 ,1
,1Phenotype Character Analysis and Evaluation of Modern Rose Cultivars
GUO Cong1, GUAN Wei1, ZENG XiangGuo1, ZHANG QingHua1, XIANG FaYun1, SONG YueJun2, HAN YongChao,1通讯作者:
责任编辑: 赵伶俐
收稿日期:2019-05-20接受日期:2019-07-25网络出版日期:2019-12-16
| 基金资助: |
Received:2019-05-20Accepted:2019-07-25Online:2019-12-16
作者简介 About authors
过聪,E-mail:guocong602@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (2053KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
过聪, 关伟, 曾祥国, 张庆华, 向发云, 宋粤君, 韩永超. 现代月季品种表型性状分析与评价[J]. 中国农业科学, 2019, 52(24): 4632-4646 doi:10.3864/j.issn.0578-1752.2019.24.017
GUO Cong, GUAN Wei, ZENG XiangGuo, ZHANG QingHua, XIANG FaYun, SONG YueJun, HAN YongChao.
0 引言
【研究意义】月季(Rosa spp.)是蔷薇科(Rosaceae)蔷薇属(Rosa L.)非常重要的观赏花卉,一直以来备受育种家的青睐,品种层出不穷,至今已经有35 000余个品种[1]。现代月季花型饱满、花瓣数多、开花直径大、花色柔和典雅、栽培品质较好,可以广泛应用于城市绿化和家庭园艺,受到国内外消费大众的热烈追捧。我国市场上的现代月季品种多从国外引入,由于国内对月季品种缺乏统一分类标准和方法,导致品种间命名混乱,性状混淆,分类不清,极大增加了月季新品种培育、科学研究和市场开发的难度[2]。【前人研究进展】分析月季资源性状并评价月季品种,有助于掌握月季资源的观赏性、抗性和适应性等品种特性[3,4,5],加强对月季资源的分类管理,发掘具有重要经济价值和科研价值的品种资源[6,7,8]。分析月季资源性状的多样性与变异性等信息,可以深入了解性状遗传规律,鉴定月季资源及其杂交后代[9,10],对月季的应用和科研都有非常积极的促进作用。月季资源性状的评价方式多样,包括形态学观察、细胞倍性检测、核型分析、分子标记等手段[11,12,13,14,15]。其中分子标记作为辅助育种手段在月季的观赏特性、病虫害抗性以及远缘杂交育种上都有深入研究[16,17,18]。现代月季品种以四倍体为主,是由10个二倍体原始种杂交而来,其间月季基因型和表型经历了高度多样化和异质化过程[19,20,21]。目前,月季分子研究工作主要基于二倍体种类,这样的研究一定程度上受二倍体群体大小限制,在商业上和月季育种上的价值不大[14,20,22]。加上月季花色、花瓣数等多种表型性状不仅是数量遗传,还是基因型与环境共同影响的结果[23,24,25]。因此,在月季品种鉴别与育种方面仍需要全面而深入地结合月季表型性状进行分析[26,27]。另外,月季的叶部表型不但与观赏性相关,在光合作用、抗病和抗逆性上也非常重要,而有关月季叶部的性状研究还有待深入[28,29,30,31]。【本研究切入点】表型观测是研究植物遗传多样性最直观和最基础的方法,通过这一方式可以从整体上了解资源的丰富程度,为基因和细胞研究提供直观依据。本研究参考前人对现代月季表型和遗传规律的研究结果,选择最能直观感受的花部和叶部表型指标,对月季进行较全面的表型分析与综合评价。【拟解决的关键问题】通过对60个现代月季品种的11个表型性状观测统计,获得各表型性状的变异性和遗传多样性,揭示各性状遗传变异规律;通过性状相关性以及主成分分析,进一步筛选合理的月季品种评价指标,为科学收集月季品种资源并开展研究提供理论依据。1 材料与方法
1.1 品种材料
供试月季为湖北省农科院经济作物研究所引进的60个现代月季栽培品种,其中中国培育的品种3个,英国品种5个,意大利品种1个,日本品种29个,美国品种4个,荷兰品种4个,德国品种1个,法国品种10个,丹麦品种3个。各月季中文商品名与对应英文名如表1,各品种月季植株均为二年生以上苗,于田间露天栽培(东经114°32',北纬30°49′)。试验观测期间进行正常水肥管理。Table 1
表1
表160个月季资源品种名与培育地
Table 1
| 品种编码 Code | 品种名 Cultivar name | 培育地 Breeding area | 品种编码 Code | 品种名 Cultivar name | 培育地 Breeding area | |
|---|---|---|---|---|---|---|
| C1 | 安布里奇Ambridge Rose | 英国Britain | C31 | 玛姬婶婶Aunt Margys | 意大利Italy | |
| C2 | 彩蝶Cai Die | 中国China | C32 | 玛丽玫瑰マリーローズ | 日本Japan | |
| C3 | 草药ラムカン | 日本Japan | C33 | 芒果浪漫Mango Romantica | 法国France | |
| C4 | 超级绿色スーパーグリーン | 日本Japan | C34 | 朦胧泡泡Misty Bubbles | 荷兰Netherlands | |
| C5 | 橙色浪漫オレンジロマンティカ | 日本Japan | C35 | 蜜桃雪山Peach Avalanche+ | 荷兰Netherlands | |
| C6 | 雏鸡ひよこ | 日本Japan | C36 | 魔果モコモコ | 日本Japan | |
| C7 | 蝶之舞セシル ドゥ ヴォーランジェ | 日本Japan | C37 | 莫奈Claude Monet | 法国France | |
| C8 | 丁香经典ライラッククラシック | 日本Japan | C38 | 耐心Patience | 英国Britain | |
| C9 | 躲躲藏藏カシュ・カシュ | 日本Japan | C39 | 纽带Kizuna | 法国France | |
| C10 | 蜂蜜焦糖Honey Caramel | 法国France | C40 | 苹果挞タルト・ポム | 日本Japan | |
| C11 | 红苹果レッド アップル ローズ | 日本Japan | C41 | 蜻蜓リベルラ | 日本Japan | |
| C12 | 胡里奥Julio Iglesias | 法国France | C42 | 秋日胭脂オータムルージュ | 日本Japan | |
| C13 | 欢笑格鲁吉亚Teasing Georgia | 英国Britain | C43 | 神秘Mysterieuse | 法国France | |
| C14 | 幻彩アルヌワブラン | 日本Japan | C44 | 桃香ももか | 日本Japan | |
| C15 | 皇家ロワイヤル | 日本Japan | C45 | 藤本彩虹仙境Rainbows End | 美国America | |
| C16 | 黄金呼呼塞拉Golden Vuvuzela | 荷兰the Netherlands | C46 | 天方夜谭シェエラザード | 日本Japan | |
| C17 | 黄金庆典Golden Celebration | 英国Britain | C47 | 甜蜜果酱Honey Jam | 日本Japan | |
| C18 | 婚礼钟声クロッシュドマリアージュ | 日本Japan | C48 | 甜蜜马车Sweet Chariot | 美国America | |
| C19 | 加百列大天使ガブリエル | 日本Japan | C49 | 我爱你ティアモ | 日本Japan | |
| C20 | 金丝雀カナリアリッチ | 日本Japan | C50 | 无条件的爱Unconditional love | 美国America | |
| C21 | 京みやこ | 日本Japan | C51 | 夏日花火Xiari Huahuo | 中国China | |
| C22 | 卡哈拉Kahala | 美国America | C52 | 香织装饰かおりかざり | 日本Japan | |
| C23 | 凯丽Carey | 英国Britain | C53 | 小魔女甜心Péché Mignon | 法国France | |
| C24 | 狂欢派对パーティーラナンキュラ | 日本Japan | C54 | 新浪潮ニューウェーブ,フォルム | 日本Japan | |
| C25 | 拉米炫耀Lamy Parade | 丹麦Denmark | C55 | 新星2009Nova 2009 Hit | 丹麦Denmark | |
| C26 | 蓝色阴雨レイニー ブルー | 日本Japan | C56 | 阳光海滩サニービーチ | 日本Japan | |
| C27 | 浪漫宝贝Baby Romantica | 法国France | C57 | 樱桃白兰地Cherry Brandy | 德国Germany | |
| C28 | 浪漫珊瑚心Romantica Vuvuzela | 荷兰the Netherlands | C58 | 永远的艾帕索El Paso Forever | 丹麦Denmark | |
| C29 | 榴花秋舞Liuhua Qiuwu | 中国China | C59 | 珍妮莫罗Jeanne Moreau | 法国France | |
| C30 | 马萨德医生Docteurs Massad | 法国France | C60 | 智慧女神M-ミネルヴァ | 日本Japan |
新窗口打开|下载CSV
1.2 数据采集方法
2018年4—8月对供试月季表型性状进行观测记录,主要记录月季植株的株形、小叶数、全叶长、顶小叶宽、节间距、花色、花朵直径、花瓣数、花头数、花朵衰老方式等。各数量性状采用具体测量法,调查数均大于等于5个。其中叶片性状取各品种植株基部以上3—4节处成熟叶调查其小叶数、全叶长和顶小叶宽;节间距为各品种中部成熟叶片间距离;取各品种盛开花测量花朵直径。根据单花茎上花头数将月季分为单头、多头、丰花3种花头数性状,其中单头为一根花枝上有且仅有一朵花,多头型为花枝上在原单头花开放时,花下1—2节处形成新的多个花苞(一般2—5个),并依次开放;丰花型为一根花枝上单个托叶以上花梗数5个以上,且花朵间开花时间较一致(图1-a—c);根据各品种花瓣数多少,取40片、90片为两个节点将月季分为3个级别:花瓣数少于40片,花瓣数多于40片且少于90片,花瓣数90片以上。月季花型分4类:外层花瓣大,盛开后花朵依然包紧不露花心或中心花瓣的归为包子型;酒杯状花型的归为杯型;花瓣均外展,中心花蕊可见或花心仅被数瓣遮挡,花开形似牡丹的归为牡丹型;花朵外部数圈花瓣较大,围成碗型,内部花瓣小而多,可分区成4或5角星形的为碗型花(图1-d—g)。花朵衰老方式分为3类:枯萎型花朵花瓣先枯焦后落瓣或不落;落瓣型花朵花瓣未枯黄先落瓣;另一种为两种衰老方式兼有。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1月季花头数和花型分类示例
a:单头(‘超级绿色’);b:多头(‘黄金呼呼塞拉’);c:丰花(‘甜蜜马车’);d:包子型(‘幻彩’);e:杯型(‘新星2009’);f:牡丹型(‘天方夜谭’);g:碗型(‘黄金呼呼塞拉’)
Fig. 1Examples in classification of flower number and flower shape
a: Single flower (Rosa ‘スーパーグリーン’); b: Successive flower (Rosa ‘Golden Vuvuzela’); c: Multi-flower (Rosa ‘Sweet Chariot’); d: Ball (Rosa ‘アルヌワブラン’); e: Cup (Rosa ‘Nova 2009 Hit’); f: Peony (Rosa ‘シェエラザード’); g: Bowl (Rosa ‘Golden Vuvuzela’)
1.3 数据统计与分析
对调查的11个表型指标数据采用Microsoft Excel 2007软件(微软公司,中国)-统计并分析变异系数、遗传多样性,使用Statistical Product and Service Solutions 19.0 (SPSS 19.0)软件(IBM公司,中国)进行指标相关性分析和主成分分析。各月季植株的4个数量指标(Quantitative character)用统计数值直接表示;花瓣数根据统计数值转化为分类指标后编码统计;7个定性指标分别使用编码分类统计。其中二元指标(Two-state character)1个,有序多态指标(Ordered multi-state character)2个,无序多态指标(Disordered multi-state character)4个,各指标类型及编码方式如表2所示。各指标统计结果使用平均值与标准差完成数据标准化。根据性状指标的平均值(Mean,μ)和标准差(SD,δ)计算变异系数(coefficient of variation,CV),计算公式为:CV=δ/μ。根据各性状级别出现频率计算Shannon-Wiener多样性指数(H),其计算公式为:H= -ƩPiInPi(i=1,2,3,……),其中Pi是某性状第i个级别的材料数占总材料数的百分比。
Table 2
表2
表2重要性状分类与编码
Table 2
| 编号 Code | 性状 Character | 分类细节 Classification detail |
|---|---|---|
| P1 | 株形Habit | 0:微型;1:直立型;2:扩张型;3:藤蔓型 0: Mini; 1: Erect; 2: Expanding; 3: Vine |
| P2 | 全叶长Leaf length (cm) | 取测量平均值Using the mean value |
| P3 | 顶小叶宽Top leaflet width (cm) | 取测量平均值Using the mean value |
| P4 | 节间距Distance between leaves (cm) | 取测量平均值Using the mean value |
| P5 | 小叶数Leaflet number | 取测量平均值Using the mean value |
| P6 | 花头数Flower number | 0:单头;1:多头;2:丰花 0: Single flower; 1: Successive flower; 2: Multi-flower |
| P7 | 花朵直径Flower diameter (cm) | 取测量平均值Using the mean value |
| P8 | 花瓣数Petal number | 0:<等于40;1:>40,<等于90;2:>90 |
| P9 | 花色 Flower color | 0:白色;1:粉色;2:红色;3:紫红色;4:紫色;5:杏色;6:黄色;7:橙色;8:绿色; 9:复色;10:混色;11:条纹 0: White; 1: Pink; 2: Red; 3: Red-purple; 4: Purple; 5: Apricot; 6: Yellow; 7: Orange; 8: Green; 9: Multi-color; 10: Mix-color; 11: Stripe |
| P10 | 花型Flower shape | 0:包子型;1:杯型;2:牡丹型;3:碗型 0: Ball; 1: Cup; 2: Peony; 3: Bowl |
| P11 | 花朵衰老方式 Flower senescence | 0:枯萎为主;1:枯萎和落瓣均有;2:落瓣为主 0: Petal abscission; 1: Abscission and withering; 2: Withering |
新窗口打开|下载CSV
性状指标相关性分析方法如下:使用Pearson检测分析4个数量性状指标间相关性;使用Kendall检测分析无序多态指标与其他指标间的相关性;使用Spearman检测分析数量性状指标与有序多态性状指标间的相关性,相关性结果分为高度相关(|r|>等于0.8)、中度相关(0.5<等于|r|<0.8)和低度相关(0.3<等于|r|<0.5)以及弱相关(|r|<0.3)4个等级。对相关性显著的指标进行进一步方差分析。将各性状数据标准化后进行主成分分析,获得主要评价指标。主成分综合得分主要计算方法:(1)计算各性状数据在主成分中的特征向量,根据各性状在4个主成分中的特征向量值及标准化数据,经计算获得月季品种的各成分因子得分(Fi值,i=成分因子序号);(2)根据各主成分因子在累积因子特征值(U)中的比值计算出各因子权重系数,最终计算月季品种的主成分综合得分(F值),计算公式为:F=ƩUiFi/U(i=1,2,3,……),进而为月季品种分组聚类,筛选评价特异资源。
2 结果
2.1 月季品种非花部表型遗传多样性
2.1.1 月季株形分类根据供试月季品种地栽生长状态,将月季分为微型、直立型、扩张型和藤蔓型4种株形。其中32个品种花枝挺立,枝条向上生长,扩张性不强,归为直立型品种,占总资源的53%;21个品种花枝细软或枝条成扩张生长,归为扩张型品种,占总资源的35%;微型品种有5个,它们枝条长度抽生较短,开花时田间株高低于40 cm;藤蔓型品种2个,枝条抽生长,具蔓性。
2.1.2 月季叶部指标性状比较
月季的叶部形态不仅与其观赏价值有关,还影响其光合作用和抗病虫害特性。选取各供试月季品种的成熟叶片测量月季叶片大小。月季单片复叶平均全叶长为11.6 cm,品种间变异系数为21.6%。‘躲躲藏藏’的叶片全长最短,为3.8 cm,全叶长最大的是‘神秘’,为18 cm,月季品种全叶长变化大的品种有‘樱桃白兰地’‘雏鸡’和‘皇家’,变异系数分别为28.7%、22.4%和22.0%,均大于品种间差异;而‘安布里奇’品种的全叶长最为均一,品种内变异系数仅为1.9%。
如表3所示,60个月季品种的顶小叶平均叶宽3.2 cm,品种间变异系数25%。其中顶小叶最宽的品种为‘马萨德医生’,叶宽5.7 cm,最窄的为‘躲躲藏藏’,叶宽0.9 cm。‘皇家’和‘加百列大天使’的顶小叶宽最不稳定,变异系数分别为35.0%和35.8%,远远大于品种间差异,而顶小叶宽相对稳定的是‘蜜桃雪山’,品种内变异系数为5.7%。
Table 3
表3
表3月季株形与叶片性状多样性分析
Table 3
| 品种编号 Code | P1株形 Habit | P2全叶长Leaf length | P3顶小叶宽Top leaflet width | P4节间距Distance between leafs | P5小叶数 Leaflet number | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 均值 Mean (cm) | 标准差 SD | 变异系数 CV (%) | 均值 Mean (cm) | 标准差 SD | 变异系数 CV (%) | 均值 Mean (cm) | 标准差 SD | 变异系数 CV (%) | |||
| C1 | 1 | 11.2 | 0.2 | 1.9 | 2.9 | 0.2 | 7.7 | 4.3 | 0.5 | 11.9 | 7 |
| C2 | 2 | 13.2 | 1.1 | 8.6 | 3.1 | 0.3 | 9.0 | 3.4 | 0.6 | 18.9 | 7 |
| C3 | 1 | 12.5 | 1.6 | 13.0 | 3.8 | 0.5 | 12.3 | 4.7 | 0.7 | 14.3 | 5 |
| C4 | 2 | 10.6 | 2.1 | 19.5 | 3.2 | 0.3 | 8.4 | 4.3 | 0.4 | 10.4 | 5 |
| C5 | 1 | 15.0 | 2.0 | 13.1 | 4.8 | 0.5 | 9.6 | 4.3 | 0.7 | 15.6 | 7 |
| C6 | 1 | 13.1 | 2.9 | 22.4 | 2.9 | 0.4 | 12.7 | 3.3 | 0.9 | 27.5 | 5 |
| C7 | 2 | 5.3 | 0.7 | 13.4 | 2.0 | 0.2 | 12.6 | 3.2 | 0.3 | 8.6 | 5 |
| C8 | 1 | 8.0 | 0.6 | 7.2 | 3.5 | 0.4 | 10.9 | 3.6 | 0.3 | 9.5 | 5 |
| C9 | 0 | 3.8 | 0.3 | 7.0 | 0.9 | 0.2 | 17.9 | 1.7 | 0.3 | 16.1 | 5 |
| C10 | 1 | 10.1 | 0.8 | 8.2 | 2.7 | 0.2 | 7.1 | 3.0 | 0.4 | 14.9 | 5 |
| C11 | 1 | 14.0 | 2.2 | 15.7 | 3.9 | 0.5 | 13.7 | 4.5 | 0.4 | 9.9 | 7 |
| C12 | 2 | 11.4 | 1.9 | 16.4 | 3.4 | 0.3 | 9.4 | 3.2 | 0.3 | 8.6 | 7 |
| C13 | 2 | 13.6 | 1.1 | 8.2 | 3.6 | 0.4 | 10.1 | 4.3 | 0.3 | 7.9 | 7 |
| C14 | 1 | 15.1 | 0.8 | 5.1 | 3.6 | 0.8 | 22.0 | 4.3 | 0.8 | 19.5 | 5 |
| C15 | 1 | 11.8 | 2.6 | 22.0 | 3.7 | 1.3 | 35.0 | 3.9 | 0.7 | 18.4 | 5 |
| C16 | 1 | 14.5 | 2.2 | 15.3 | 3.7 | 0.6 | 15.4 | 4.2 | 0.6 | 13.6 | 7 |
| C17 | 2 | 11.1 | 0.9 | 8.4 | 2.9 | 0.3 | 10.8 | 4.0 | 0.6 | 14.8 | 5 |
| C18 | 1 | 14.5 | 1.1 | 7.7 | 2.0 | 0.2 | 7.8 | 4.3 | 0.4 | 9.1 | 5 |
| C19 | 2 | 11.9 | 1.8 | 15.1 | 3.3 | 1.2 | 35.8 | 3.8 | 0.6 | 15.0 | 7 |
| C20 | 1 | 10.8 | 2.2 | 20.0 | 2.8 | 0.6 | 22.9 | 1.7 | 0.3 | 16.1 | 5 |
| C21 | 1 | 11.1 | 0.8 | 7.0 | 3.2 | 0.6 | 17.7 | 4.6 | 0.4 | 9.1 | 5 |
| C22 | 2 | 14.8 | 1.2 | 8.2 | 4.0 | 0.3 | 6.6 | 4.5 | 0.4 | 9.9 | 5 |
| C23 | 1 | 12.8 | 0.7 | 5.1 | 3.2 | 0.3 | 10.1 | 5.0 | 0.6 | 12.2 | 7 |
| C24 | 2 | 12.6 | 1.2 | 9.3 | 3.2 | 0.3 | 10.3 | 3.4 | 0.6 | 17.1 | 5 |
| C25 | 2 | 7.7 | 0.7 | 9.5 | 1.9 | 0.2 | 12.9 | 2.2 | 0.2 | 9.5 | 5 |
| C26 | 3 | 14.0 | 1.5 | 10.6 | 2.9 | 0.3 | 11.7 | 2.8 | 0.2 | 8.1 | 5 |
| C27 | 0 | 9.0 | 0.8 | 8.5 | 2.6 | 0.3 | 12.8 | 4.2 | 0.6 | 13.6 | 5 |
| C28 | 1 | 11.6 | 0.8 | 6.8 | 4.6 | 0.6 | 12.5 | 3.2 | 0.3 | 8.6 | 7 |
| C29 | 2 | 9.1 | 1.7 | 19.1 | 1.8 | 0.2 | 9.8 | 3.0 | 0.0 | 0.0 | 7 |
| C30 | 1 | 12.2 | 1.3 | 11.0 | 5.7 | 0.5 | 9.2 | 3.2 | 1.0 | 32.4 | 5 |
| C31 | 0 | 10.7 | 1.0 | 9.4 | 2.0 | 0.2 | 9.9 | 2.5 | 0.4 | 14.1 | 7 |
| C32 | 2 | 10.6 | 0.7 | 6.2 | 2.9 | 0.3 | 9.9 | 2.8 | 0.2 | 6.2 | 5 |
| C33 | 1 | 8.9 | 1.3 | 14.7 | 3.5 | 0.4 | 12.7 | 3.2 | 0.8 | 23.7 | 5 |
| C34 | 1 | 11.8 | 1.4 | 11.9 | 3.8 | 0.3 | 9.0 | 3.5 | 0.5 | 14.3 | 7 |
| C35 | 1 | 11.6 | 1.4 | 11.9 | 3.2 | 0.2 | 5.7 | 4.8 | 0.3 | 7.1 | 5 |
| C36 | 2 | 13.4 | 0.9 | 6.7 | 3.0 | 0.2 | 6.8 | 4.0 | 0.6 | 15.3 | 5 |
| C37 | 1 | 10.4 | 0.8 | 7.6 | 2.8 | 0.3 | 10.0 | 3.1 | 0.4 | 13.5 | 5 |
| C38 | 1 | 15.3 | 1.8 | 11.6 | 4.1 | 0.6 | 13.7 | 5.3 | 0.2 | 4.3 | 7 |
| C39 | 2 | 14.6 | 2.6 | 17.9 | 3.6 | 0.4 | 10.8 | 3.4 | 0.1 | 3.8 | 5 |
| C40 | 2 | 10.8 | 0.5 | 4.5 | 2.8 | 0.5 | 16.6 | 2.8 | 0.3 | 9.8 | 5 |
| C41 | 1 | 8.3 | 0.6 | 7.1 | 3.1 | 0.3 | 10.1 | 3.1 | 0.3 | 11.1 | 5 |
| C42 | 2 | 11.3 | 1.2 | 11.1 | 3.2 | 0.4 | 11.7 | 5.7 | 0.7 | 11.8 | 5 |
| C43 | 1 | 18.0 | 1.8 | 10.1 | 4.0 | 0.4 | 9.3 | 3.1 | 0.2 | 7.2 | 7 |
| C44 | 0 | 10.5 | 0.5 | 4.9 | 2.0 | 0.2 | 11.9 | 4.9 | 0.2 | 4.6 | 5 |
| C45 | 3 | 7.5 | 0.8 | 10.0 | 2.1 | 0.5 | 23.3 | 2.3 | 0.1 | 5.5 | 5 |
| C46 | 2 | 11.3 | 2.4 | 21.5 | 3.2 | 0.4 | 14.1 | 3.3 | 0.2 | 6.9 | 7 |
| C47 | 1 | 12.4 | 2.3 | 18.7 | 3.7 | 0.6 | 16.2 | 3.3 | 0.4 | 13.6 | 5 |
| C48 | 0 | 7.9 | 1.5 | 19.5 | 1.7 | 0.4 | 23.1 | 3.0 | 0.7 | 23.6 | 7 |
| C49 | 1 | 12.8 | 0.9 | 7.3 | 3.9 | 0.3 | 8.0 | 2.7 | 0.3 | 10.1 | 5 |
| C50 | 2 | 10.2 | 1.2 | 11.3 | 2.0 | 0.4 | 22.1 | 2.3 | 0.4 | 19.4 | 7 |
| C51 | 2 | 12.3 | 1.1 | 8.9 | 2.9 | 0.3 | 9.4 | 3.9 | 0.2 | 5.7 | 5 |
| C52 | 2 | 12.9 | 1.9 | 14.8 | 2.6 | 0.7 | 25.4 | 3.3 | 0.3 | 8.3 | 5 |
| C53 | 1 | 10.8 | 0.8 | 7.6 | 3.0 | 0.2 | 8.2 | 3.2 | 0.3 | 10.8 | 7 |
| C54 | 2 | 14.6 | 1.0 | 7.2 | 3.9 | 0.4 | 10.0 | 4.5 | 0.4 | 7.9 | 5 |
| C55 | 1 | 11.9 | 1.1 | 9.3 | 2.8 | 0.2 | 5.8 | 3.3 | 0.2 | 6.9 | 5 |
| C56 | 1 | 11.7 | 1.5 | 12.7 | 3.8 | 0.5 | 12.9 | 4.1 | 0.5 | 13.4 | 5 |
| C57 | 1 | 14.3 | 4.1 | 28.7 | 4.6 | 1.1 | 23.2 | 3.9 | 0.4 | 10.7 | 5 |
| C58 | 1 | 11.9 | 0.9 | 7.9 | 3.8 | 0.3 | 8.6 | 4.0 | 0.0 | 0.0 | 5 |
| C59 | 1 | 12.2 | 0.9 | 7.0 | 3.2 | 0.5 | 17.2 | 3.7 | 0.6 | 15.3 | 5 |
| C60 | 1 | 11.2 | 0.4 | 3.2 | 3.0 | 0.4 | 12.4 | 2.4 | 0.1 | 5.3 | 5 |
| 总资源Sum | 11.6 | 2.5 | 21.6 | 3.2 | 0.8 | 25.0 | 3.6 | 0.9 | 25.0 | ||
| 多样性H | 1.02 | 3.73 | 2.98 | 3.15 | 0.62 | ||||||
新窗口打开|下载CSV
月季枝条的节间距大小可以影响月季叶片数量,在一定程度上反映了植株叶片密度。供试月季平均节间距为3.6 cm,品种间变异系数为25%,其中节间距最小的是‘躲躲藏藏’,节间距为1.7 cm,最大的是‘秋日胭脂’,为5.7 cm。其中‘榴花秋舞’和‘永远的艾帕索’枝条抽节均匀,变异系数为0,‘马萨德医生’和‘雏鸡’叶片着生距离差异大,品种内变异系数分别为32.4%和27.5%,大于品种间差异。
供试月季资源均为奇数羽状复叶。月季植株上复叶可以分为两种:一种为中下节处成熟叶片,一种为花梗上部1—2节处的复叶,一般前者小叶数较后者多2片。研究中统计各品种的成熟叶片小叶数,其中小叶数为5的有41个品种,小叶数为7的19个,分别占资源的68%和32%。除两个藤本月季小叶数均为5以外,在其他3种株形的月季中,小叶数5与小叶数7的比例基本保持2﹕1的比例。
综合以上数据可知,引进月季品种以灌木月季为主,月季品种中小叶数5为主要群体,其中叶片最小的月季资源是微型品种‘躲躲藏藏’,其全叶长和顶小叶宽均为最小值;绝大部分月季的叶部形态变化较小,变异系数保持在25.0%及以下。对月季的非花部表型性状进行遗传多样性分析结果显示,遗传多样性指数H变化范围为0.62—3.73(表3)。其中H>1的有株形、全叶长、顶小叶宽和节间距,全叶长的多样性指数最高,为3.73。仅小叶数的多样性最低,为0.62。说明月季植株小叶数在月季育种中较为稳定和单一,而叶片大小受到育种者关注的程度更高。
2.2 月季品种间花朵性状多样性
2.2.1 花色分类月季花色丰富多彩,供试月季资源以纯色花为主,占品种总资源的82%,其中紫色和黄色品种居首,各有10个品种,粉色品种9个,橙色6个,红色和杏色品种各4个,紫红色3个,白色品种2个,绿色仅1个,开发潜力大;非纯色品种中花瓣颜色统一渐变的复色月季有5个,其中‘加百列大天使’为白紫渐变,‘玛丽玫瑰’为粉色白边,‘我爱你’为粉橙渐变;‘香织装饰’和‘甜蜜果酱’为杏粉渐变;条纹品种4个,其中‘彩蝶’为橙白条纹,‘胡里奥’和‘苹果挞’为红白条纹,‘莫奈’为黄红条纹;混色月季的花色差异在于外轮花瓣与内轮花瓣之间,其花色形成值得深入研究,该类月季有2个,‘智慧女神’和‘幻彩’,均为外花瓣白色,内花瓣粉色。
2.2.2 花形态指标多样性
供试月季中单头品种15个,开花时花枝上有且仅有一个花蕾,花量相对较少;39个月季品种开花时呈现出多头特性,占品种总数的65%。这类月季花枝上花朵数虽多,同一花序上花朵仍表现出次第开放,同时期开花量类似单头品种,而持续花量多于单头品种;丰花品种较少,只有6个,花苞多且同时期开花量大,整体效果好。
月季花朵直径平均7.6 cm,花朵最大的为‘卡哈拉’和‘樱桃白兰地’,花朵平均直径为10.5 cm,‘甜蜜马车’花朵平均直径最小,为3.2 cm。花朵直径8.0 cm的月季资源最多,有6个品种,各品种间花朵大小变异系数为25.0%。品种内花朵直径变异较小,仅2个品种变异系数大于20.0%,其中‘魔果’花朵大小最不稳定,品种内变异系数为28.3%,大于品种间差异;而‘桃香’‘安布里奇’‘蝶之舞’‘玛丽玫瑰’‘甜蜜果酱’‘我爱你’‘幻彩’‘欢笑格鲁吉亚’‘皇家’和‘黄金庆典’10个品种的花朵大小性状非常稳定,调查数据几乎完全一致。
花瓣数是月季观赏性状中非常重要的因素之一。供试月季中没有单瓣品种,有13个月季品种花瓣数在40以下,27个品种花瓣数在40—90,20个品种花瓣数在90以上。其中花瓣数最少的是‘新浪潮’,花瓣数平均值为24;花瓣数最多的为‘夏日花火’,花瓣数平均值达203。
月季花型千变万化,形态各异。包子型是近年来市场上备受追捧的月季花型,该类品种在供试月季资源中有20个,占总资源的33%;杯型为月季经典花型,总资源中有19个,占32%;牡丹型品种15个,花型开展形似牡丹,观赏价值高,且大部分花心显露,花药明显且瓣化少,适合月季育种亲本筛选;碗型品种6个,花心多为四分碗或五分碗,其中‘蝶之舞’外层环绕花瓣轮几乎为零,花朵分为四区,酷似蝴蝶展翅。
2.2.3 衰老方式分类
月季花朵的衰老方式是月季观赏价值中最重要的指标之一。田间统计结果显示,供试月季中28个品种的花瓣在枯萎前凋零,而26个月季品种花瓣宿存直至花朵枯萎或花梗萎蔫,6个品种兼有枯萎和落瓣表型,分别占总资源的47%、43%和10%。衰老方式的遗传多样性为0.95。
表4中反映了月季花朵各形态指标的遗传多样性。其多样性指数H分布范围为0.86—3.46。多样性分布从小到大依次为花头数、衰老方式、花瓣数、花型、花色和花朵直径。H>1的性状有花朵直径、花色、花瓣数、花型。由此可见,月季的花朵直径以及花色是开展月季育种的主要方向,而以月季花朵的衰老方式和花头数为目标方向的育种工作开展较少,值得进一步拓展。
Table 4
表4
表4月季品种花朵性状
Table 4
| 品种编号 Code | P6花头数 Bud number | P7 花朵直径 Flower diameter | P8花瓣数 Petal number | P9花色 Color | P10花型 Shape | P11衰老方式 Senescence | ||
|---|---|---|---|---|---|---|---|---|
| 均值Mean (cm) | 标准差SD | 变异系数CV (%) | ||||||
| C1 | 0 | 8.0 | 0.0 | 0.0 | 1 | 5 | 0 | 2 |
| C2 | 1 | 9.1 | 0.3 | 2.7 | 2 | 11 | 2 | 2 |
| C3 | 1 | 6.2 | 0.2 | 2.9 | 1 | 3 | 2 | 0 |
| C4 | 0 | 9.0 | 2.1 | 22.9 | 2 | 8 | 1 | 0 |
| C5 | 1 | 6.0 | 0.3 | 5.4 | 1 | 7 | 0 | 0 |
| C6 | 1 | 9.8 | 0.2 | 2.0 | 2 | 6 | 0 | 0 |
| C7 | 1 | 8.0 | 0.0 | 0.0 | 2 | 1 | 0 | 2 |
| C8 | 0 | 8.6 | 0.6 | 7.4 | 2 | 4 | 1 | 1 |
| C9 | 1 | 4.2 | 0.3 | 7.4 | 1 | 7 | 2 | 0 |
| C10 | 1 | 6.6 | 0.6 | 8.6 | 1 | 7 | 0 | 2 |
| C11 | 1 | 7.4 | 0.5 | 6.6 | 2 | 2 | 3 | 0 |
| C12 | 0 | 9.0 | 0.5 | 6.0 | 1 | 11 | 1 | 1 |
| C13 | 1 | 9.0 | 0.0 | 0.0 | 1 | 6 | 2 | 2 |
| C14 | 0 | 8.5 | 0.0 | 0.0 | 1 | 10 | 1 | 0 |
| C15 | 0 | 9.0 | 0.0 | 0.0 | 1 | 4 | 0 | 2 |
| C16 | 1 | 8.0 | 0.4 | 5.2 | 1 | 6 | 3 | 0 |
| C17 | 1 | 10.0 | 0.0 | 0.0 | 2 | 6 | 0 | 2 |
| C18 | 0 | 10.3 | 0.6 | 6.3 | 2 | 0 | 0 | 0 |
| C19 | 0 | 7.6 | 0.4 | 4.6 | 1 | 9 | 1 | 1 |
| C20 | 1 | 5.8 | 0.2 | 2.6 | 1 | 6 | 0 | 0 |
| C21 | 1 | 6.5 | 0.2 | 2.6 | 1 | 1 | 0 | 0 |
| C22 | 0 | 10.5 | 0.3 | 2.7 | 2 | 7 | 1 | 0 |
| C23 | 1 | 7.5 | 0.7 | 9.7 | 0 | 1 | 0 | 2 |
| C24 | 1 | 9.6 | 0.1 | 1.5 | 1 | 1 | 0 | 0 |
| C25 | 2 | 6.0 | 0.3 | 5.4 | 2 | 4 | 3 | 2 |
| C26 | 1 | 6.6 | 0.3 | 3.8 | 1 | 4 | 2 | 2 |
| C27 | 1 | 6.0 | 0.7 | 11.8 | 0 | 6 | 0 | 0 |
| C28 | 1 | 6.2 | 0.9 | 13.8 | 2 | 5 | 0 | 0 |
| C29 | 2 | 4.5 | 0.7 | 15.7 | 2 | 3 | 1 | 0 |
| C30 | 1 | 9.2 | 0.2 | 2.3 | 0 | 2 | 2 | 2 |
| C31 | 2 | 4.0 | 0.1 | 3.5 | 1 | 4 | 3 | 2 |
| C32 | 0 | 8.0 | 0.0 | 0.0 | 1 | 9 | 2 | 2 |
| C33 | 1 | 4.8 | 0.3 | 6.1 | 1 | 7 | 0 | 0 |
| C34 | 2 | 6.0 | 0.6 | 9.8 | 2 | 4 | 0 | 0 |
| C35 | 1 | 8.5 | 1.5 | 17.3 | 1 | 5 | 1 | 0 |
| C36 | 1 | 5.0 | 1.4 | 28.3 | 0 | 1 | 0 | 0 |
| C37 | 1 | 9.3 | 0.3 | 3.5 | 2 | 11 | 1 | 2 |
| C38 | 1 | 8.5 | 0.3 | 3.5 | 2 | 6 | 3 | 0 |
| C39 | 1 | 10.2 | 0.6 | 6.3 | 2 | 5 | 2 | 2 |
| C40 | 1 | 5.4 | 0.5 | 9.3 | 1 | 11 | 2 | 1 |
| C41 | 1 | 7.5 | 0.5 | 6.6 | 1 | 4 | 2 | 0 |
| C42 | 0 | 9.8 | 0.2 | 1.8 | 0 | 3 | 0 | 2 |
| C43 | 1 | 8.4 | 0.3 | 3.7 | 1 | 4 | 2 | 2 |
| C44 | 0 | 7.5 | 0.0 | 0.0 | 0 | 1 | 0 | 0 |
| C45 | 1 | 5.5 | 0.5 | 9.4 | 2 | 6 | 1 | 2 |
| C46 | 1 | 9.1 | 0.4 | 4.2 | 0 | 2 | 2 | 2 |
| C47 | 0 | 8.0 | 0.0 | 0.0 | 0 | 9 | 1 | 2 |
| C48 | 2 | 3.2 | 0.2 | 6.2 | 0 | 4 | 2 | 2 |
| C49 | 1 | 8.0 | 0.0 | 0.0 | 2 | 9 | 1 | 0 |
| C50 | 1 | 4.6 | 0.1 | 1.9 | 1 | 2 | 1 | 2 |
| C51 | 1 | 8.2 | 0.2 | 2.6 | 1 | 1 | 3 | 1 |
| C52 | 1 | 6.7 | 0.7 | 9.8 | 2 | 9 | 2 | 2 |
| C53 | 2 | 4.9 | 0.5 | 9.6 | 0 | 1 | 2 | 1 |
| C54 | 0 | 9.4 | 0.5 | 5.3 | 0 | 4 | 1 | 2 |
| C55 | 1 | 7.8 | 0.5 | 6.6 | 1 | 1 | 1 | 0 |
| C56 | 1 | 8.9 | 0.9 | 10.0 | 0 | 6 | 1 | 0 |
| C57 | 1 | 10.5 | 0.7 | 6.7 | 2 | 7 | 1 | 2 |
| C58 | 1 | 8.7 | 0.4 | 4.1 | 1 | 6 | 1 | 0 |
| C59 | 0 | 9.8 | 0.4 | 3.7 | 0 | 0 | 1 | 2 |
| C60 | 1 | 5.5 | 0.5 | 9.1 | 1 | 10 | 0 | 0 |
| 总资源 Sum | 7.6 | 1.9 | 25.0 | |||||
| 多样性H | 0.86 | 3.46 | 1.06 | 2.31 | 1.31 | 0.95 | ||
新窗口打开|下载CSV
2.3 性状指标的相关性分析
表5中各性状间相关性分析结果表明,月季不同品种的全叶长与顶小叶宽为极显著(P<0.01)的中度相关,(0.5<等于|r|<0.8),而节间距与全叶长、顶小叶宽极显著低度相关(0.3<等于|r|<0.5),花头数与节间距极显著负相关;月季衰老方式与月季花朵花型之间相关性不显著,但与月季株形之间显著相关;而花头数与花朵直径显著负相关,与小叶数、花型显著正相关。Table 5
表5
表5性状间相关性分析
Table 5
| 性状编号 Index | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 |
|---|---|---|---|---|---|---|---|---|---|---|
| P2 | 0.083 | |||||||||
| P3 | -0.068 | 0.631** | ||||||||
| P4 | -0.069 | 0.459** | 0.392** | |||||||
| P5 | -0.05 | 0.141 | 0.139 | 0.028 | ||||||
| P6 | -0.119 | -0.199 | -0.225 | -0.329** | 0.287* | |||||
| P7 | 0.027 | 0.157 | 0.101 | 0.17 | -0.04 | -0.255 * | ||||
| P8 | 0.22 | 0.05 | -0.001 | -0.105 | -0.012 | 0.119 | -0.119 | |||
| P9 | 0.054 | 0.009 | 0.094 | -0.153 | -0.047 | -0.138 | 0.032 | 0.281* | ||
| P10 | 0.118 | 0.108 | 0.037 | -0.163 | 0.21 | 0.267* | 0.054 | 0.048 | 0.056 | |
| P11 | 0.339** | -0.084 | -0.168 | -0.24 | 0.101 | -0.05 | 0.243 | -0.106 | -0.012 | 0.224 |
Value was correlation coefficient (r). * represented significant correlation (P<0.05). ** represented extremely significant correlation (P<0.01)
新窗口打开|下载CSV
2.4 月季表型性状的分类评价
对11个指标数据进行主成分分析,结果显示前4个主成分累计贡献率达63.4%,代表月季表型性状的大部分信息。将每个表型性状载荷除以其对应成分因子的初始特征值的平方根,得到各成分因子在各性状指标上的特征向量值,各数据结果如表6所示。第一主成分方差贡献率最大,达21.1%,特征值为2.321。第二到第四主成分的方差贡献率分别为15.1%、14.3%和12.9%,对应特征值分别为1.663、1.570和1.417。全叶长、顶小叶宽、节间距在第一主成分中的特征向量绝对值最高,第二主成分中特征向量绝对值高的是花型,第三主成分中特征向量绝对值高的为小叶数和花头数,花瓣数和衰老方式在第四主成分中特征向量绝对值高。花色、花朵直径以及月季株形在4个主成分中在各成分因子中特征向量绝对值均低于0.45,其中月季株形主要与第二、第三主成分有关,花朵直径和花色主要与第四主成分因子有关。Table 6
表6
表6主成分分析各性状的特征向量与贡献率
Table 6
| 性状编号 Index | 第一主成分 First component | 第二主成分 Second component | 第三主成分 Third component | 第四主成分 Fourth component |
|---|---|---|---|---|
| P1 | 0.007 | 0.416 | -0.393 | 0.018 |
| P2 | 0.489 | 0.307 | 0.192 | -0.099 |
| P3 | 0.475 | 0.201 | 0.168 | -0.195 |
| P4 | 0.510 | -0.140 | 0.154 | 0.036 |
| P5 | -0.015 | 0.327 | 0.497 | 0.137 |
| P6 | -0.406 | 0.134 | 0.456 | -0.002 |
| P7 | 0.257 | 0.079 | -0.266 | 0.402 |
| P8 | -0.143 | 0.303 | -0.130 | -0.550 |
| P9 | -0.009 | 0.279 | -0.302 | -0.410 |
| P10 | -0.134 | 0.489 | 0.227 | 0.168 |
| P11 | -0.074 | 0.364 | -0.270 | 0.521 |
| 特征值Eigenvalue | 2.321 | 1.663 | 1.57 | 1.417 |
| 贡献率Percentage (%) | 21.10 | 15.12 | 14.28 | 12.88 |
| 累计贡献率Cumulative percentage (%) | 21.10 | 36.22 | 50.49 | 63.38 |
新窗口打开|下载CSV
月季品种主成分综合得分F值最终计算公式为:F=0.33F1+0.24F2+0.23F3+0.20F4。说明该综合得分数值不能以某一性状为主要参考,需要综合评价月季各表型性状。从图2可以看出,60个品种的主成分综合得分主要集中在-1.878—1.522分,其中综合得分-1.5处、-0.5处和1处附近均有较明显群组区分。根据综合得分图,可以将月季品种划分为如表7所示4个群组。各群组的主成分综合得分平均值为1.186、0.121、-0.798和-1.878。其中群组1有8个品种,占总品种数的13%,它们的主成分综合得分平均值最高。该群组品种叶形较大,花型以碗状为主,花头多而花朵以枯萎为主。其中得分最高的为‘神秘’,1.522分,说明该品种株形、花型和叶形较大。群组2有36个品种,品种数最多,占总品种数的60%,该群组中品种各表型指标均分布范围较大,但综合评价得分高。群组3有15个品种,该群组品种以叶形态中等偏小为主,花头数偏单头,花瓣数较少;群组4有且仅有1个品种,即‘躲躲藏藏’,综合得分-1.878,其株形、叶形到花型均以微小为特色,是非常突出的微型月季资源。
图2
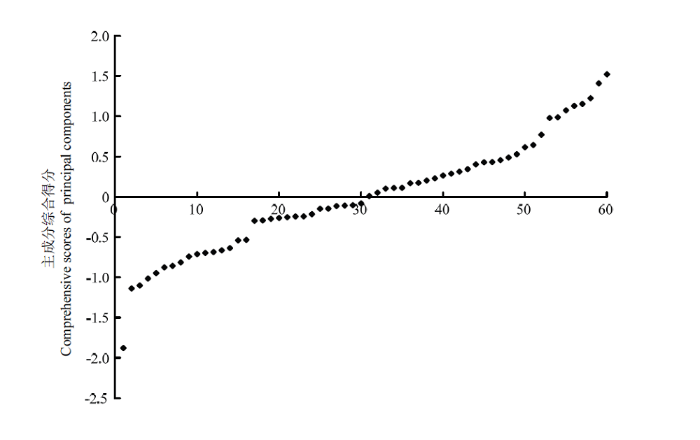 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图260个月季品种主成分综合得分分布
Fig. 2Comprehensive scores of 60 rose cultivars based on the principal component analysis
Table 7
表7
表7月季品种的主成分综合得分分组
Table 7
| 分组 Clusters | 品种数 Amount | 主成分综合得分均值 Average score of principal component | 品种代号 Code of cultivar |
|---|---|---|---|
| 群组1 Cluster 1 | 8 | 1.186 | C54, C11, C30, C13, C23, C16, C38, C43 |
| 群组2 Cluster 2 | 36 | 0.121 | C40, C17, C32, C21, C55, C44, C18, C24, C52, C50, C31, C35, C48, C34, C58, C28, C12, C26, C15, C36, C51, C22, C56, C47, C2, C14, C53, C1, C39, C59, C42, C19, C3, C46, C57, C5 |
| 群组3 Cluster 3 | 15 | -0.798 | C45, C7, C20, C33, C10, C37, C29, C60, C49, C25, C41, C27, C6, C8, C4 |
| 群组4 Cluster 4 | 1 | -1.878 | C9 |
新窗口打开|下载CSV
3 讨论
现代月季品种以四倍体为主,基因组复杂。由于表型与基因型密切相关,受遗传背景的影响,对现代月季品种展开表型的分析研究非常重要。供试的60个月季品种均为现代月季资源,遗传背景不一。各品种在11个表型性状上的遗传多样性信息指数H分布于0.62—3.73,表型研究结果丰富多样。各数量指标的品种间变异系数范围在21%—25%,相比于有关二倍体和四倍体月季的研究结果,本研究中各性状的变化幅度均不大,变异系数范围较集中,且更接近于吴超等[6]和周利君等[9]在四倍体月季中的研究结果,而与KAWAMURA等[10]有关二倍体月季的研究结果差异较大。由此推测供试月季绝大部分为多倍体现代月季,而对于多倍体月季的研究确实不能完全参照二倍体月季模式[32]。月季表型与基因型研究显示某些开花表型与一些非花部表型之间有密切联系。例如月季重复开花特性与直立型特性有连锁现象,而枝条蔓性与一次开花特性连锁[10]等。在对供试月季进行各指标相关性分析时发现,月季花头数与节间距呈极显著负相关,与小叶数呈显著正相关;花朵衰老方式与植株株形显著正相关;花瓣数与花朵花型相关性不大,与花色呈显著正相关。但由于各组相关性系数偏低(|r|<0.5),说明这种表型上的相关性虽然有关联,但关联性不强,还需要后期通过在更多的月季材料上收集数据加以验证。
花是月季最重要的观赏部位,花部性状指标对月季品种的观赏价值定位和分类起决定作用。叶部性状不但与月季的观赏特性有关,还参与其光合能力以及抗性。本研究对月季11个表型性状进行主成分分析显示,前4个主成分方差贡献率达63.28%,可以作为月季评价的主要指标。而这4个成分指标依次侧重于叶型、花型、花头数、花瓣数及花朵衰老方式等性状。与吴超等[6]对国外微型月季的主成分分析结果类似,但次序上有差异。究其原因,可能与性状变量的选择密切相关。由于主成分分析法侧重于降维,本研究中花部各性状变量间相关程度均不高,而叶部指标三变量间相关度较好(表5),因此最终获得的主成分分析结果中花部特征依然比较离散,而叶部仅在第一主成分中有突出贡献率。因此,在月季品种性状调查中,还应多开发叶部或其他非花部性状指标,以便全面综合评价月季品种。
近年来,国内外月季育种事业取得了丰硕成果[33]。我国自育月季目标主要分为花色育种、花香育种、抗病育种和抗性育种4类[34],而以其他表型为育种目标开展的工作较少。例如,与王镭等[2]分类结果一致,本研究中发现月季花朵自然衰老方式中落瓣与枯萎的品种数几乎均等。未来可以在月季育种过程中有效利用和改进这种多样性。此外,本研究中发现的特异微型月季、独特复色花色资源以及特异花型资源等也将为月季在不同育种方向上的创新提供材料。
4 结论
现代月季形态多样,花色丰富,尤其以叶型大小居中、5小叶、花型紧包、重瓣性强、花头多且持续开放的灌木型纯色花月季品种为主流。月季叶部和花部表型性状多而复杂,部分花形态指标与叶形态或生长形态显著相关。主成分分析结果表明,叶型与花型是评价月季性状非常重要的两个因子。通过主成分综合得分能够合理分类月季品种,突显特殊资源。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 1]
In: VAN HUYLENBROECK J.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}