Genetic Diversity of Common Millet Resources Assessed with EST-SSR Markers
SHI TianTian1, HE JieLi2, GAO ZhiJun3, CHEN Ling4, WANG HaiGang4, QIAO ZhiJun,4, WANG RuiYun,1,41 College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi 2 College of Arts and Sciences, Shanxi Agricultural University, Taigu 030801, Shanxi 3 Erdos Institute of Agriculture and Animal Husbandry, Erdos 017000, Inner Mongolia 4 Institute of Crop Germplasms Resources of Shanxi Academy of Agricultural Sciences/Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture, Taiyuan 030031
Abstract 【Objective】The objective of this study is to analyze the genetic diversity and relationship of common millet accessions (six different ecotopes at home and abroad) by microsatellite markers, to provide available data for classification, selection and utilization of elite germplasm resources. 【Method】 One hundred and forty-four SSRs (64 high motif nucleotide sequence repeat and 80 low ones) are used to identify polymorphisms in ninety-six common millet accessions (71 home accessions and 25 abroad accessions). Genetic diversity parameters were calculated using software PowerMarker 3.25 and PopGen 1.32. Genetic distance and Structure on accessions were classified with software MEGA 5.0 and Structure 2.2, respectively. PCA (principal component analysis) was conducted by software Ntsys 2.11. 【Result】 Using 144 EST-SSR markers, a total of 368 observed alleles (Na) were detected with 2-3 alleles (mean = 2.5556) per locus. The observed heterozygosity (Ho) ranged from 0.4070 (RYW15) to 0.9789 ( RYW85) with an average of 0.8288. The expected heterozygosity (He) ranged from 0.4369 ( RYW59) to 0.6693 ( RYW58) with an average of 0.5535. The Nei's gene diversity index ranged from 0.4344 (RYW59) to 0.6653 (RYW58) with a mean of 0.5505. The polymorphism information content (PIC) ranged from 0.1811 (RYW68) to 0.7508 (RYW58) with an average of 0.4279. The Shannon diversity index (I) range was 0.6474 to 1.0956 with an average of 0.8415. In the case of genetic diversity parameters, the PIC and I of common millet accessions from Northern spring-sowing region were the most abundant than that of others. On the contrary, the accessions from Northwest spring & summer-sowing region were the lowest. For the different geographical regions accessions, the range of genetic distance was 0.0111 to 0.1425 and the scope of genetic consistency was 0.8672 to 0.9889. The genetic distance of accessions between Loess Plateau spring & summer-sowing region and Northern spring-sowing region was the least and their genetic consistency was the highest. The genetic distance between Northwest spring & summer-sowing region and North of China summer-sowing region was the largest and their genetic consistency was the lowest. UPGMA analysis divided 96 accessions into three groups (GroupⅠ, Ⅱ and Ⅲ). Group I were mainly Northern spring-sowing region common millet, Group Ⅱ were mainly foreign accessions, and Group Ⅲ were mainly Loess Plateau spring & summer-sowing region and Northern spring-sowing region accessions. Structure cluster divided resources into four groups. The red group contained Loess Plateau spring & summer-sowing region and Northern spring-sowing region accessions, which represented the gene pool of Loess Plateau and North. The green group included Northern spring-sowing region accessions, which represented the North gene pool. The blue group were Loess Plateau spring & summer-sowing region accessions, which represented the gene pool of Loess Plateau. The yellow group represented foreign gene pool. In terms of diversity parameters, the PIC value of GroupⅡwas the highest at 0.4606 and the Group Ⅳ was the lowest at 0.3539. The PCA analysis classified all accessions into six clusters, which are related to their geographical region. 【Conclusion】144 SSRs can evaluate 96 common millet resources accurately. The genetic relationships are related to their geographical region. Genetic diversity of accessions from Northern spring-sowing region are more abundant. Keywords:common millet;SSR;cluster analysis;genetic structure;principal component analysis
PDF (3288KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 石甜甜, 何杰丽, 高志军, 陈凌, 王海岗, 乔治军, 王瑞云. 利用EST-SSR评估糜子资源遗传差异[J]. 中国农业科学, 2019, 52(22): 4100-4109 doi:10.3864/j.issn.0578-1752.2019.22.014 SHI TianTian, HE JieLi, GAO ZhiJun, CHEN Ling, WANG HaiGang, QIAO ZhiJun, WANG RuiYun. Genetic Diversity of Common Millet Resources Assessed with EST-SSR Markers[J]. Scientia Acricultura Sinica, 2019, 52(22): 4100-4109 doi:10.3864/j.issn.0578-1752.2019.22.014
0 引言
【研究意义】糜子(Panicum miliaceum L.)属禾本科黍属,又称黍稷或糜黍,抗旱耐瘠,在中国、俄罗斯、印度和美国主要用作粮食和饲料[1,2,3]。随着全球气温日益变暖、未来环境渐趋干旱,作为哺育中华民族的载体作物和战略储备作物,糜子在保障国际粮食安全中起重要作用[4]。了解糜子种质现状,准确评估资源遗传背景,有助于加速糜子优异种质的发掘,并为其高效利用提供理论基础。【前人研究进展】糜子分布广,种植历史超过一万年,是中国北方冷凉地区的主粮,也是美国大平原小麦旱作农业系统中的轮作作物[5,6]。在印度国际半干旱热带作物研究所(International Crops Research Institute for the Semi-Arid Tropics,ICRISAT)和中国农业科学院种质资源研究所国家种质资源库(Institute of Crop Germplasm Resources, Chinese Academy of Agricultural Sciences,ICGR- CAAS)分别保存有29 000余份和9 885份糜子[2,3]。首先,基于表型性状调查,发现糜子农艺性状遗传多样性丰富。2008年,对8 016份(来自ICGR-CAAS)糜子资源的11个农艺性状进行主成分和聚类分析,结果归纳为8个主成分,聚为5个群组,群组4综合性状表现较好[7]。2016年,调查8 515份糜子资源(来自ICGR-CAAS)的7个农艺性状,发现株高为80—200 cm,野生型偏低,农家种偏高;落粒性程度(轻度<5%,中度6%—10%,重度>11%)的分布呈区域性差异,轻度落粒性和株高较高(高产潜势)存在协同选择效应(植株越矮,落粒程度越严重)[8]。2019年,调查849份糜子资源(来自ICRISAT)的18个农艺性状,发现播种至开花的时间为26—50 d,株高为20—133 cm(平均62 cm),穗长为22—400 mm(平均209 mm)[2]。第二,基于基因型检测,鉴定到糜子资源存在诸多遗传差异。简单重复序列(simple sequence repeat,SSR)在基因组中含量丰富、多态性高、重复性好,是研究群体遗传变异常用的分子检测工具,已广泛用于高粱等作物的遗传多样性评估[9,10,11,12,13]。2009年,用46个糜子非特异性SSR分析118份糜子资源(中国110份、美国6份和俄罗斯2份),发现黄土高原生态栽培区材料的遗传多样性最丰富[14]。2010年,基于基因组DNA富集SSR文库,25个糜子特异性微卫星标记首次被开发[15]。2011年,用其中16个标记分析98份欧亚大陆糜子资源,发现基于遗传距离划分的东部和西部2个群组与地理来源相符,东部类群进一步划分为4个亚类群或基因库,而西部类群划分为2个亚类群。东部群组的材料来源于中国、蒙古、尼泊尔、印度东北部、俄罗斯远东地区、韩国和日本,西部群组的材料来源于乌克兰、俄罗斯北高加索、俄罗斯欧洲大陆部分、中亚、印度西北部、巴基斯坦、中国和蒙古[16]。2015年,用其中19个标记分析来自7个地理群组的96份(95份来自中国6个省,1份来自俄罗斯)糜子资源,发现山西资源的遗传多样性最丰富[17]。2017年,对其中的15个标记标注荧光后,检测来源于中国11个省/自治区的132份糜子资源,获得107个等位变异,平均每个位点为7个,中国资源来自4个基因库(东北地区、黄土高原、北方地区和西北地区),其丰富的遗传变异主要存在于材料间[18]。2018年,用其中16个SSR和2个GBSSI标记分析341份欧亚大陆糜子资源(195份来自中国),发现黄土高原西部为糜子的主要驯化地[19]。近年来,利用转录组测序手段,构建出一批糜子中开发的EST-SSR,基于这些标记进行了一系列糜子遗传多样性研究。2016年,利用67个SSR分析来自中国7个糜子主产区的88份糜子资源,检测到179个等位变异,基于遗传距离将试材聚为4个群组[20]。同年,利用63个SSR评估来自国内外10个不同地理来源群体的192份糜子资源,检测到161个等位变异,平均PIC值为0.4855,发现内蒙、东北和黄土高原地区种质资源遗传多样性最丰富[21]。2017年,基于85个高基元SSR标记扩增中国6个糜子生态栽培区的96份糜子资源,检测到232个等位变异,发现黄土高原春夏糜子区和北方春糜子区资源遗传多样性最丰富[22]。2018年,用103个SSR评估国内外12个群体的146份糜子资源(85份国内野生资源和61份国外地方品种),检测到308个等位变异,PIC为0.5544,发现中国河北群体的遗传多样性最丰富[23];对雁黍5号进行高通量测序,挖掘到8 139个全基因组EST-SSR,这些标记有望用于农艺性状定位、连锁图谱构建和分子育种等遗传研究[24]。【本研究切入点】丰富的糜子资源在旱作生态农业建设中起重要作用,而搞清其遗传背景是合理利用的前提。但作为小宗作物,其遗传多样性评估研究偏少,亟待深入开展。【拟解决的关键问题】本研究利用144个糜子中开发的EST-SSR标记评估96份国内外糜子资源,搞清不同生态区资源的遗传差异,为糜子优异种质的鉴定和筛选提供理论依据。
▲国外Abroad;◇华北夏糜子区Northern Summer-sowing ecotope;▉西北春夏糜子区Northwest spring & summer-sowing ecotope;□北方春糜子区Northern spring-sowing ecotope;●黄土高原春夏糜子区Loess plateau spring & summer-sowing ecotope;○东北春糜子区Northeast Spring-sowing ecotope Fig. 1Cluster diagram of 96 accessions of common millet
ZHANG LZ, JI PS, LUO XX, MU HZ, QIN YP, NIUY, QIAO ZJ . Physicochemical properties and digestive properties of millet starch Food Science, 2016,37(19):76-81. (in Chinese) [本文引用: 1]
VETRIVENTHANM, AZEVEDO V CR, UPADHYAYA HD, NARESHD . Variability in the global proso millet ( Panicum miliaceum L.) germplasm collection conserved at the ICRISAT Genbank Agriculture, 2019,9(5):112 [本文引用: 3]
WANG RY. Genetic Diversity and Evolution Advancement in Common Millet (Panicum miliaceum L.). Beijing: China Agriculture Press, 2017: 2-3. (in Chinese) [本文引用: 2]
DIAO XM, CHENG RH . Current breeding situation of foxtail millet and common millet in China as revealed by exploitation of 15 years regional adaptation test data Scientia Agricultura Sinica, 2017,50(23):4469-4474. (in Chinese) [本文引用: 1]
LU HY, ZHANG JP, LIU KB, WU NQ, LI YM, ZHOU KS, YE ML, ZHANG TY, ZHANG HJ, YANG XY, SHEN LC, XU DK, LIQ . Earliest domestication of common millet ( Panicum miliaceum) in East Asia extended to 10,000 years ago Proceedings of the National Academy of Sciences of the United States of America, 2009,106(18):7367-7372. [本文引用: 1]
RAJPUT SG, TAMMY PH, DIPAK KS . Development and characterization of SSR markers in proso millet based on switchgrass genomics American Journal of Plant Sciences, 2014,5(1):175-186. [本文引用: 1]
HU XY, LUP, HE JB, WANGL, WANG YX, ZHANG HS, ZHANG ZW, WUB . Principal components and cluster analysis of agronomic traits of proso millet ( Panicum miliaceum) Journal of Plant Genetic Resources, 2008,9(4):492-496. (in Chinese) [本文引用: 1]
WANG RY, HUNT HV, QIAO ZJ, WANGL, HAN YH . Diversity and cultivation of broomcorn millet ( Panicum miliaceum L.) in China: A Review Economic Botany, 2016,70(3):332-342. [本文引用: 1]
ELEUCHL, JILALA, GRANDOS, CECCARELLIS, SCHMISING M VK, TSUJIMOTOH, HAJERA, DAALOULA, BAUMM . Genetic diversity and association analysis for salinity tolerance, heading date and plant height of barley germplasm using simple sequence repeat markers Journal of Integrative Plant Biology, 2008(8):1004-1014. [本文引用: 1]
WEN ZX, DING YL, ZHAO TN, GAI JY . Genetic diversity and peculiarity of annual wild soybean ( G. soja Sieb. et Zucc.) from various eco-regions in China Theoretical and Applied Genetics, 2009,119(2):371-381. [本文引用: 1]
LI RY, ZHANGH, ZHOU XC, GUAN YA, YAO FX, SONG GA, Wang JC, ZHANG CQ . Genetic diversity in Chinese sorghum landraces revealed by chloroplast simple sequence repeats Genetic Resources and Crop Evolution, 2010,57(1):1-15. [本文引用: 1]
BADIANE FA, GOWDA BS, CISSN, DIOUFD, SADIOO, TIMKO MP . Genetic relationship of cowpea ( Vigna unguiculata) varieties from Senegal based on SSR markers Genetics and Molecular Research, 2012,11(1):292-304. [本文引用: 1]
WANG CF, JIA GQ, ZHIH, NIU ZG, CHAIY, LIW, WANG YF, LI HQ, LUP, ZHAO BH, DIAO XM . Genetic diversity and population structure of Chinese foxtail millet [Setaria italica (L.) Beauv.] landraces. Genes Genomes Genetics, 2012,2(7):769-777. [本文引用: 1]
HU XY, WANG JF, LUP, ZHANG HS . Assessment of genetic diversity in broomcorn millet ( Panicum miliaceum L.) using SSR markers Journal of Genetics and Genomics, 2009,36(8):491-500. [本文引用: 1]
CHO YI, CHUNG JW, LEE GA, MA KH, DIXITA, GWAG JG, PARK YJ . Development and characterization of twenty-five new polymorphic microsatellite markers in proso millet ( Panicum miliaceum L.) Genes &Genomics, 2010,32(3):267-273. [本文引用: 2]
HUNT HV, CAMPANA MG, LAWES MC, PARK YJ, BOWER MA, HOWE CJ, JONES MK . Genetic diversity and phylogeography of broomcorn millet ( Panicum miliaceum L.) across Eurasia Molecular Ecology, 2011,20(22):4756-4771. [本文引用: 1]
DONG JL, WANG HG, CHENL, WANG JJ, CAO XN, WANGL, QIAO ZJ . Analysis of genetic diversity and structure of proso millet core germplasm Scientia Agricultura Sinica, 2015,48(16):3121-3131. (in Chinese) [本文引用: 1]
WANG RY, JIX, LUP, LIU MX, XUY, WANGL, WANG HG, QIAO ZJ . Analysis of genetic diversity in common millet ( Panicum miliaceum) using fluorescent SSR in China Acta Agronomica Sinica, 2017,43(4):530-548. (in Chinese) [本文引用: 1]
HUNT HV, RUDZINSKIA, JIANG HE, WANG RY, THOMAS MG, JONES MK . Genetic evidence for a western Chinese origin of broomcorn millet ( Panicum miliaceum) The Holocene, 2018,28(12):1968-1978. [本文引用: 1]
LIU MX, XUY, HE JH, ZHANGS, WANG YY, LUP . Genetic diversity and population structure of broomcorn millet ( Panicum miliaceum L.) cultivars and landraces in China based on microsatellite markers International Journal of Molecular Sciences, 2016,17(3):370. [本文引用: 1]
LIANS, LUP, QIAO ZJ, ZHANGQ, ZHANGQ, LIU MX, WANG RY . Genetic diversity in broomcorn millet ( Panicum miliaceum L.) from China and abroad by using SSR markers Scientia Agricultura Sinica, 2016,49(17):3264-3275. (in Chinese) [本文引用: 2]
WANG RY, LIU XY, WANG HG, LUP, LIU MX, CHENL, QIAO ZJ . Evaluation of genetic diversity of common millet ( Panicum miliaceum) germplasm available in China using high motif nucleotide repeat SSR markers Scientia Agricultura Sinica, 2017,50(20):3848-3859. (in Chinese) [本文引用: 1]
XUE YT, LUP, QIAO ZJ, LIU MX, WANG RY . Genetic diversity and genetic relationship of broomcorn millet ( Panicum miliaceum L.) germplasm based on SSR markers Scientia Agricultura Sinica, 2018,51(15):2846-2859. (in Chinese) [本文引用: 2]
JIANG YM, Li HQ, ZHANG JN, XIANG JY, CHENG RH, LIU GQ . Whole genomic EST-SSR development based on high-throughput transcript sequencing in proso millet ( Panicum miliaceum) International Journal of Agriculture & Biology, 2018,20(3):617-620. [本文引用: 3]
LIUK, MUSE SV . PowerMarker: Integrated analysis environment for genetic marker data Bioinformatics, 2005,21:2128-2129. [本文引用: 1]
YEH FC, BOYLE TJ . Population genetic analysis of co-dominant and dominant markers and quantitative traits Belgian Journal of Botany, 1997,130:129-157. [本文引用: 1]
TAMURAK, PETERSOND, PETERSONN, STECHERG, NEIM, KUMARS . MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods Molecular Biology and Evolution, 2011,28(10):2731-2739. [本文引用: 1]
FALUSHD, STEPHENSM, PRITCHARD JK . Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies Genetics, 2003,164(4):1567-1587. [本文引用: 1]
NEIM, LI WH . Mathematical model for studying genetic variation in terms of restriction endonucleases Proceedings of the National Academy of Sciences of the United States of America, 1979,76(10):5269-5273. [本文引用: 1]
BONMAN JM, BABIKER EM, CUESTA-MARCOSA, ESVELT-KLOSK, BROWN-GUEDIRAG, CHAOS, SEED, CHENJ, AKHUNOVE, ZHANGJ, BOCKELMAN HE, GORDON TC . Genetic diversity among wheat accessions from the USDA national small grains collection Crop Science, 2015,55(3):1243-1253. [本文引用: 1]
COURTOISB, FROUINJ, GRECOR, BRUSCHIG, DROCG, HAMELINC, RUIZM, CLéMENTG, EVRARD JC, VAN COPPENOLES, KATSANTONISD, OLIVEIRAM, NEGR?OS, MATOSC, CAVIGIOLOS, LUPOTTOE, PIFFANELLIP, AHMADIN . Genetic diversity and population structure in a European collection of rice Crop Science, 2012,52(4):1663-1675. [本文引用: 1]
VAN INGHELANDTD, MELCHINGER AE, LEBRETONC, STICHB . Population structure and genetic diversity in a commercial maize breeding program assessed with SSR and SNP markers Theoretical and Applied Genetics, 2010,120(7):1289-1299. [本文引用: 1]
WANG ST, HE JL, SHI TT, CHENL, WANG HG, WANG RY, QIAO ZJ . Genetic diversity analysis of broomcorn millet ( Panicum miliaceum L.) of Shanxi Province using microsatellite markers Journal of Plant Genetic Resources, 2019,20(1):69-78. (in Chinese) [本文引用: 1]
LIU XY . Analysis of genetic diversity of Chinese proso millet resources by high substrate SSR [D]. Taigu: Shanxi Agricultural University, 2017. (in Chinese) [本文引用: 3]
ZHANG YD, PENGC, LI ZF, YANG YL, HU XY . Genetic diversity of genomic-SSR and EST-SSR markers in interspecies of poplar Journal of Northeast Forestry University, 2011 39(12):8-11, 117. (in Chinese) [本文引用: 1]
XUY, CHEN JH, LIY, HONGZ, WANGY, ZHAO YQ, WANG XM, SHI JS . Development of EST-SSR and genomic-SSR in Chinese fir Journal of Nanjing Forestry University (Natural Sciences Edition), 2014,38(1):9-14. (in Chinese) [本文引用: 1]
CHO YG, ISHIIT, TEMNYKHS, CHENX, LIPOVICHL, MCCOUCH SR, PARK WD, AYRESN, CARTINHOURS . Diversity of microsatellites derived from genomic libraries and GenBank sequences in rice ( Oryza sativa) Theoretical and Applied Genetics, 2000,100(5):713-722. [本文引用: 1]
EUJAYLI, SORRELLSM, BAUMM, WOLTERSP, POWELLW . Assessment of genotypic variation among cultivated durum wheat based on EST-SSRs and genomic SSRs Euphytica, 2001,119(1):39-43. [本文引用: 1]
TUSKAN GA, GUNTER LE, YANG, YANG ZK, YIN TM, SEWELL MM, DIFAZIO SP . Characterization of microsatellites revealed by genomic sequencing of Populus trichocarpa. Canadian Journal of Forest Research-Revue, 2004,34(1):85-93. [本文引用: 1]
DUTTAS, KUMAWATG, SINGH BP, GUPTA DK, SINGHS, DOGRAV, GAIKWADK, SHARMA TR, RAJE RS, BANDHOPADHYA TK, DATTAS, SINGH MN, BASHASABF, KULWALP, WANJARI KB, VARSHNEY RK, COOK DR, SINGH NK . Development of genic-SSR markers by deep transcriptome sequencing in pigeonpea [Cajanus cajan (L.) Millspaugh]. Biomed Central Plant Biology, 2011,11(1):17. [本文引用: 1]
 ,4, 王瑞云
,4, 王瑞云
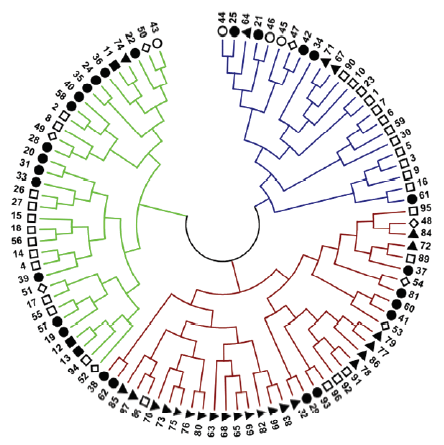 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT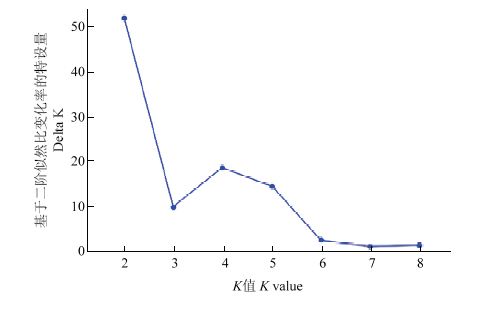 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT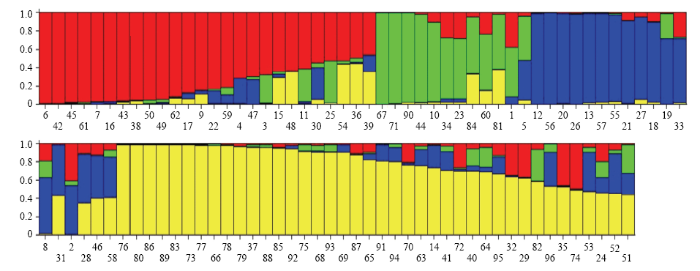 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT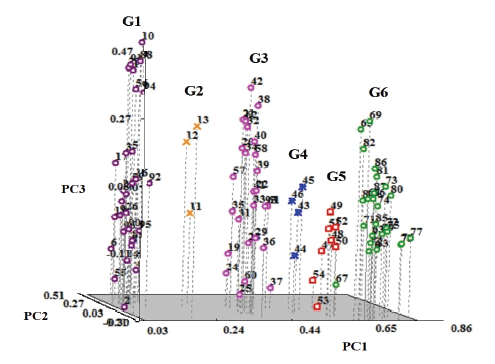 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}