP ver5.10.01 was used to calculate the genetic diversity parameters of chloroplast DNA, gene flow and gene differentiation among different populations. Arlequin v3.5 was used to analyze standard molecular variation (AMOVA), and NetWork 4.6.1.2 was used to construct Median-Joining network for cpDNA haplotypes among intraspecific populations of Malus baccata.【Result】The length of four non-coding regions of chloroplast DNA was 3 777 bp after sequencing, splicing, alignment and merging, and 171 variable sites were detected, which included 1 singleton variable sites, 20 parsimony informative sites and 150 insertion-deletion gaps. Among 215 accessions of Malus baccata, the number of variable sites of region trnH-psbA, trnS-trnG spacer + intron, trnT-5'trnL and 5'trnL-trnF were 26, 32, 103 and 10, respectively. The number of haplotypes for four regions were 8, 8, 6 and 4, respectively, and after four regions merged, the haplotypes of chloroplast DNA fragments were 24. The region with highest nucleotide diversity was trnT-5'trnL (Pi=0.01174), and the region with highest haplotype diversity was trnS-trnG spacer + intron (Hd=0.599), and the haplotype diversity of 5'trnL-trnF was the lowest (Hd=0.228). The cpDNA diversity of Malus baccata was high (Hd=0.727, Pi=0.00577). Tajima’s test showed all Tajima’s D values were not statistical at different levels, which indicated that variation of those chloroplast regions followed natural theory of molecular evolution. AMOVA showed that genetic variation mainly existed within populations.【Conclusion】The four non-coding regions of chloroplast DNA were suitable for the analysis of genetic diversity and phylogenetics of Malus baccata. At the cpDNA level, it was not natural selection, but mutation pressure and genetic drift that led to population evolution of Malus baccata. The genetic differentiation among populations was not completely correlated with their geographical distance. Malus baccata might originate from several sites, and the three possible origins, including Heilongjiang and Jilin, Inner Mongolia, Gansu and Shanxi, were inferred. Keywords:Malus baccata;chloroplast DNA;non-coding region;genetic diversity;phylogenetics
PDF (698KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 高源, 王大江, 王昆, 丛佩华, 张彩霞, 李连文, 朴继成. 基于叶绿体DNA变异的山荆子种质遗传多样性和系统演化[J]. 中国农业科学, 2020, 53(3): 600-611 doi:10.3864/j.issn.0578-1752.2020.03.012 GAO Yuan, WANG DaJiang, WANG Kun, CONG PeiHua, ZHANG CaiXia, LI LianWen, PIAO JiCheng. Genetic Diversity and Phylogenetics of Malus baccata (L.) Borkh Revealed by Chloroplast DNA Variation[J]. Scientia Acricultura Sinica, 2020, 53(3): 600-611 doi:10.3864/j.issn.0578-1752.2020.03.012
WANG DJ, WANGK, GAOY, ZHAO JR, LIU LJ, GONGX, LI LW . Preliminary investigation of modern distribution of Malus resources in China. Journal of Plant Genetic Resources, 2017,18(6):1116-1124. (in Chinese) [本文引用: 1]
DU XM, YANG TZ, GAO JD, WANGQ, CAI HC, LI CY, GONG GH . Wild Malus baccata(L.) Borkh. in China: Natural distribution, utilization and study status. Chinese Agricultural Science Bulletin, 2017,33(5):24-28. (in Chinese) [本文引用: 1]
ZOUX, PENGY, WANGL, LIY, ZHANG WX, LIUX . Impact of climate change on the distribution pattern of Malus baccata(L.) Borkh. in China since the Last Glacial Maximum. Plant Science Journal, 2018,36(5):676-686. (in Chinese) [本文引用: 2]
WANG LH, TANG GG, XIA HW, CAIH . Study on leaf venation of Malus baccata(L.) Borkh. Journal of Nanjing Forestry University (Natural Sciences Edition), 2008,32(2):39-42. (in Chinese) [本文引用: 1]
CHENX, TANG GG, ZHENG YH, WANG LH . RAPD analysis of genetic diversity of Malus baccata Acta Botanica Boreali Occidentalia Sinica, 2008,28(10):1954-1959. (in Chinese) [本文引用: 2]
ROBINSON JP, HARRIS SA, JUNIPER BE . Taxonomy of the genus Malus Mill.(Rosaceae ) with emphasis on the cultivated apple, Malus domestica Borkh. Plant Systematic and Evolution, 2001,226(1/2):35-58. [本文引用: 1]
WANG LH, YANG JX, ZHENG YH, TANG GG . Modelling the geographical distribution of Malus baccata. Journal of Beijing Forestry University, 2011,33(3):70-73. (in Chinese) [本文引用: 1]
YANGF, LIUZ, YIK, LIU YJ, WANGQ, SUN JS . Studies on geographical regions and analysis on genetic diversity of phenotypic of natural population of ‘Malus baccata ( L.) Borkh’ in Northeast of China. Journal of Plant Genetic Resources, 2015,16(3):490-496. (in Chinese) [本文引用: 1]
WANG LH, ZHENG YH, TANG GG . Analyses of genetic diversity and genetic relationship of eight populations of Malus baccata based on SSR marker. Journal of Plant Resources and Environment, 2012,21(1):42-46. (in Chinese) [本文引用: 2]
GAOY, WANGK, WANG DJ, ZHAO JR, ZHANG CX, CONG PH, LIU LJ, LI LW, PIAO JC . The genetic diversity and population structure analysis on Malus baccata(L.) Borkh. from 7 sources. Scientia Agricultura Sinica, 2018,51(19):3766-3777. (in Chinese) [本文引用: 4]
NI LH, ZHAO ZL, MIM . Advances in research on chloroplast genome of medicinal plants Journal of Chinese Medicinal Materials, 2015,38(9):1990-1994. (in Chinese) [本文引用: 2]
ZHANG YJ, ZHANG DZ . Advances in phylogenomics based on complete chloroplast genomes Plant Diversity and Resources, 2011,33(4):365-375. (in Chinese) [本文引用: 1]
FUT, WANG ZL, QIAN PX, LIW, YUAN DM, YAN CF . The latest research progress and application of the DNA barcode in higher plants Journal of Nuclear Agricultural Sciences, 2016,30(5):887-896. (in Chinese) [本文引用: 1]
SAVOLAINENV, CORBAZR, MONCOUSINC, SPICHIPERR, MANEN JF . Chloroplast DNA variation and parentage analysis in 55 apples Theoretical and Applied Genetics, 1995,90:1138-1141. [本文引用: 1]
ROBINSON JP, HARRIS SA, JUNIPER BE . Taxonomy of the genus Malus Mill. (Rosaceae) with emphasis on the cultivated apple, Malus domestica Borkh. Plant Systematics and Evolution, 2001,226:35-58. [本文引用: 1]
COARTE, VAN GLABEKES, DE LOOSEM, LARSEN AS, ROLDAN-RUIZI . Chloroplast diversity in the genus Malus: new insights into the relationship between the European wild apple (Malus sylvestris (L) Mill.) and the domesticated apple (Malus domestica Borkh.). Molecular Ecology, 2006,15:2171-2182. [本文引用: 1]
NIKIFOROVA SV, CAVALIERID, VELASCOR, GOREMYKINV . Phylogenetic analysis of 47 chloroplast genomes clarifies the contribution of wild species to the domesticated apple maternal line Molecular Biology Evolution, 2013,30(8):1751-1760. [本文引用: 1]
DING FB . Sequence analysis of matK and ITS between Malus hupehensis and its relative species [D]. Nanjing: Nanjing Forestry University, 2012. ( in Chinese) [本文引用: 1]
ZHU YD, CAO MG, XUZ, WANGK, ZHANGW . Phylogenetic relationship between Xinjiang wild apple(Malus sieversii Roem.)and Chinese apple(Malus × domestica subsp. chinensis)based on ITS and matK sequences. Acta Horticulturae Sinica, 2014,41(2):227-239. (in Chinese) [本文引用: 1]
XU RX . Taxonomic status and phylogenetic relationships of some Malus plants based on molecular marker technology [D]. Nanjing: Nanjing Forestry University, 2018. ( in Chinese) [本文引用: 1]
LI HF . Studies on the taxonomy of the genus Malus Mill. (Rosaceae) of Taiyi-Mountains [D]. Shenyang: Shenyang Agricultural University, 2012. ( in Chinese) [本文引用: 1]
BARAKETG, OLFAS, KHALEDC, MESSAOUDM, MOHAMEDM, MOKHTART, AMEL SH . Chloroplast DNA analysis in Tunisian fig cultivars (Ficus carica L.): Sequence variations of the trnl-trnf intergenic spacer. Biochemical Systematics and Ecology, 2009,36(11):828-835. [本文引用: 1]
POTTERD, LUBY JJ, HARRISON RE . Phylogenetic relationships among species of Fragaria (Rosaceae) inferred from non-coding nuclear and chloroplast DNA sequences. Systematic Botany, 2009,25(2):337-348. [本文引用: 1]
GHADAB, AHMED BA, KHALEDC, OLFAS, MESSAOUDM, MOKHTART, AMEL SH . Molecular evolution of chloroplast DNA in fig (Ficus carica L.): Footprints of sweep selection and recent expansion. Biochemical Systematics and Ecology, 2010,38(4):563-575. [本文引用: 1]
SHAWJ, LICKEY EB, BECK JT, FARMER SB, LIU WS, MILLERJ, SIRIPUN KC, WINDER CT, SCHILLING EE, SMALL RL . The tortoise and the hare Ⅱ: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis American Journal of Botany, 2005,921(1):142-166. [本文引用: 1]
VOLK GM, HENK AD, BALDOA, FAZIOG, CHAO CT, RICHARDS CM . Chloroplast heterogeneity and historical admixture within the genus Malus. American Journal of Botany, 2015,102(7):1198-1208. [本文引用: 1]
KUMARS, STECHERG, TAMURAK . MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets Molecular Biology Evolution, 2016,33(7):1870-1874. [本文引用: 1]
LIBRADOP, ROZASJ . DnaSP v5: A software for comprehensive analysis of DNA polymorphism data Bioinformatics, 2009,25:1451-1452. [本文引用: 1]
SAITON, NEIM . The neighbor-joining method: A new method for reconstructing phylogenetic trees Molecular Biology and Evolution, 1987,4(4):406-425. [本文引用: 1]
EXCOFFIERL, LISCHER HE L . Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows Molecular Ecology Resources, 2010,10:564-567. [本文引用: 1]
KIMURAT, IKETANIH, KOTOBUKIK, MATSUTAN, BANY, HAYASHIT, YAMAMOTOT . Genetic characterization of pear varieties revealed by chloroplast DNA sequences Journal of Horticultural Science and Biotechnology, 2003,7:241-247. [本文引用: 2]
PETITR, AGUINAGALDEI, DE BEAULIEUJ, BITTKAUC, BREWERS, CHEDDADIR, ENNOSR, FINESCHIS, GRIVETD, LASCOUXM, MOHANTYA, MüLLER-STARCKG, DEMESURE- MUSCHB, PALMéA, MARTíNJ P, RENDELLS, VENDRAMING G . Glacial refugia: Hotspots but not melting pots of genetic diversity Science, 2003,300:1563-1565. [本文引用: 1]
KATAYAMAH, OGIHARAY . Phylogenetic affinities of the grasses to other monocots as revealed by molecular analysis of chloroplast DNA Current Genetics, 1996,20:527-581. [本文引用: 1]
EIADTHONGW, YONEMORIK, SUGIURAA, UTSUNOMIYAN, SUBHADRABANDHUS . Analysis of phylogenetic relationships in Mangifera by restriction site analysis of an amplified region of cpDNA Scientia Horticulturae, 1999,80:145-150. [本文引用: 1]
LIUJ, ZHENG XY, DANEILP, HU CY, TENG YW . Genetic diversity and population structure of Pyrus Calleryana (Rosaceae) in Zhejiang province, China. Biochemical Systematics and Ecology, 2012,45:69-78. [本文引用: 1]
ZONGY . Studies on genetic diversity and phylogeography of wild Pyrus betulaefolia in Northern China [D]. Hangzhou: Zhejiang University, 2014. ( in Chinese) [本文引用: 1]
NEIM . Estimation of average heterozygosity and genetic distance from a small number of individuals Genetics, 1978,89(3):583-590. [本文引用: 1]
AVISE JC, ARNOLDJ, BALL RM, BERMINGHAME, LAMBT, NEIGEL JE, REEB CA, SAUNDER NC . Intraspecific phylogeography: The mitochondrial DNA bridge between population genetics and systematics Annual Review of Ecology and Systematics, 1987,18:489-522. [本文引用: 1]
WRIGHTS. Evolution and the Genetics of Populations, Variability Within and Among Natural Populations. Chicago: The University of Chicago Press, 1978: 4. [本文引用: 1]
ZHANG CY, CHEN XS, HE TM, LIU XL, FENGT, YUAN ZH . Genetic structure of Malus sieversii population from Xinjiang, China, revealed by SSR markers. Journal of Genetics and Genomics, 2007,34(10):947-955. [本文引用: 1]
POSADAD, CRANDALL KA . Intraspecific gene genealogies: Trees grafting into networks Trends in Ecology and Evolution, 2001,16:37-45. [本文引用: 1]
 ,, 丛佩华
,, 丛佩华
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT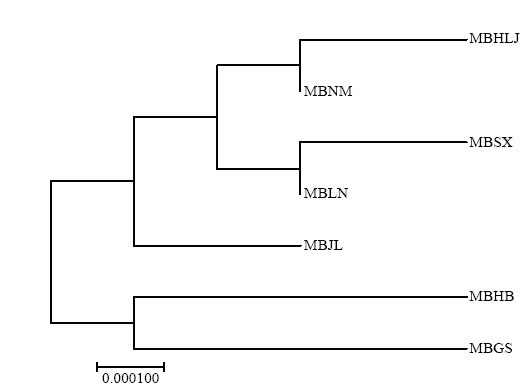 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT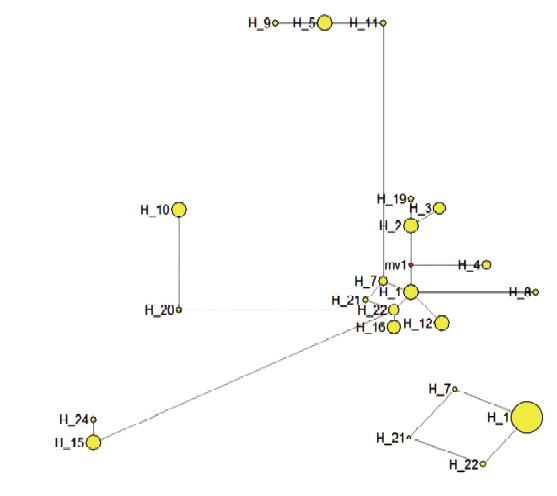 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}