Expression Pattern of KDM1A in the Development of Yak Follicles
HUANG XiangYue1, XIONG XianRong,2, HAN Jie1, YANG XianYing1, WANG Yan1, WANG Bin1, LI Jian,11 College of Life Science and Technology, Southwest Minzu University, Chengdu 610041 2 Key Laboratory of Ministry of Education for Qinghai-Tibet Plateau Animal Genetic Resources Reservation and Exploitation, Chengdu 610041
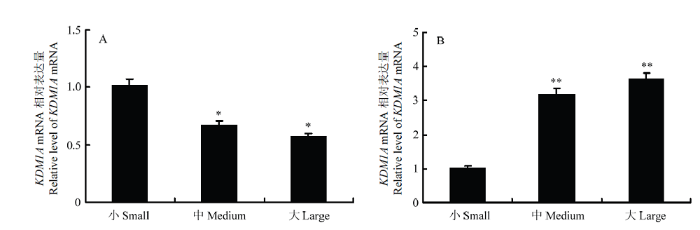
Abstract 【Objective】 The aim of this study was to analyze the role of the lysine-specific histone demethylase 1A (KDM1A) in follicle development and oocyte maturation of yak. 【Method】 Taking yak follicles as research objects and according to the size of the follicles, they were divided into three groups: large-sized (6.0-9.0 mm), medium-sized (3.0-5.9 mm), and small-sized (1.0-2.9 mm) follicles. And the cumulus-oocyte complexes (COCs) were collected from each group and cultured in vitro. The maturation rate of oocytes was counted and analyzed. The total RNA was extracted from oocyte and granulosa cells in each group of follicles. The real time quantitative PCR (RT-qPCR) was used to detect the relative expression of KDM1A during follicular development. The cell localization and expression of KDM1A in yak follicle were detected by immunohistochemistry, and the correlation analysis was performed by in vitro maturation and RT-qPCR. 【Result】 The maturation rate of oocytes in vitro maturation was positively related to the size of follicles and showed a rising trend with progressing of follicular development. Meanwhile, the oocyte maturation rates of large-sized, medium-sized and small-sized follicle oocytes were 90.53 %, 88.10 % and 55.14 %, respectively. The result of RT-qPCR was found that KDM1A gene of yak was widely expressed during the development of follicles, and its expression level was significantly different in developmental stages of follicles. The relative expression of mRNA in the oocytes of the large and medium-sized follicle was significantly lower than small-sized follicles (P < 0.05), but the relative expression of granulosa cells in the small-sized follicle was significantly lower than large and medium-sized (P < 0.01), and there was no difference in mRNA expression levels between oocytes and granulosa cells in the large and middle-sized (P > 0.05). The results of immunohistochemistry showed that KDM1A was expressed in granulosa cells and membrane cells of follicles, and its expression trend was consistent with RT-qPCR. The expression of KDM1A was the highest in the large follicle and increased with the development of follicles. 【Conclusion】 The expression levels of KDM1A mRNA and protein in oocytes and granulosa cells at the development of yak follicles were dynamic, which indicated that KDM1A played an important role in follicular development and oocyte maturation. It might be related to meiosis of oocytes and proliferation and differentiation of granulosa cells, and these results of study would provide a basic data for further research of the mechanism of KDM1A in the meiosis of yak oocytes. Keywords:Yak;KDM1A;immunohistochemistry;follicle;expression
PDF (1456KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄向月, 熊显荣, 韩杰, 杨显英, 王艳, 王斌, 李键. KDM1A在牦牛卵泡发育过程中的表达[J]. 中国农业科学, 2019, 52(24): 4624-4631 doi:10.3864/j.issn.0578-1752.2019.24.016 HUANG XiangYue, XIONG XianRong, HAN Jie, YANG XianYing, WANG Yan, WANG Bin, LI Jian. Expression Pattern of KDM1A in the Development of Yak Follicles[J]. Scientia Acricultura Sinica, 2019, 52(24): 4624-4631 doi:10.3864/j.issn.0578-1752.2019.24.016
** 表示差异极显著(P < 0.01);* 表示差异显著(P < 0.05) Fig. 1The expression of KDM1A in the oocyte (A) and the granular (B) cells of the different development stages in Yak follicle
** show extremely significant difference (P < 0.01); * show significant difference (P < 0.05)
Table 1 表1 表1牦牛卵母细胞成熟度 Table 1Maturation rate of yak oocytes
A:牦牛大卵泡组织中KDM1A的定位;B:牦牛中卵泡组织中KDM1A的定位情况;C:牦牛小卵泡组织中KDM1A的定位情况(GC:壁层颗粒细胞;TC:膜细胞) Fig. 2Localization of KDM1A in Yak follicle
A: Localization of KDM1A in large-sized follicle of yak; B: Localization of KDM1A in medium-sized follicle of yak; C: Localization of KDM1A in small-sized follicle of yak (GC: Granular cells; TC: Theca cells)
WANGR, YU SJ, CUIY . Observation of microstructure and ultrastructure of the thyroid gland in juvenile yak Veterinary Science in China, 2009,39(4):357-361. (in Chinese) [本文引用: 1]
LAN DL, XIONG XR, WEI YL, XUT, ZHONG JC, ZI XD, WANGY, LIJ . RNA-Seq analysis of yak ovary: improving yak gene structure information and mining reproduction-related genes Scientia Sinica,2014,44(3):307-317. (in Chinese) [本文引用: 1]
SINHA PB, TESFAYED, RINGSF, HOSSIENM, HOELKERM, HELDE, NEUHOFFC, THOLENE, SCHELLANDERK, SALILEW- WONDIMD . MicroRNA-130b is involved in bovine granulosa and cumulus cells function, oocyte maturation and blastocyst formation Journal of Ovarian Research, 2017,10(1):37. [本文引用: 1]
HUNT PA, HASSOLD TJ . Human female meiosis: what makes a good egg go bad ? Trends in Genetics, 2008,24(2):86-93. [本文引用: 1]
PETRO EM, LEROY JL, COVACIA, FRANSENE, DE NEUBOURGD, DIRTU AC, DE PAUWI, BOLS PE . Endocrine- disrupting chemicals in human follicular fluid impair in vitro oocyte developmental competence Human Reproduction, 2012,27(4):1025-1033. [本文引用: 1]
HAYASHIK, YOSHIDAK, MATSUIY . A histone H3 methyltransferase controls epigenetic events required for meiotic prophase Nature, 2005,438(7066):374-378. [本文引用: 1]
TACHIBANAM, NOZAKIM, TAKEDAN, SHINKAIY . Functional dynamics of H3K9 methylation during meiotic prophase progression Embo Journal, 2014,26(14):3346-3359. [本文引用: 1]
FEIL, NOTTKE AC, YANGS . Mechanisms involved in the regulation of histone lysine demethylases. Current Opinion in Cell Biology, 2008,20(3):316-325. [本文引用: 1]
SHAO GB, HUANG XJ, GONG AH, ZHANG ZJ, LU RZ, SANG JR . Histone to methylase LSD1 and its biological functions Hereditas, 2010,04:331-338. (in Chinese) [本文引用: 1]
GARCIA-BASSETSI, KWON YS, TELESEF, PREFONTAINE GG, HUTT KR, CHENG CS, JU BG, OHGI KA, WANGJ, ESCOUBET-LOZACHL, ROSE DW, GLASS CK, FU XD, ROSENFELD MG . Histone methylation-dependent mechanisms impose ligand dependency for gene activation by nuclear receptors Cell, 2007,128(3):505-518. [本文引用: 1]
JEESUNK, KUMAR SA, YOKOT, LINK, SHENJ, LUY, KERENYI MA, ORKIN SH, CHENT . LSD1 is essential for oocyte meiotic progression by regulating CDC25B expression in mice Nature, 2015,6:10116. [本文引用: 2]
SHAOG, WANGJ, LIY . Lysine-specific demethylase1 mediates epidermal growth factor signaling to promote cell migration in ovarian cancer cells Scientific Reports, 2015,5:15344. [本文引用: 2]
ZHENG YC, MAJ, WANGZ, LIJ, JIANGB, ZHOUW, SHIX, WANGX, ZHAOW, LIU HM . A systematic review of histone lysine-specific demethylase 1 and its inhibitors Medicinal Research Reviews, 2015,35(5):1032-1071. [本文引用: 1]
LYNCH JT, HARRIS WJ, SOMERVAILLE TC . LSD1 inhibition: a therapeutic strategy in cancer? Expert Opinion on Therapeutic Targets, 2012,16(12):1239-1249. [本文引用: 1]
OMBRA MN, DI SA, ABBONDANZAC, MIGLIACCIOA, AVVEDIMENTO EV, PERILLOB . Retinoic acid impairs estrogen signaling in breast cancer cells by interfering with activation of LSD1 via PKA Biochimica et Biophysica Acta, 2013,1829(5):480-486. [本文引用: 1]
CICCONE DN, SUH, HEVIS, GAYF, LEIH, BAJKOJ, XUG, LIE, CHENT . KDM1B is a histone H3K4 demethylase required to establish maternal genomic imprints Nature, 2009,461(7262):415-418. [本文引用: 1]
JOHNSON MT, FREEMAN EA, GARDNER DK, HUNTPA . Oxidative metabolism of pyruvate is required for meiotic maturation of murine oocytes in vivo Biology of Reproduction, 2007,77(1):2-8. [本文引用: 1]
SANCHEZLAZOL, BRISARDD, ELISS, MAILLARDV, UZBEKOVR, LABASV, DESMARCHAISA, PAPILLIERP, MONGETP, UZBEKOVAS . Fatty acid synthesis and oxidation in cumulus cells support oocyte maturation in bovine Molecular Endocrinology, 2014,28(9):1502-1521. [本文引用: 1]
HUANGZ, LIS, WEIS, LIX, LIQ, ZHANGZ, HANY, ZHANGX, MIAOS, DUR, WANGL . Lysine-specific demethylase 1 (LSD1/ KDM1A) contributes to colorectal tumorigenesis via activation of the Wnt/β-catenin pathway by down-regulating Dickkopf-1 (DKK1) PLoS One, 2013,8(7):70077. [本文引用: 1]
ANCELINK, SYXL, BORENSZTEINM, RANISAVLJEVICN, VASSILEVI, BRISE?O-ROAL, LIUT, METZGERE, SERVANTN, BARILLOTE1, CHEN CJ, SCHüLER, HEARDE . Maternal LSD1/KDM1A is an essential regulator of chromatin and transcription landscapes during zygotic genome activation Elife, 2016,2(5):e08851. [本文引用: 1]
WASSON JA, SIMON AK, MYRICK DA, WOLFG, DRISCOLLS, PFAFFSL, MACFARLAN TS, KATZ DJ . Maternally provided LSD1/KDM1A enables the maternal-to-zygotic transition and prevents defects that manifest postnatally Elife, 2016,27(5):e08848. [本文引用: 1]
YOKOYAMAA, IGARASHIK, SATOT, TAKAGIK, OTSUKA IM, SHISHIDOY, BABAT, ITOR, KANNOJ, OHKAWAY, MOROHASHIK, SUGAWARAA . Identification of myelin transcription factor 1 (MyT1) as a subunit of the neural cell type-specific lysine-specific demethylase 1 (LSD1) complex Journal of Biological Chemistry, 2014,289(26):18152-18162. [本文引用: 1]
SU OJ, JIN HS, MARCOC . Wee1B, Myt1, and Cdc25 function in distinct compartments of the mouse oocyte to control meiotic resumption Journal of Cell Biology, 2010,188(2):199-207. [本文引用: 1]
ABDIS, SALEHNIAM, HOSSEINKHANI . Quality of oocytes derived from vitrified ovarian follicles cultured in two- and three-dimensional culture system in the presence and absence of kit ligand Biopreservation and Biobanking, 2016,14(4):279-288. [本文引用: 1]
WIGGLESWORTHK, LEE KB, O'BRIEN MJ, PENGJ, MATZUK MM, EPPIG JJ . Bidirectional communication between oocytes and ovarian follicular somatic cells is required for meiotic arrest of mammalian oocytes Proceedings of the National Academy of Sciences of the United States of America, 2013,110(39):3723-3729. [本文引用: 1]
ALMH, KATSKAKSIAZKIEWICZL, RY?SKAB, TUCHSCHERERA . Survival and meiotic competence of bovine oocytes originating from early antral ovarian follicles Theriogenology, 2006,65(7):1422-1434. [本文引用: 1]
LEQUARRE AS, VIGNERONC, RIBAUCOURF, HOLMP, DONNAYIDALBIèS-TRANRCALLESENHMERMILLODP . Influence of antral follicle size on oocyte characteristics and embryo development in the bovine Theriogenology, 2005,63(3):841-859. [本文引用: 1]
YU XL, ZAN LS, DENGW, PANG YZ, WANG XZ . Effects of follicle size and bovine follicular fluid on developmental competence of bovine oocytes following maturation, fertilization and culture in vitro. Scientia Agricultural Sinica, 2005,38(8):1664-1668. (in Chinese) [本文引用: 1]
KIM JH, PARK UH, MOONM, UM SJ, KIM EJ . Negative regulation of ERa by a novel protein CAC1 through association with histone demethylase LSD1 Febs Letters, 2013,587(1):17-22. [本文引用: 1]
 ,2, 韩杰1, 杨显英1, 王艳1, 王斌1, 李键
,2, 韩杰1, 杨显英1, 王艳1, 王斌1, 李键
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT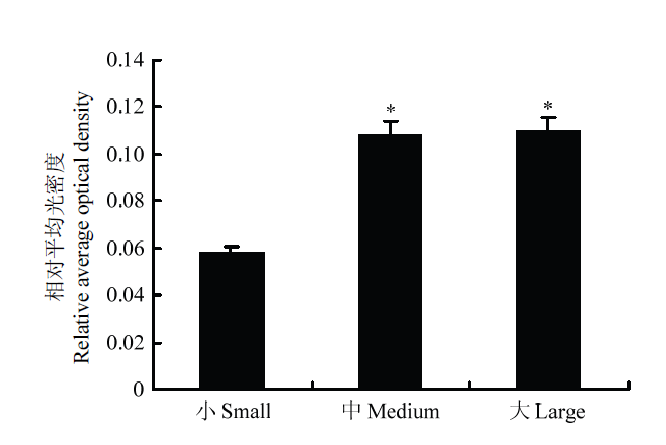 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}