 ,甘肃农业大学农学院/甘肃省干旱生境作物学重点实验室/甘肃省油菜工程技术研究中心/甘肃省作物遗传改良与种质创新重点实验室,兰州 730070
,甘肃农业大学农学院/甘肃省干旱生境作物学重点实验室/甘肃省油菜工程技术研究中心/甘肃省作物遗传改良与种质创新重点实验室,兰州 730070Comprehensive Assessment on Cold Tolerance of the Strong Winter Brassica napus L. Cultivated in Northern China
PU YuanYuan, ZHAO YuHong, WU JunYan, LIU LiJun, BAI Jing, MA Li, NIU ZaoXia, JIN JiaoJiao, FANG Yan, LI XueCai, SUN WanCang,College of Agronomy, Gansu Agricultural University/Gansu Provincial Key Laboratory of Arid Land Crop Sciences/Gansu Research Center of Rapeseed Engineering and Technology/Key Laboratory of Crop Genetics Improvement and Germplasm Enhancement of Gansu Province, Lanzhou, 730070通讯作者:
责任编辑: 李莉
收稿日期:2019-04-19接受日期:2019-05-30网络出版日期:2019-10-01
| 基金资助: |
Received:2019-04-19Accepted:2019-05-30Online:2019-10-01
作者简介 About authors
蒲媛媛,E-mail:vampirepyy@126.com。

摘要
关键词:
Abstract
Keywords:
PDF (6346KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
蒲媛媛, 赵玉红, 武军艳, 刘丽君, 白静, 马骊, 牛早霞, 金姣姣, 方彦, 李学才, 孙万仓. 北方强冬性甘蓝型冬油菜品种(系)抗寒性评价[J]. 中国农业科学, 2019, 52(19): 3291-3308 doi:10.3864/j.issn.0578-1752.2019.19.002
PU YuanYuan, ZHAO YuHong, WU JunYan, LIU LiJun, BAI Jing, MA Li, NIU ZaoXia, JIN JiaoJiao, FANG Yan, LI XueCai, SUN WanCang.
0 引言
【研究意义】甘蓝型油菜原产欧洲,1943年由原浙江大学孙逢吉教授从日本引进中国,1955年引入陕西汉中地区,此后逐渐北移至陕西关中一带,并逐步发展到甘肃天水以南的川水区[1,2]。甘蓝型冬油菜具有产量高、抗倒伏、品质优良等优良性状,在中国油菜生产中处于主导地位,总产量占全国油菜的86.3%[3]。由于抗寒性差,甘蓝型冬油菜主要分布于北纬34o以南,海拔约1 100 m、冬季负积温-200℃以下的地区[2]。因此,研究甘蓝型冬油菜的抗寒性及相应的抗寒性鉴定方法,筛选适宜于中国北方种植的优良抗寒种质,对中国西北甘蓝型冬油菜产业发展至关重要。【前人研究进展】甘蓝型冬油菜抗寒性改良研究备受关注,雷建明等[4]育成了强冬性甘蓝型杂交油菜天油9号,在甘肃天水中梁(北纬34°61′,海拔1 650 m)越冬率达到95%以上。黄继英等[5]提出,甘蓝型冬油菜北移应选用耐冻性强,冬前苗期形态为匍匐或半直立生长的品种。宋图良[6]、史鹏辉[7]、马骊等[8]研究表明,抗寒性强的甘蓝型冬油菜具有苗期叶色深绿微带紫红色,叶柄短,半直立生长,进入越冬期,叶片生长习性趋于匍匐生长等特点。植物的抗寒性与生理变化有着密切的关系,杜春芳[9]研究发现,耐寒品系L9比敏感品系L7响应低温胁迫(4℃)更迅速,随着低温胁迫时间的延长,L9相对电导率逐渐下降,恢复到正常水平,表现出较强的耐寒性和适应性,而L7的相对电导率则逐渐升高,在12—24 h急剧上升,细胞膜发生了较严重的破坏,电解质大量外渗,表现出较差的耐寒性和适应性。耐寒品系L9低温胁迫下(4℃),脯氨酸积累明显高于敏感品系L7,L9的可溶性糖含量的明显变化比L7灵敏,耐寒性和适应性更强。王月等[10]研究表明甘蓝型冬油菜北移后,叶片中SOD、POD和CAT酶活性及可溶性蛋白含量都有所升高。宋良图[6]通过对甘蓝型油菜越冬过程的生理研究发现,秦油3号越冬前可溶性糖含量较低,最低值为6.58%,随着气温降低,可溶性糖含量提高较快,达26%以上。目前,越冬率[11]和冻害指数[12]为冬油菜抗寒评价最常用的指标。闫蕾等[13]通过低温处理油菜幼苗,统计存活率,计算抗寒指数,来评估油菜的抗寒性强弱。杨宁宁等[14]通过统计田间越冬率、测定其半致死温度(lethal temperature,LT50)及应用隶属函数分析等方法,对7个冬油菜的抗寒性进行了综合评价,LT50测定结果和隶属函数分析结果均与田间越冬率的结果一致。李瑞雪等[15]用电导法和隶属函数法对6种含笑属植物的抗寒性进行了评价,其结果与田间调查结果基本一致。迄今为止,隶属函数广泛应用于许多物种的抗逆性评价,如,芍药[16]耐热性评价,菊芋[17]、花生[18]、老芒麦[19]耐盐性评价及种质筛选,观鹅草属[20]、紫花苜蓿[21]耐寒性评价,绿豆[22]耐旱种质筛选及综合评价,花生幼苗[23]耐酸性综合评价,油菜苗期耐铝的鉴定[24],种子萌发期抗旱性评价[25]和耐低温评价[26]等。春化是指冬性作物必须经历一段时间的持续低温才能由营养生长阶段转入生殖生长的现象,是通过低温处理获得或加速植物开花的过程[27]。根据春化所需日照时长和春化时间的不同,将油菜分为冬性、春性、半冬性3种类型。十字花科植物冬季暴露在低温环境中,延缓从营养生长向生殖生长转变的过程,同时获得了抗寒能力[28]。FOWLER等[29]发现4℃(16 h光照)条件下持续处理到冬小麦春化饱和时,其抗寒性最强。【本研究切入点】甘蓝型冬油菜品种抗寒性得到改良,种植区虽然已经由陕西关中一带北移到北纬34°左右的甘肃省天水市秦州区以南地区,但在天水秦州周边地区,仍然以沟播和冬苫栽培技术来保障甘蓝型冬油菜的越冬,因此,甘蓝型冬油菜在北纬34°至北纬35°左右地区,仍然难以越冬。2000年起,甘肃农业大学等针对北方甘蓝型冬油菜越冬问题,进行了抗寒性品种改良研究,初步育成16TS 309-4、16TS 306-3、16TS 309-10、15NS 45-4、2016 8(G)、2016TS(G)10等一批具有优异抗寒性的甘蓝型冬油菜新品系。【拟解决的关键问题】本研究选择甘肃农业大学选育的9个可在北纬35°左右越冬的冬性甘蓝型冬油菜、2个天水农业科学院选育的材料(天油14和天油2288)及新疆农业科学院选育的新油23为材料,对其越冬率、LT50、冬前植株形态、低温下生理生化响应、冬油菜春播后春化率差异进行研究,旨在对冬性甘蓝型冬油菜品种(系)的抗寒性进行划分,提出鉴定冬性甘蓝型冬油菜抗寒性的方法,为强抗寒甘蓝型冬油菜品种选育提供依据。1 材料与方法
1.1 材料及试验地点概况
试验以12个甘蓝型冬油菜为材料,分别在天水市农业科学研究所试验田(天水中梁站)、甘肃农业大学试验田(兰州)、上川镇(兰州永登县)的油菜工程中心试验基地进行,参试材料名称、来源及试验点信息见表1和表2。Table 1
表1
表1试验材料一览表
Table 1
| 品种(系) Variety (line) | 类型 Type | 选育单位 Source |
|---|---|---|
| 新油23 Xinyou23 | 甘蓝型冬油菜 Winter Brassica napus | 新疆农业科学院 Xinjiang Academy of Agricultural Sciences |
| 16TS 309-4 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 16TS 306-3 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 16TS 312-2 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 16TS 309-10 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 16NPZ 269-1 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 16NTS 158 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 15NS 45-4 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 2016 8(G) | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 2016TS(G)10 | 甘蓝型冬油菜 Winter Brassica napus | 甘肃农业大学 Gansu Agricultural University |
| 天油14 Tianyou14 | 甘蓝型冬油菜 Winter Brassica napus | 天水市农业科学研究所 Tianshui Agricultural Science Research Institute |
| 天油2288 Tianyou2288 | 甘蓝型冬油菜 Winter Brassica napus | 天水市农业科学研究所 Tianshui Agricultural Science Research Institute |
新窗口打开|下载CSV
Table 2
表2
表2种植区主要气候因子
Table 2
| 环境因子 Environment factor | 天水 Tianshui | 上川 Shangchuan | 兰州 Lanzhou |
|---|---|---|---|
| 经度 Longitude | 105°69′ E | 103°40′ E | 103°73′ E |
| 纬度 Latitude | 34°60′ N | 36°03′ N | 36°73′ N |
| 海拔 Altitude (m) | 1084—1650 | 2150 | 1517 |
| 年平均气温 Annual average temperature (℃) | 11.5 | 6.5 | 10.3 |
| 最冷月平均最低气温 Lowest temperature in the coldest month (℃) | -2.0 | -14.6 | -5.0 |
| 极端气温 Extreme low temperature (℃) | -14.7 | -26.5 | -16.0 |
| 无霜期 Free frost days (d) | 155 | 142 | 180 |
| 平均降雨量 Precipitation (mm) | 491 | 175 | 327 |
新窗口打开|下载CSV
1.2 试验设计
试验分秋播和春播2部分。秋播:试验于2016—2018年分别在天水(渭南,海拔1 084 m;中梁,海拔1 650 m)和上川镇油菜工程中心试验基地进行。小区面积4 m2,3次重复,随机区组排列,开沟条播,行距20 cm,株距8—10 cm,天水于9月中旬播种,上川于8月中旬播种,出苗后及时定苗,越冬前、返青后统计苗数,计算越冬率,越冬率(%)=返青期植株数/苗期植株总数×100%。2017年为提高越冬率,冬前分别对甘肃农业大学试验点和上川镇试验点种植的材料进行地膜覆盖。
春播:试验于2017—2018年3月下旬播种于上川镇油菜工程中心试验基地,小区面积:4 m2,3次重复,随机区组排列,开沟条播,行距20 cm,株距8—10 cm。8月下旬统计小区总株数及处于不同生长发育阶段(主要包括未现蕾、现蕾期、花期和成熟期)的株数,计算春化率,春化率(%)=未现蕾的植株数/小区总株数×100%,分别测量每个植株的株(薹)高。
采样:2016年分别于9月26日(气温:7—18℃)和10月29日(气温:0—8℃)在上川镇油菜工程中心试验基地进行田间采样。挖取整个植株用于测定地上、地下干物质。取每株植株展开的第三片叶,用于测定生理指标。
1.3 测定项目及方法
1.3.1 半致死温度(LT50) 2016年9月上旬,播种于树脂花盆中(直径20 cm),置于自然条件下生长(甘肃农业大学,兰州),待植株长至6、7叶,分别置于5℃、0℃、-5℃、-10℃和-15℃各处理3 h,取每株植物展开的第三片叶,用直径约8 mm的打孔器,避开主叶脉打叶圆片,将叶圆片装入15 mL带盖的离心管中,每个温度下3个重复,每个离心管放10片圆叶,加入10 mL去离子水,于0℃冰水混合物中解冻2 h,充分震荡3 h,静置12 h,采用雷磁DJS-1D电导仪测得电导率(R1),再将试管置于沸水浴中40 min,震荡3 h,静置12 h,测得电导率(R2),相对电导率=R1/R2×100%。1.3.2 植物学观察 参照《油菜种质资源描述规范和数据标准》[30]观察记载冬前油菜植株的心叶色、幼茎色、叶柄色及生长习性,统计植株叶片数。
1.3.3 干物质测定 将挖取的植株用蒸馏水清洗干净,用滤纸吸干表面水分,分别称量地上部与地下部的鲜重,再置于烘箱内105℃杀青40 min,85℃烘至恒重,用电子天平(精确度为0.01 g)称重记录数据[31]。
1.3.4 生理指标测定 采用电导仪测定相对电导率;采用氮蓝四唑法(nitroblue tetrazolium,NBT)测定超氧化物歧化酶(superoxide dismutase,SOD);采用愈创木酚法测定过氧化物酶(peroxidase,POD);采用紫外吸收法测定过氧化氢酶(catalase,CAT);采用考马斯亮蓝法测定可溶性蛋白质(soluble protein,SP)含量;采用蒽酮比色法测量可溶性糖(soluble sugar,SS)含量;采用磺基水杨酸浸提-酸性茚三酮显色法测量游离脯氨酸(proline,Pro)含量[31]。
1.4 数据处理及评价方法
1.4.1 Logistic回归方程分析 将相对电导率进行Logistic回归分析,回归模型为:$y=\frac{k}{1+a{{e}^{bt}}}$
其中y为相对电导率,t代表处理温度,k为细胞伤害率的饱和容量,a、b为方程参数。为了确定a、b的值,将方程进行线性化处理,
$\text{ln}\left( \frac{k-y}{y} \right)=\text{ln}a\text{-}bt$,令$y'=\text{ln}\left( \frac{k-y}{y} \right)$
则转化为细胞伤害率(y′)与处理温度(t)的直线方程。通过直线回归的方法,求得a、b值及相关系数R,半致死温度LT50=$\frac{ln a}{b}$ [32]。
1.4.2 综合评价数据分析 单项指标耐寒系数计算为K=(低温测定值/常温测定值)。采用聚类分析和标准差系数赋予权重法进行综合评价,运用SPSS19.0进行聚类分析和方差分析,标准差系数赋予权重法计算方法如下:
(1)数据标准化 运用隶属函数对各指标进行标准化处理:
式中,Xij表示i品种第j个指标的隶属值;Xmin表示第j个综合指标的最小值;Xmax表示第j个综合指标的最大值,指标与抗寒性成正相关用隶属函数公式(1)计算隶属函数值,指标与抗寒性成负相关用反隶属函数公式(2)计算隶属函数值。
(2)权重确定 采用标准差系数法(S),用公式(3)计算标准差系数Vj,公式(4)归一化后得到各指标的权重系数Wj。
式中,n表示12个品种,m表示SOD、POD、CAT、SP、SS、Pro 6个指标。
(3)综合评价值 用公式(5)计算各株系的综合评价值。
式中,D值为各参试材料的综合评价值。
2 结果
2.1 甘蓝型冬油菜越冬率及叶片半致死温度(LT50)
2.1.1 甘蓝型冬油菜越冬率 通过比较种植在天水和上川的冬油菜越冬率,发现参试材料在天水均能安全越冬,平均越冬率达92.1%—97.8%,差异不显著(P<0.05),种植区域从海拔1 084 m提高到1 650 m,越冬率有小幅度的下降,但仍达到89.5%以上。北移至兰州和上川后,越冬率大幅度下降,兰州为6.0%—22.5%,上川仅有0—5%,地膜覆盖后,兰州越冬率提高到36.0%—78.6%,上川越冬率仍然较低,仅为0—14.4%,且差异显著(P<0.05)。中国西北冬季地面无积雪覆盖,气温低,气候干燥,新油23和天油2288未能越冬,天油14的越冬率仅为1.0%,抗寒性弱。根据天水、兰州和上川的平均越冬率,将12个参试材料进行聚类分析,初步划分为抗寒强(强)、抗寒较强(中)和抗寒弱(弱)3类(表3)。Table 3
表3
表3参试材料名称及越冬情况
Table 3
| 品种(系) Variety (line) | 越冬率Overwintering rate (%) | 抗寒性分类 Cold tolerance classification | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 天水Tianshui | 兰州Lanzhou | 上川Shangchuan | ||||||||
| 2016—2018 | 平均值Mean | 2016—2018 | 平均值 Mean | 2016—2018 | 平均值 Mean | |||||
| 新油23 Xinyou23 | 94.1 | 90.1 | 92.1a | 6.0 | 40.0 | 23.0g | 0.0 | 0.0 | 0.0e | 弱 Weak |
| 16TS 309-4 | 98.4 | 96.2 | 97.3a | 21.0 | 65.0 | 43.0bc | 4.9 | 14.3 | 9.6a | 强 Strong |
| 16TS 306-3 | 98.9 | 96.5 | 97.7a | 15.4 | 57.5 | 36.5d | 1.8 | 13.6 | 7.7bc | 中 Medium |
| 16TS 312-2 | 94.1 | 93.7 | 93.9a | 22.5 | 65.4 | 44.0ab | 3.3 | 13.7 | 8.5ab | 强 Strong |
| 16TS 309-10 | 98.8 | 94.0 | 96.4a | 15.6 | 44.2 | 30.0f | 3.8 | 12.6 | 8.2abc | 中 Medium |
| 16NPZ 269-1 | 97.7 | 92.5 | 95.1a | 19.5 | 54.8 | 37.2d | 2.9 | 11.7 | 7.3bcd | 中 Medium |
| 16NTS 158 | 95.8 | 91.0 | 93.4a | 16.0 | 50.0 | 33.0e | 2.0 | 10.6 | 6.3cd | 中 Medium |
| 15NS 45-4 | 96.6 | 96.4 | 96.5a | 12.0 | 78.6 | 45.3a | 2.1 | 14.4 | 8.2abc | 强 Strong |
| 2016 8(G) | 95.9 | 93.5 | 94.7a | 20.8 | 63.0 | 42.0c | 1.2 | 10.2 | 5.7d | 强 Strong |
| 2016TS(G)10 | 99.3 | 96.3 | 97.8a | 18.5 | 70.0 | 44.3abc | 4.4 | 11.0 | 7.7bc | 强 Strong |
| 天油14 Tianyou14 | 97.0 | 95.0 | 96.0a | 9.2 | 36.0 | 22.6g | 0.0 | 2.0 | 1.0e | 弱 Weak |
| 天油2288 Tianyou2288 | 95.9 | 89.5 | 92.7a | 7.2 | 38.5 | 22.9g | 0.0 | 0.0 | 0.0e | 弱 Weak |
Different letters indicate significant differences at P<0.05. The same as below
新窗口打开|下载CSV
2.1.2 叶片电导率及半致死温度(LT50) 相对电导率可反映低温胁迫下植物细胞膜受伤害程度。叶片相对电导率随着温度降低,呈增加趋势,在LT50之前,电导率缓慢增加,LT50之后,细胞膜受损严重,电解质外渗,电导率迅速升高(表4)。根据叶片相对电导率拟合成Logistic回归方程,拟合度(R2)在0.87— 0.96,LT50介于-13.38—-4.40℃,根据LT50对参试材料抗寒性的强弱排序为2016TS(G)10(-13.38℃)>16TS 309-4(-9.58℃)>16TS 306-3(-7.96℃)>15NS 45-4(-7.28℃)>16TS 309-10(-6.98℃)>2016 8(G)(-6.83℃)>16TS 312-2(-6.55℃)>16NTS 158(-6.11℃)>16NPZ 269-1(-5.71℃)>天油14(-5.59℃)>新油23(-5.39℃)>天油2288(-4.4℃)。
Table 4
表4
表4不同低温下叶片相对电导率及Logistic拟合方程
Table 4
| 品种(系) Variety(line) | 相对电导率 Relative electrolytic leakage (%) | 回归方程 Regression equation | 拟合度 R2 | 半致死 温度 LT50 (℃) | 抗寒性排序 Cold tolerance ranking | ||||
|---|---|---|---|---|---|---|---|---|---|
| 5℃ | 0℃ | -5℃ | -10℃ | -15℃ | |||||
| 新油23 Xinyou23 | 25.30b | 32.54a | 42.40ab | 61.27a | 80.45abc | y=100/(1+1.949e0.1237x) | 0.96 | -5.4 | 11 |
| 16TS 309-4 | 20.07cd | 31.20b | 31.83abc | 46.67a | 68.67c | y=100/(1+2.6e0.0998x) | 0.92 | -9.6 | 2 |
| 16TS 306-3 | 19.87de | 22.03b | 32.22cd | 48.80a | 80.55abc | y=100/(1+2.976e0.1369x) | 0.89 | -8.0 | 3 |
| 16TS 312-2 | 17.26de | 28.57b | 34.20a | 53.07a | 85.92ab | y=100/(1+2.775e0.1558x) | 0.90 | -6.6 | 7 |
| 16TS 309-10 | 15.14bc | 21.75c | 36.16cd | 52.03a | 85.97ab | y=100/(1+3.251e0.1687x) | 0.93 | -7.0 | 5 |
| 16NPZ 269-1 | 15.95e | 25.16c | 34.23bcd | 66.31a | 88.85a | y=100/(1+2.872e0.1849x) | 0.94 | -5.7 | 9 |
| 16NTS 158 | 19.60bc | 26.33b | 39.00abcd | 54.63a | 86.36ab | y=100(1+2.57e0.1546x) | 0.91 | -6.1 | 8 |
| 15NS 45-4 | 20.05e | 35.76bc | 37.73abcd | 57.80a | 84.99ab | y=100/(1+2.496e0.1256x) | 0.94 | -7.3 | 4 |
| 2016 8(G) | 25.92bc | 31.67a | 41.23ab | 52.13a | 75.63bc | y=100/(1+2.04e0.1044x) | 0.93 | -6.8 | 6 |
| 2016TS(G)10 | 20.66bc | 21.21b | 29.37d | 41.37a | 57.63d | y=100/(1+3.134e0.0854x) | 0.93 | -13.4 | 1 |
| 天油14 Tianyou14 | 20.71bc | 29.04a | 32.90abcd | 58.17a | 76.61bc | y=100/(1+2.22e0.1427x) | 0.91 | -5.6 | 10 |
| 天油2288 Tianyou2288 | 22.71a | 34.65a | 40.22ab | 73.63a | 79.17abc | y=100/(1+1.734e0.125x) | 0.91 | -4.4 | 12 |
新窗口打开|下载CSV
2.2 甘蓝型冬油菜冬前植物学形态及干物质积累特征
在上川试验点,参试材料冬前为半直立或匍匐生长,叶色深绿,心叶、幼茎色呈紫色,或心叶叶缘呈紫色,叶柄出现少量紫色。新油23、16NTS 158、天油14和天油2288地上部分生长茂盛,冬前叶片数达到11.3—12.8片,其余品系的叶片数介于9.1—11.0片(表5和图1)。Table 5
表5
表5甘蓝型冬油菜越冬前植物学记载
Table 5
| 品种(系) Variety(line) | 心叶色 Leaf color | 幼茎色 Young stem color | 叶柄色 Petiole color | 生长习性 Growth habit | 叶片数 Leaves amount |
|---|---|---|---|---|---|
| 新油23 Xinyou23 | 黄绿/紫色 Yellow-green/Purple | 黄绿/紫色 Yellow-green/Purple | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 12.0 |
| 16TS 309-4 | 紫色 Purple | 绿色 Green | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 9.8 |
| 16TS 306-3 | 紫色 Purple | 黄绿 Yellow-green | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 9.1 |
| 16TS 312-2 | 紫色 Purple | 紫色 Purple | 黄绿 Yellow-green | 半直立 Semi-erect | 10.7 |
| 16TS 309-10 | 紫色 Purple | 紫色 Purple | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 9.8 |
| 16NPZ 269-1 | 紫色 Purple | 黄绿 Yellow-green | 黄绿 Yellow-green | 半直立/匍匐 Semi-erect/Prostrate | 11.0 |
| 16NTS 158 | 黄绿/紫色 Yellow-green/Purple | 黄绿 Yellow-green | 黄绿 Yellow-green | 半直立/匍匐 Semi-erect/Prostrate | 11.3 |
| 15NS 45-4 | 紫色 Purple | 黄绿 Yellow-green | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 10.0 |
| 2016 8(G) | 紫色 Purple | 黄绿/紫色 Yellow-green/Purple | 黄绿/紫色 Yellow-green/Purple | 半直立/匍匐 Semi-erect/Prostrate | 9.1 |
| 2016TS(G)10 | 紫色 Purple | 黄绿 Yellow-green | 黄绿 Yellow-green | 半直立/匍匐 Semi-erect/Prostrate | 10.8 |
| 天油14 Tianyou14 | 黄绿/紫色 Yellow-green/Purple | 黄绿 Yellow-green | 黄绿 Yellow-green | 半直立 Semi-erect | 12.6 |
| 天油2288 Tianyou2288 | 黄绿/紫色 Yellow-green/Purple | 黄绿 Yellow-green | 黄绿 Yellow-green | 半直立 Semi-erect | 12.8 |
新窗口打开|下载CSV
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1甘蓝型冬油菜越冬前形态特征
1:新油23;2:16TS 309-4;3:16TS 306-3;4:16TS 312-2;5:16TS 309-10;6:16NPZ 269-1;7:16NTS 158;8:15NS 45-4;9:2016 8(G);10:2016TS(G)10;11:天油14;12:天油2288;13:半直立生长;14:半直立-匍匐生长;15:匍匐生长;16:心叶和幼茎色呈黄绿色;17:心叶呈紫色;18:心叶、幼茎、叶柄呈紫色
Fig. 1Morphological characteristics of Brassica napus before overwintering
1: Xinyou23; 2: 16TS 309-4; 3: 16TS 306-3; 4: 16TS 312-2; 5: 16TS 309-10; 6: 16NPZ 269-1; 7: 16NTS 158; 8: 15NS 45-4; 9: 2016 8(G); 10: 2016TS(G)10; 11: Tianyou14; 12: Tianyou2288; 13: Semi-erect; 14: Semi-erect/Prostrate; 15: Prostrate; 16: The heart leaves and young stems were yellow-green; 17: The heart leaves are purple; 18: The heart leaves, young stems and petiole were purple
冬前优先将有机物质分配至地下部分,是发展冬前壮苗的关键[11]。越冬前(10月29日),16TS 312-2、16NTS 158、天油14和天油2288地上部分的干物质积累量在各品系间相对较大(12.24—15.68 g),地下部的干物质量相对较少(2.25—3.67 g),根冠比较小,为0.19—0.25。16TS 309-4和15NS 45-4根冠比最大,为0.95和0.88,2016TS(G)10次之,为0.59,且材料间差异显著(P<0.05)(表6)。
Table 6
表6
表6冬前参试材料间干物质积累比较
Table 6
| 品种(系) Variety (line) | 地上部干重 Above-ground dry matter | 地下部干重 Under-ground dry matter | 干根冠比 Dry root shoot ratio | |||||
|---|---|---|---|---|---|---|---|---|
| 9/26 | 10/29 | 9/26 | 10/29 | 9/26 | 10/29 | |||
| 新油23 Xinyou23 | 4.75a | 10.98abcd | 0.73a | 3.86bcd | 0.16a | 0.36bcd | ||
| 16TS 306-4 | 3.40ab | 5.35d | 0.82a | 5.12abc | 0.30a | 0.95abc | ||
| 16TS 306-3 | 3.29ab | 6.31bcd | 0.87a | 2.56cd | 0.27a | 0.40cd | ||
| 16TS 312-2 | 3.91ab | 14.58ab | 0.79a | 3.67bcd | 0.21a | 0.24bcd | ||
| 16TS 309-10 | 3.26ab | 5.51cd | 0.73a | 2.69cd | 0.23a | 0.49cd | ||
| 16NPZ269-1 | 2.69b | 14.12abc | 0.65a | 6.08ab | 0.24a | 0.46ab | ||
| 16NTS 158 | 4.58ab | 12.24abcd | 0.66a | 2.27d | 0.14a | 0.19d | ||
| 15NS 45-4 | 3.32ab | 8.95abcd | 0.89a | 7.08a | 0.27a | 0.88a | ||
| 2016 8(G) | 3.78ab | 5.69cd | 0.88a | 2.39d | 0.24a | 0.42d | ||
| 2016TS(G)10 | 3.66ab | 7.52abcd | 0.73a | 4.21bcd | 0.20a | 0.59bcd | ||
| 天油14 Tianyou14 | 2.62b | 14.72ab | 0.58a | 3.25cd | 0.22a | 0.23cd | ||
| 天油2288 Tianyou2288 | 3.91ab | 15.65a | 0.78a | 2.25d | 0.28a | 0.25d | ||
新窗口打开|下载CSV
2.3 甘蓝型冬油菜越冬前生理响应特征
2.3.1 叶片中抗氧化酶活性变化 SOD、POD、CAT酶活性随着气温的降低呈不同程度的增加趋势,且品种(系)差异显著(P<0.05)(图2-A—图2-C)。新油23对低温的响应较早,9月26日测定时,SOD酶活性为293.94 U·g-1 FW,与其他材料相比,活性较高,但10月29日测定时仅为356.46 U·g-1 FW,活性最低。2次测定结果显示,新油23和天油2288抗氧化酶活性增幅较小,而16TS 309-4、16TS 306-3、16TS 309-10、15NS 45-4、2016TS(G)10和天油14随气温降低酶活性增加幅度较大,其中,16TS 309-4、15NS 45-4和天油14的POD酶活性较高,分别为110.6、115.3和118.1 U·g-1·min-1。16TS 306-3、15NS 45-4和2016TS(G)10叶片内CAT酶活性相对较高,为15.97—18.73 U·g-1·min-1,较高的抗氧化酶能清除细胞内过多的活性氧,维持细胞正常代谢,表现出较强的抗寒性。新油23和天油2288随着气温的下降,抗氧化酶活性合成较慢,活性氧的积累对细胞造成氧化胁迫,影响细胞正常代谢,表现出较弱的抗寒性。图2
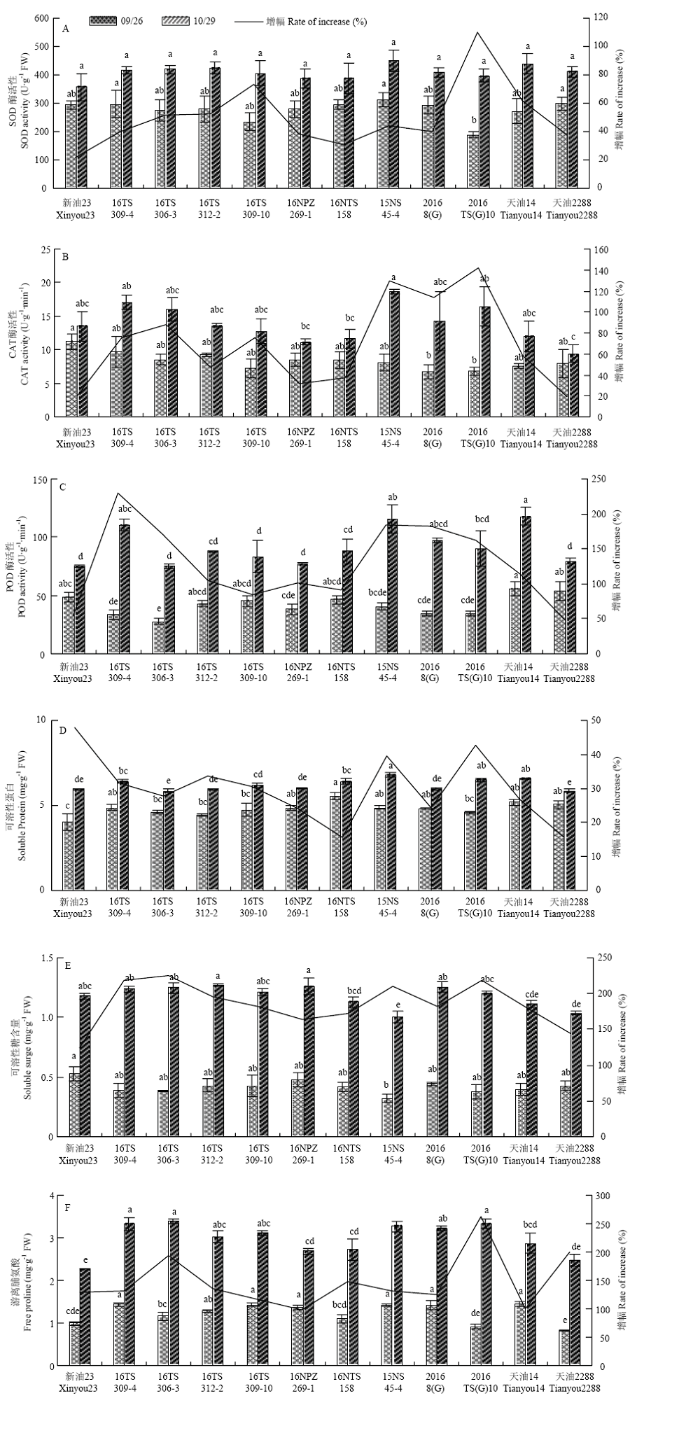 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2甘蓝型冬油菜越冬前生理响应
不同小写字母表示处理在0.05水平差异显著。下同
Fig. 2Physiological Responses of Brassica napus before overwintering
Different letters indicate significant differences at P<0.05. The same as below
2.3.2 叶片渗透调节物质含量的变化 可溶性蛋白和游离脯氨酸含量随气温的降低积累速度缓慢,可溶性糖含量增速相对较快,材料间差异显著(P<0.05)(图2-D—图2-F)。9月26日测定时,16NTS 158、天油14和天油2288有较高的可溶性蛋白质含量,分别为5.54、5.18和5.05 mg·g-1。随着气温降低,10月29日测定时,16TS 309-4、16NTS 158、15NS 45-4、2016TS(G)10和天油14低温胁迫下仍保持较高的可溶性蛋白含量。新油23可溶性糖含量在9月26日测定时为0.524 mg·g-1,与其他材料相比含量最高,随着气温下降增加缓慢,10月29日测定时含量为1.178 mg·g-1。2次测定结果显示,低温胁迫下,16TS 312-2、16NPZ 269-1和2016 8(G)在品种(系)间一直保持相对较高的可溶性糖含量。16TS 309-4、16TS 306-3和2016TS(G)10的游离脯氨酸含量在10月29日测定时为品种(系)间相对较高,分别为3.33、3.39和3.32 mg·g-1。16NTS 158、新油23和天油2288对低温较敏感,可溶性糖和可溶性蛋白的积累早于其他品系,但积累速度缓慢,且天油2288抗氧化酶的含量也较低(图2-A—图2-C),表现出较弱的抗寒性。
2.4 甘蓝型冬油菜春播后的差异比较
2.4.1 春化率差异比较 春化是通过低温处理获得或加速植物开花[27]。春播后,品种(系)间及同一品种(系)内,植株生长发育均表现出巨大差异(图3),主要处于5种不同的生长发育阶段:抽薹后只进行营养生长阶段、现蕾阶段、开花阶段、灌浆及完全成熟阶段。田间统计结果(表7)显示,参试材料间各生长发育阶段的植株比例差异较大。甘蓝型冬油菜在生长发育过程中先抽薹后现蕾,以现蕾作为通过春化阶段的标志性状,比较参试材料间通过春化的植株比例。各品种(系)春化率在4.05%—87.65%,且差异显著(P<0.05)。2016TS(G)10和16TS 309-4春化率最低,分别为4.05%和9.39%,大部分植株处于抽薹后只进行营养生长。新油23、天油14号和天油2288春化率分别为87.65%、55%和85.19%,其中,天油2288成熟植株数占72.22%,比例最高,其次为新油23,成熟植株比例为48.18%。新油23进入现蕾阶段的植株数占39.51%,高于其他材料。其余品系的春化率在17.16%(16TS 306-3)—43.98%(16NPZ 269-1)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3参试材料春播后的田间表现
1:新油23;2:16TS 309-4;3:16TS 306-3;4:16TS 312-2;5:16TS 309-10;6:16NPZ 269-1;7:16NTS 158;8:15NS 45-4;9:2016 8(G);10:2016TS(G)10;11:天油14;12:天油2288
Fig. 3Field performance of materials by spring sowing
1: Xinyou23; 2: 16TS 309-4; 3: 16TS 306-3; 4: 16TS 312-2; 5: 16TS 309-10; 6: 16NPZ 269-1; 7: 16NTS 158; 8: 15NS 45-4; 9: 2016 8(G); 10: 2016TS(G)10; 11: Tianyou14; 12: Tianyou2288
Table 7
表7
表7参试材料春播后不同生长发育阶段的植株个数及所占比例
Table 7
| 品种(系) Variety (line) | 春化率 Vernalization rate (%) | 小区平均株数 Amount | 比例Ratio (%) | 抗寒性分类 Cold tolerance classification | |||
|---|---|---|---|---|---|---|---|
| 未现蕾 Not budding | 现蕾期 Squaring stage | 花期 Flowering stage | 成熟期 Maturing stage | ||||
| 新油23 Xinyou23 | 87.65a | 162 | 12.35 | 39.51 | — | 48.15 | Ⅴ |
| 16TS 309-4 | 9.39g | 181 | 90.61 | 9.39 | — | — | Ⅰ |
| 16TS 306-3 | 17.16f | 204 | 82.84 | 17.16 | — | — | Ⅱ |
| 16TS 312-2 | 27.13e | 387 | 72.87 | 27.13 | — | — | Ⅱ |
| 16TS 309-10 | 25.00e | 216 | 75.00 | 24.54 | — | 0.46 | Ⅱ |
| 16NPZ 269-1 | 43.98c | 241 | 56.43 | 39.00 | 0.41 | 4.15 | Ⅲ |
| 16NTS 158 | 38.71d | 124 | 61.29 | 38.71 | — | — | Ⅲ |
| 15NS 45-4 | 16.40f | 189 | 83.60 | 16.40 | — | — | Ⅱ |
| 2016 8(G) | 24.37e | 119 | 75.63 | 23.53 | 0.84 | — | Ⅱ |
| 2016TS(G)10 | 4.05h | 222 | 95.95 | 4.05 | — | — | Ⅰ |
| 天油14 Tianyou14 | 55.00b | 100 | 45.00 | 31.00 | 7.00 | 17.00 | Ⅳ |
| 天油2288 Tianyou2288 | 85.19a | 108 | 14.81 | 10.19 | 2.78 | 72.22 | Ⅴ |
新窗口打开|下载CSV
根据参试材料间春化率的差异,将参试材料的抗寒性分为5个等级(表7),Ⅰ级(春化率≤10%):16TS 309-4和2016TS(G)10;Ⅱ级(30%≥春化率≥10%):16TS 306-3、16TS 312-2、16TS 309-10、15NS 45-4和2016 8(G);Ⅲ级(50%≥春化率≥30%):16NPZ 269-1和16NTS 158;Ⅳ级(70%≥春化率≥50%):天油14;Ⅴ级(春化率≥70%):新油23和天油2288。
2.4.2 不同生长发育阶段株(薹)高差异比较 由表8可以看出,春化率低的品系,其小区平均薹高亦低,2016TS(G)10的春化率最低为4.05%,小区平均株高为10.77 cm,未现蕾阶段的植株薹高10.50 cm,现蕾阶段的植株薹高17.10 cm,均为品种(系)间最低。新油23和天油2288春化率最高,分别为87.65%和85.19%,小区平均株高80.18和58.21 cm,亦为品种(系)间最高。天油2288成熟阶段的植株平均株高为69.40 cm,低于新油23(122.50 cm)。将春化率、平均株(薹)高、不同生育阶段的平均株(薹)高所占比例进行相关性分析,结果(表9)显示,春化率与平均株(薹)高、成熟期植株所占比例相关系数达0.90—0.96,极显著正相关。
Table 8
表8
表8春播后不同生长发育阶段的株(薹)高
Table 8
| 品种(系) Variety (line) | 平均值Mean | 平均株(薹)高 Average height of plant (bolt) (cm) | ||||
|---|---|---|---|---|---|---|
| 春化率 Vernalization rate (%) | 平均株(薹)高 Average plant (bolt) height (cm) | 未现蕾 Not budding | 现蕾期 Squaring stage | 花期 Flowering stage | 成熟期 Maturing stage | |
| 新油23 Xinyou23 | 87.65 | 80.18 | 28.7 | 44.7 | — | 122.5 |
| 16TS 309-4 | 9.39 | 15.88 | 14.5 | 29.2 | — | — |
| 16TS 306-3 | 17.16 | 18.74 | 17.8 | 23.3 | — | — |
| 16TS 312-2 | 27.13 | 27.46 | 24.8 | 34.6 | — | — |
| 16TS 309-10 | 25.00 | 28.84 | 26.2 | 35.5 | — | 103.0 |
| 16NPZ 269-1 | 43.98 | 37.83 | 34.5 | 35.6 | 85.0 | 99.3 |
| 16NTS 158 | 38.71 | 33.42 | 32.3 | 35.2 | — | — |
| 15NS 45-4 | 16.4 | 20.73 | 16.6 | 41.8 | — | — |
| 2016 8(G) | 24.37 | 20.21 | 17.6 | 28.1 | 34.0 | — |
| 2016TS(G)10 | 4.05 | 10.77 | 10.5 | 17.1 | — | — |
| 天油14 Tianyou14 | 55.00 | 38.79 | 23.1 | 28.7 | 49.4 | 94.4 |
| 天油2288 Tianyou2288 | 85.19 | 58.21 | 22.7 | 33.8 | 46.0 | 69.4 |
新窗口打开|下载CSV
Table 9
表9
表9春化率与平均株(薹)高的相关性分析
Table 9
| 指标 Index | 小区平均株高 Average plant height | 未现蕾 Not budding | 现蕾期 Squaring stage | 成熟期 Maturing stage | |||
|---|---|---|---|---|---|---|---|
| 比例 Proportion | 平均薹高 Average blot height | 比例 Proportion | 平均薹高 Average blot height | 比例 Proportion | 平均株高 Plant height | ||
| 春化率 Vernalization rate | 0.96** | -1.00** | 0.57 | 0.46 | 0.53 | 0.89** | 0.71** |
新窗口打开|下载CSV
2.5 甘蓝型冬油菜抗寒性评价
2.5.1 隶属函数法综合评价 通过计算SOD、POD、CAT、SP、SS和Pro 6个生理指标的抗寒系数(表10),可以看出,参试材料间抗寒系数差异显著(P<0.05)。运用隶属函数法,以抗寒系数为依据,对参试材料的抗寒性进行评价,对每个指标赋予权重,定量比较材料间的抗寒性强弱。用标准差系数赋予权重法计算综合评价(D)值(表11中D值代表参试材料的抗寒性),新油23的D值最小,为0.343,2016TS(G)10的D值最大,为3.106,是新油23的9.06倍,是其他品种(系)的1.05—6.30倍。根据D值对参试材料的抗寒性进行了排序2016TS(G)10>16TS 309-4>16TS 306-3>16TS 309-10>15NS 45-4>2016 8(G)>16TS 312-2>16NTS 158>16NPZ 269-1>天油14>天油2288>新油23,结果与LT50的结果基本吻合。Table 10
表10
表10主要生理指标的抗寒系数评价
Table 10
| 品种(系) Variety(line) | 耐寒系数(K) Anti-cold coefficiency | |||||
|---|---|---|---|---|---|---|
| SOD | CAT | POD | SP | SS | Pro | |
| 新油23 Xinyou23 | 1.267b | 1.267d | 1.563c | 1.478a | 2.870ab | 2.292c |
| 16TS 309-4 | 1.377b | 0.688d | 3.369bc | 1.316abcd | 3.898a | 3.633c |
| 16TS 306-3 | 1.459b | 0.486cd | 2.765abc | 1.278bcd | 3.248ab | 2.959bc |
| 16TS 312-2 | 1.259ab | 0.252bcd | 1.844abc | 1.174cd | 2.889ab | 2.356bc |
| 16TS 309-10 | 1.629ab | 0.407bcd | 2.045abc | 1.336abc | 3.229ab | 2.167bc |
| 16NPZ 269-1 | 1.393ab | 0.232bcd | 2.064abc | 1.243bcd | 2.747ab | 1.995bc |
| 16NTS 158 | 1.291ab | 0.184bcd | 1.950abc | 1.090d | 2.794ab | 2.573bc |
| 15NS 45-4 | 1.242ab | 0.155abc | 2.846abc | 1.395abc | 3.267ab | 2.307bc |
| 2016 8(G) | 1.426ab | 0.158abc | 2.818abc | 1.243bcd | 2.829ab | 2.293ab |
| 2016TS(G)10 | 1.565ab | 0.157abc | 2.641abc | 1.426ab | 3.325ab | 2.318ab |
| 天油14 Tianyou14 | 1.375ab | 0.125ab | 2.157ab | 1.267bcd | 2.688ab | 2.007a |
| 天油2288 Tianyou2288 | 1.198a | 0.100a | 1.542a | 1.100d | 2.338b | 2.990a |
新窗口打开|下载CSV
Table 11
表11
表11隶属函数值和综合评价值结果
Table 11
| 品种(系) Variety (line) | 隶属函数值 Subordinative function | 综合评价值(D) Comprehensive evaluation value | 抗寒性排序 Cold tolerance ranking | |||||
|---|---|---|---|---|---|---|---|---|
| μ(1) | μ(2) | μ(3) | μ(4) | μ(5) | μ(6) | |||
| 新油23 Xinyou23 | 0.000 | 0.436 | 0.000 | 0.125 | 0.643 | 0.000 | 0.343 | 12 |
| 16TS 309-4 | 4.674 | 0.820 | 0.828 | 0.591 | 0.869 | 0.944 | 2.961 | 2 |
| 16TS 306-3 | 3.609 | 0.705 | 0.002 | 0.020 | 0.899 | 1.000 | 2.378 | 3 |
| 16TS 312-2 | 2.804 | 0.352 | 0.194 | 0.355 | 0.777 | 0.740 | 1.839 | 7 |
| 16TS 309-10 | 3.935 | 0.454 | 0.299 | 0.101 | 1.000 | 0.680 | 2.364 | 4 |
| 16NPZ 269-1 | 1.652 | 0.190 | 0.051 | 0.183 | 0.942 | 0.373 | 1.180 | 9 |
| 16NTS 158 | 2.304 | 0.271 | 1.000 | 0.754 | 0.409 | 0.540 | 1.544 | 8 |
| 15NS 45-4 | 3.413 | 1.000 | 0.935 | 1.000 | 0.122 | 0.949 | 2.345 | 5 |
| 2016 8(G) | 3.022 | 0.520 | 0.528 | 0.138 | 0.918 | 0.853 | 2.090 | 6 |
| 2016TS(G)10 | 5.283 | 0.754 | 0.359 | 0.692 | 0.744 | 0.940 | 3.106 | 1 |
| 天油14 Tianyou14 | 1.674 | 0.242 | 0.302 | 0.583 | 0.460 | 0.401 | 1.143 | 10 |
| 天油2288 Tianyou2288 | 1.000 | 0.000 | 0.105 | 0.000 | 0.000 | 0.179 | 0.493 | 11 |
| 权重系数 Weighted value | 0.249 | 0.209 | 0.068 | 0.051 | 0.174 | 0.248 | ||
新窗口打开|下载CSV
2.5.2 春化差异与抗寒性关系 将LT50、越冬率、春化率、抗寒相关的生理指标(SOD、CAT、POD、SP、SS和Pro)进行相关性分析,结果显示,春化率与LT50呈极显著正相关,相关系数为0.741,与越冬率、综合评价(D)值、SOD、CAT、POD和SP极显著负相关,相关系数为-0.96—-0.64(表12),结果表明,春化率可以反映出参试材料抗寒性的强弱,可用于品种(系)的抗寒性评价,其评价结果为2016TS(G)10>16TS 309-4>15NS 45-4>16TS 306-3>2016 8(G)>16TS 309-10>16TS 312-2>16NTS 158>16NPZ 269-1>天油14>新油23>天油2288。得到的结果与LT50、越冬率和综合评价的结果基本吻合。
Table 12
表12
表12春化率与抗寒指标的相关性分析
Table 12
| 指标 Index | 越冬率 Overwintering rate | 半致死温度 LT50 | 综合评价值(D) Comprehensive evaluation value | SOD | CAT | POD | SP |
|---|---|---|---|---|---|---|---|
| 春化率 Vernalization rate | -0.91** | 0.74** | -0.96** | -0.64* | -0.84** | -0.80** | -0.93** |
新窗口打开|下载CSV
3 讨论
3.1 甘蓝型冬油菜抗寒性研究
甘蓝型冬油菜以甘肃天水为原种植区代表,北纬34°33′,海拔从1 084 m上升至1 650 m,越冬率小幅度下降,由94.1%—99.0%下降至89.5%—96.4%。北移至兰州永登后,由于西北地区气候干燥,无积雪覆盖,越冬期漫长,冬前冬后冬油菜生长期短,越冬率大幅下降,甘肃农业大学新育成的16TS 309-4、16TS 306-3、16TS 312-2、16TS 309-10、15NS 45-4、2016 8(G)和2016TS(G)10一系列抗寒材料越冬率显著高于天油14、天油2288和新油23。为了适应低温,植株形态相应发生了变化,越冬前植株半直立或匍匐生长,叶色深绿微带紫红色,根颈大、根冠比高等特点[11, 33-35]。根部有机物的积累,是越冬期植株抵御低温的能量保障,是形成强抗寒能力的物质基础。抗寒性强的品种,冬前优先将有机物分配至地下部,发展壮苗,弱抗寒品种将光合产物运输至地上部,不仅减少了地下部有机物的积累,且易造成地上部的徒长,降低了其抗寒能力[33]。本研究中,越冬前参试材料半直立或匍匐生长,部分品系心叶及幼茎呈紫色,地下部有机物质的积累大于地上部,根冠比增加,与前人研究有较高的一致性。为了适应低温,植物体内引起一系列错综复杂的生理变化,低温胁迫使细胞内活性氧积累,激发抗氧化酶防御系统[36]。可溶性糖、可溶性蛋白、游离脯氨酸含量的积累与抗寒性呈显著的正相关关系[37,38]。闫彪等[39]研究显示,低温诱导使油菜幼苗中的SOD、POD和CAT先升高后降低。杜春芳[9]研究表明,抗寒性强的品系L9叶片中的游离脯氨酸和可溶性糖含量积累明显高于抗寒性弱的品系L7,低温下L9能及时调节可溶性糖含量来抵御低温胁迫,表现出更强的耐寒性和适应性。本研究中,新油23和天油2288对低温较敏感,抗氧化酶系统对低温的响应较早,随着气温下降,植株受害程度加深,抗氧化酶活性和渗透调节物质的积累减少,表现出较弱的抗寒性。16TS 309-4、16TS 306-3、16TS 309-10、15NS 45-4和2016TS(G)10在低温下可溶性蛋白质、可溶性糖及游离脯氨酸含量积累较快,调节细胞渗透势,增加细胞质浓度,降低冰点,预防胞内结冰,表现出较强的抗寒性,与前人的研究结果相符。
春化是一种与低温持续时间成正比的定量反应,当达到植物春化特定要求(即春化饱和),继续置于低温下对加速从营养生长阶段向生殖发育阶段的转变没有影响。WAALEN等[40]研究发现,大田试验条件下,Californium的春化需求低于Banjo,Californium在38.2 d达到了春化饱和,Banjo在52.6 d达到了春化饱和。California和Banjo的耐寒性分别保持了31和67 d。整个越冬期Banjo(LT50 -16.5℃)的平均耐寒性显著高于Californium(LT50 -15.8℃)。2个品种在-18℃处理下的存活率呈显著差异,Banjo平均存活率为16%,显著高于Californium。孙万仓等[41]研究发现,抗寒性强的白菜型冬油菜,其冬性则强,通过春化所需的低温时间越长。本研究利用春季有限的低温时间,诱导植株春化,因参试材料抗寒性不同,春化结果差异巨大,春化率低,未通过春化作用的植株比例高,无法从营养生长进入生殖生长,则抗寒性强。冬性弱的材料通过春化作用的植株比例高,甚至可以完成整个生育期。本研究结果与前人结果有较高的一致性,抗寒性弱的新油23和天油2288在春季有限的低温条件下通过春化作用,进入生殖生长并完成整个生育期,而新选育的16TS 309-4、16TS 306-3、16TS 312-2、16TS 309-10、15NS 45-4、2016 8(G)和2016TS(G)10品系则未能在春季有限的低温时间完成春化作用,则抗寒性强。
3.2 甘蓝型冬油菜新品系的抗寒性评价
低温胁迫下相对电导率是反映植物受冻后细胞膜透性的重要指标,能较直观且准确地反映出植物在低温下受胁迫程度[42,43],外渗电解质越多,电导率越大,细胞膜受害程度越深,则抗寒性越弱,反之抗寒性越强。结合Logistic方程求得半致死温度(LT50),能更加准确地评价植物的抗寒性[14],LT50越低植物的抗寒性越强,反之则越弱。本研究根据LT50将参试材料的抗寒性强弱排序为2016TS(G)10(-13.38℃)>16TS 309-4(-9.58℃)>16TS 306-3(-7.96℃)>15NS 45-4(-7.28℃)>16TS 309-10(-6.98℃)>2016 8(G)(-6.83℃)>16TS 312-2(-6.55℃)>16NTS 158(-6.11℃)>16NPZ 269-1(-5.71℃)>天油14(-5.59℃)>新油23(-5.39℃)>天油2288(-4.4℃)。为了避免因单一指标鉴定结果不一致,而造成最终评价结果的不准确,本研究将SOD、POD、CAT、SP、SS和Pro 6个抗寒指标进行综合评价,根据综合评价(D)值的大小,评价参试材料的抗寒性,得到的结果与LT50基本吻合,该方法已在小麦[44]、胡麻[45]、草坪草[46]等植物在抗逆的综合评价中广泛应用。冬油菜春播后,通过田间植株生长发育的差异,能直观的将参试材料抗寒性进行划分。相关性分析显示,春化率与越冬率、LT50呈极显著正相关,与综合评价(D)值极显著负相关,与SOD、CAT、POD、SP、SS和Pro呈不同程度负相关关系,冬油菜适时春播,田间春化结果的差异可以反应出其抗寒性,可用于评价冬油菜的抗寒性,应用于生产实践中。
3.3 甘蓝型冬油菜新抗寒性品系的意义
随着强抗寒白菜型冬油菜的育成,冬油菜种植区域向北推进5°—13°,使之成为中国北方重要的油料作物,及继冬小麦之后重要的冬季覆盖作物[1, 47-48]。中国西北地区冬寒春旱,研究甘蓝型冬油菜抗寒性,提高甘蓝型冬油菜抗寒性,对优良的种质资源进行品种遗传改良,是中国西北冬油菜产业发展的关键。孙万仓等[49]对白菜型冬油菜抗寒育种研究结果发现,轮回选择代数与越冬率为显著的线性正相关关系,将抗寒性强的品种在寒冷的生态环境下进行轮回选择,是改良北方冬油菜抗寒性的有效途径。本研究中上川试验点地处北纬36°03′,海拔2 150 m,最大冻土深度113 cm、最冷月平均气温-8.1℃、最冷月平均最低气温-13.3℃、极端最低气温-26.5℃、年均气温6.5℃、无霜期142 d、最大降雨量200 mm,为典型的寒旱区,在该生态点能安全越冬的冬油菜品系,利用轮回选择的方法及适当的选择年限,可有效地选育出适应于中国北方种植的强抗寒型甘蓝型冬油菜。21世纪中国气候将持续变暖,尤以北方冬半年最为明显,降水量也呈增加趋势,气候变化将会造成全国种植制度界限不同程度北移[50],种植界限的北移西扩,为甘蓝型冬油菜品种向北向西扩展提供了可能。甘蓝型冬油菜地上部的生长较白菜型冬油菜旺盛,冬、春季可在地表形成植被层。研究表明[51],甘蓝型冬油菜越冬时采摘部分青叶后越冬死苗率为69.1%,全部采光的越冬死苗率在90%以上,因此,叶片不仅对油菜越冬至关重要,还能增加地表覆盖度,保证春播土壤墒情,减少地表裸露面积,切断沙尘来源,有效预防沙尘暴,利于西北农业生产和生态环境改善,使之成为西北生态环境建设的重要组成部分。
4 结论
冬油菜适时春播,可以通过田间春化率、植株的生长发育时期及平均株(薹)高的差异,来鉴定冬油菜抗寒性的强弱。甘肃农业大学新育成的一系列甘蓝型冬油菜在北纬36°03′,海拔2 150 m的生态条件下可以越冬,抗寒性显著优于天油14和天油2288(天水市农业科学研究所选育)及新油23(新疆农业科学院选育)。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.3724/SP.J.1006.2010.02124Magsci [本文引用: 2]

采用多点试验,对冬油菜在原种植区(天水)与北移种植区(张掖等)的主要气候因子、越冬率、生育时期及经济性状进行分析比较。研究结果表明,冬油菜北移后越冬率由原种植区的93.0%~100.0%降为40.0%~95.0%;生育期由280~284 d延至287~289 d,其冬前与冬后生长期缩短,而越冬期延长1倍左右,即140 d左右,整个生长期呈‘短-长-短’的特点,即营养生长有效期缩短及营养生长至生殖生长的过渡期均缩短;株高与分支部位降低, 分支数减少,单株角果数减少, 但千粒重、角粒数等相对增加。冬油菜北移后产量发生了较大变化,抗寒性弱的早熟品种天油8号由原种植区的2 518.8 kg hm<sup>–2</sup>降为1 666.5 kg hm<sup>–2</sup>,减产34.0%,而抗寒性强的晚熟品种陇油6号等由原种植区的741 kg hm<sup>–2</sup>增加到3 333 kg hm<sup>–2</sup>,增幅349.5%。由于生长在相对恶劣的气候生态条件下,越冬期漫长而极端低温低,北移冬油菜栽培品种必须具备优异的抗寒性,同时采用合理播期和密度,保证冬前营养生长期和营养生长量,以确保安全越冬。
DOI:10.3724/SP.J.1006.2010.02124Magsci [本文引用: 2]
采用多点试验,对冬油菜在原种植区(天水)与北移种植区(张掖等)的主要气候因子、越冬率、生育时期及经济性状进行分析比较。研究结果表明,冬油菜北移后越冬率由原种植区的93.0%~100.0%降为40.0%~95.0%;生育期由280~284 d延至287~289 d,其冬前与冬后生长期缩短,而越冬期延长1倍左右,即140 d左右,整个生长期呈‘短-长-短’的特点,即营养生长有效期缩短及营养生长至生殖生长的过渡期均缩短;株高与分支部位降低, 分支数减少,单株角果数减少, 但千粒重、角粒数等相对增加。冬油菜北移后产量发生了较大变化,抗寒性弱的早熟品种天油8号由原种植区的2 518.8 kg hm<sup>–2</sup>降为1 666.5 kg hm<sup>–2</sup>,减产34.0%,而抗寒性强的晚熟品种陇油6号等由原种植区的741 kg hm<sup>–2</sup>增加到3 333 kg hm<sup>–2</sup>,增幅349.5%。由于生长在相对恶劣的气候生态条件下,越冬期漫长而极端低温低,北移冬油菜栽培品种必须具备优异的抗寒性,同时采用合理播期和密度,保证冬前营养生长期和营养生长量,以确保安全越冬。
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2014.03.005Magsci [本文引用: 2]
【目的】低温是影响中国北方冬油菜生长的重要因素之一,冬油菜抗寒机制研究至关重要。从形态、生理、解剖等水平对冬油菜抗寒特性进行研究,探讨各相关指标与冬油菜抗寒性之间的关系,筛选可靠的抗寒鉴定指标,建立冬油菜抗寒性评价数学模型,为中国北方冬油菜抗寒种质筛选及品种抗寒性鉴定提供依据。【方法】对7个不同抗寒性代表性冬油菜品种幼苗进行低温处理,观察其生长习性和叶片解剖特征,测定叶片的生理生化指标、根颈直径及根部干重,采用主成分分析、回归分析、聚类分析和通径分析等方法,对抗寒性进行分析并建立抗寒性评价模型。【结果】研究结果表明,冬油菜低温半致死温度(LT50)与越冬率、5—0℃的POD活性变化、0—-5℃的CAT活性变化、10—5℃的游离脯氨酸变化量(X1)、5—0℃的可溶性蛋白变化量(X2)、生长习性(X5)、根颈直径及地下部干重(X6)呈显著负相关,与0—-5℃的POD变化量、栅海比(X3)、栅栏组织厚度、叶片厚度和气孔面积(X4)呈显著正相关。【结论】主成分分析得到7个品种抗寒性排序与田间越冬率一致;以回归分析筛选出6个指标建立冬油菜抗寒性评价数学模型:Y=-7.58-0.018X1-0.542X2+3.903X3-0.002X4+0.195X5-0.066X6,(R2=1.000);聚类分析将7个品种按抗寒性强弱分为5类,抗寒性强的类型中冬油菜苗期表现匍匐生长,栅海比小,气孔面积小,地下部干重大,Pro活性和SP含量随温度降低增加量大,与之前所做抗寒性排序一致,通径分析表明可溶性蛋白的含量变化是影响冬油菜抗寒性的直接因素。
DOI:10.3864/j.issn.0578-1752.2014.03.005Magsci [本文引用: 2]
【目的】低温是影响中国北方冬油菜生长的重要因素之一,冬油菜抗寒机制研究至关重要。从形态、生理、解剖等水平对冬油菜抗寒特性进行研究,探讨各相关指标与冬油菜抗寒性之间的关系,筛选可靠的抗寒鉴定指标,建立冬油菜抗寒性评价数学模型,为中国北方冬油菜抗寒种质筛选及品种抗寒性鉴定提供依据。【方法】对7个不同抗寒性代表性冬油菜品种幼苗进行低温处理,观察其生长习性和叶片解剖特征,测定叶片的生理生化指标、根颈直径及根部干重,采用主成分分析、回归分析、聚类分析和通径分析等方法,对抗寒性进行分析并建立抗寒性评价模型。【结果】研究结果表明,冬油菜低温半致死温度(LT50)与越冬率、5—0℃的POD活性变化、0—-5℃的CAT活性变化、10—5℃的游离脯氨酸变化量(X1)、5—0℃的可溶性蛋白变化量(X2)、生长习性(X5)、根颈直径及地下部干重(X6)呈显著负相关,与0—-5℃的POD变化量、栅海比(X3)、栅栏组织厚度、叶片厚度和气孔面积(X4)呈显著正相关。【结论】主成分分析得到7个品种抗寒性排序与田间越冬率一致;以回归分析筛选出6个指标建立冬油菜抗寒性评价数学模型:Y=-7.58-0.018X1-0.542X2+3.903X3-0.002X4+0.195X5-0.066X6,(R2=1.000);聚类分析将7个品种按抗寒性强弱分为5类,抗寒性强的类型中冬油菜苗期表现匍匐生长,栅海比小,气孔面积小,地下部干重大,Pro活性和SP含量随温度降低增加量大,与之前所做抗寒性排序一致,通径分析表明可溶性蛋白的含量变化是影响冬油菜抗寒性的直接因素。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2015.01287Magsci [本文引用: 1]
<p class="MsoNormal" align="left"><span lang="EN-US" ><o:p>作物种质资源抗旱性鉴定是获得抗旱资源和挖掘抗旱基因的前提。本研究以21份绿豆种质资源为材料,采用温室内盆栽控水旱胁迫,考察不同品种单株产量、单株荚重、单株粒数、单株地上部生物量、单株生物量、根冠比等12项指标,计算各指标在旱胁迫与对照条件下的比值,运用相关性分析、隶属函数、抗旱系数和抗旱指数方法,评价绿豆成株期抗旱性,筛选抗旱优异种质。结果表明,抗旱系数与单株地上部生物量、单株总生物量、单株荚重、单株粒数、单株有效荚数在旱胁迫与对照条件下的比值呈极显著正相关,与根冠比呈极显著负相关,从而遴选出这7项指标作为绿豆成株期抗旱性鉴定的评价指标;基于隶属函数、抗旱系数和抗旱指数三者呈极显著正相关,且3种评价抗旱分级结果一致性较高,故认为抗旱指数法适宜于大规模绿豆成株期抗旱性鉴定;筛选获得高抗种质3份、抗旱种质6份、中抗种质4份、敏感种质5份和极敏感种质3份。</o:p></span></p>
DOI:10.3724/SP.J.1006.2015.01287Magsci [本文引用: 1]
<p class="MsoNormal" align="left"><span lang="EN-US" ><o:p>作物种质资源抗旱性鉴定是获得抗旱资源和挖掘抗旱基因的前提。本研究以21份绿豆种质资源为材料,采用温室内盆栽控水旱胁迫,考察不同品种单株产量、单株荚重、单株粒数、单株地上部生物量、单株生物量、根冠比等12项指标,计算各指标在旱胁迫与对照条件下的比值,运用相关性分析、隶属函数、抗旱系数和抗旱指数方法,评价绿豆成株期抗旱性,筛选抗旱优异种质。结果表明,抗旱系数与单株地上部生物量、单株总生物量、单株荚重、单株粒数、单株有效荚数在旱胁迫与对照条件下的比值呈极显著正相关,与根冠比呈极显著负相关,从而遴选出这7项指标作为绿豆成株期抗旱性鉴定的评价指标;基于隶属函数、抗旱系数和抗旱指数三者呈极显著正相关,且3种评价抗旱分级结果一致性较高,故认为抗旱指数法适宜于大规模绿豆成株期抗旱性鉴定;筛选获得高抗种质3份、抗旱种质6份、中抗种质4份、敏感种质5份和极敏感种质3份。</o:p></span></p>
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2015.16.002Magsci [本文引用: 1]
【目的】油菜是中国重要的油料作物,主要种植于长江以南区域,该区域铝毒害较为严重,铝毒害已成为限制中国南方油菜产量的重要因素之一。探讨油菜苗期耐铝性评判方法,筛选耐铝基因型,以便减少和防止铝毒对油菜的危害。【方法】以浙油50、中油杂12号、南油杂1号等23个江西省生产上大面积应用的油菜品种为材料,通过盆栽试验,考查铝胁迫处理和对照的株高、根长、根茎粗、地下部干重、地上部干重、叶和根中可溶性糖含量、叶和根中脯氨酸含量、SPAD等性状,以各性状的耐铝系数作为衡量耐铝性的指标,利用主成分分析法、隶属函数法、聚类分析法和逐步回归分析法,对不同基因型油菜进行耐铝性综合评价。【结果】在铝胁迫下,不同基因型油菜形态指标和生理指标对铝胁迫的反应不同,基因型间差异显著。相关性分析表明,各性状的耐铝系数间均存在或大或小的相关性,使它们所提供的信息发生重叠,这些单项指标不能准确评价各油菜品种的耐铝性。利用主成分分析法将10个单项指标综合成为4个相互独立的综合指标,可代表油菜耐铝性86.36%的原始数据信息量。再根据4个综合指标值的贡献率求出其相应的隶属函数值,并依据各综合指标的相对重要性(权重)进行加权,得到不同基因型的耐铝性综合评价值(D值)。通过聚类分析将23个油菜品种划分为3类,浙油50、德油5号、湘杂油2号等3个品种为耐铝类型,中油杂12号、丰油730等13个品种为中度耐铝类型,南油杂1号、创杂油5号等7个品种为不耐铝类型。为了筛选油菜苗期的耐铝性鉴定指标,分析耐铝性鉴定指标与耐铝性之间的关系,建立了耐铝性评价的数学模型,以D值作为因变量、各指标耐铝系数为自变量进行逐步回归分析,得到最优回归方程,23个品种苗期的耐铝性预测值与D值极显著相关。并且筛选出株高、根长、叶中可溶性糖含量、叶中脯氨酸含量等4项对耐铝性有显著影响的指标,可作为油菜苗期耐铝性的鉴定指标。【结论】采用主成分分析、聚类分析和逐步回归分析的方法进行油菜苗期耐铝性的综合评价较为可靠,既避免了单一指标的片面性和不稳定性,又揭示了油菜耐铝相关性状与耐铝性的关系。
DOI:10.3864/j.issn.0578-1752.2015.16.002Magsci [本文引用: 1]
【目的】油菜是中国重要的油料作物,主要种植于长江以南区域,该区域铝毒害较为严重,铝毒害已成为限制中国南方油菜产量的重要因素之一。探讨油菜苗期耐铝性评判方法,筛选耐铝基因型,以便减少和防止铝毒对油菜的危害。【方法】以浙油50、中油杂12号、南油杂1号等23个江西省生产上大面积应用的油菜品种为材料,通过盆栽试验,考查铝胁迫处理和对照的株高、根长、根茎粗、地下部干重、地上部干重、叶和根中可溶性糖含量、叶和根中脯氨酸含量、SPAD等性状,以各性状的耐铝系数作为衡量耐铝性的指标,利用主成分分析法、隶属函数法、聚类分析法和逐步回归分析法,对不同基因型油菜进行耐铝性综合评价。【结果】在铝胁迫下,不同基因型油菜形态指标和生理指标对铝胁迫的反应不同,基因型间差异显著。相关性分析表明,各性状的耐铝系数间均存在或大或小的相关性,使它们所提供的信息发生重叠,这些单项指标不能准确评价各油菜品种的耐铝性。利用主成分分析法将10个单项指标综合成为4个相互独立的综合指标,可代表油菜耐铝性86.36%的原始数据信息量。再根据4个综合指标值的贡献率求出其相应的隶属函数值,并依据各综合指标的相对重要性(权重)进行加权,得到不同基因型的耐铝性综合评价值(D值)。通过聚类分析将23个油菜品种划分为3类,浙油50、德油5号、湘杂油2号等3个品种为耐铝类型,中油杂12号、丰油730等13个品种为中度耐铝类型,南油杂1号、创杂油5号等7个品种为不耐铝类型。为了筛选油菜苗期的耐铝性鉴定指标,分析耐铝性鉴定指标与耐铝性之间的关系,建立了耐铝性评价的数学模型,以D值作为因变量、各指标耐铝系数为自变量进行逐步回归分析,得到最优回归方程,23个品种苗期的耐铝性预测值与D值极显著相关。并且筛选出株高、根长、叶中可溶性糖含量、叶中脯氨酸含量等4项对耐铝性有显著影响的指标,可作为油菜苗期耐铝性的鉴定指标。【结论】采用主成分分析、聚类分析和逐步回归分析的方法进行油菜苗期耐铝性的综合评价较为可靠,既避免了单一指标的片面性和不稳定性,又揭示了油菜耐铝相关性状与耐铝性的关系。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2013.22.005Magsci [本文引用: 2]
【目的】为白菜型冬油菜抗寒种质创新及抗寒品种选育提供依据。【方法】以抗寒性不同的白菜型冬油菜品种为材料,采用田间观察、比色法、冰晶消长法、显微测微、光合测定等技术,探讨北方寒旱区主要冬油菜品种抗寒性差异形成的形态、生理及分子机理。【结果】冬前低温阶段,弱抗寒品种多数气孔关闭或半开放,而强抗寒品种(陇油6号)气孔仍保持完全开放状态,且光合速率、根冠比、根部干物质积累均较高;可溶性蛋白与抗寒性显著正相关,而SOD、POD、CAT活性、游离脯氨酸、MDA含量与抗寒性无明显相关关系;与天油2号相比,低温下陇油7号质外体蛋白提取液形成冰晶体积小、分布均匀,形态以五边形、椭圆形和双菱形为主,具有明显重结晶抑制活性和饰晶活性。【结论】冬前低温阶段,白菜型冬油菜强抗寒品种叶片气孔多数仍保持开放,Pn较高,光合产物优先分配至地下部贮存;根部抗冻蛋白表达上调,增强了其抗寒性。
DOI:10.3864/j.issn.0578-1752.2013.22.005Magsci [本文引用: 2]
【目的】为白菜型冬油菜抗寒种质创新及抗寒品种选育提供依据。【方法】以抗寒性不同的白菜型冬油菜品种为材料,采用田间观察、比色法、冰晶消长法、显微测微、光合测定等技术,探讨北方寒旱区主要冬油菜品种抗寒性差异形成的形态、生理及分子机理。【结果】冬前低温阶段,弱抗寒品种多数气孔关闭或半开放,而强抗寒品种(陇油6号)气孔仍保持完全开放状态,且光合速率、根冠比、根部干物质积累均较高;可溶性蛋白与抗寒性显著正相关,而SOD、POD、CAT活性、游离脯氨酸、MDA含量与抗寒性无明显相关关系;与天油2号相比,低温下陇油7号质外体蛋白提取液形成冰晶体积小、分布均匀,形态以五边形、椭圆形和双菱形为主,具有明显重结晶抑制活性和饰晶活性。【结论】冬前低温阶段,白菜型冬油菜强抗寒品种叶片气孔多数仍保持开放,Pn较高,光合产物优先分配至地下部贮存;根部抗冻蛋白表达上调,增强了其抗寒性。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
为建立葡萄抗寒性的综合评价方法,以7个葡萄品种的枝条为试材,通过人工模拟低温的方法,分别于10、-5、-15、-25、-35、-45℃下处理葡萄枝条,测定枝条可溶性糖、可溶性蛋白、丙二醛、游离脯氨酸、组织含水量、相对电导率等抗寒相关指标,根据抗寒性指标在低温处理过程中的变化,确定葡萄枝条抗寒性指标的有效低温适用范围。并以相对电导率结合Logistic方程计算葡萄枝条的半致死温度(LT<sub>50</sub>),以LT<sub>50</sub>为葡萄抗寒能力的衡量指标,用相关分析法确定评价葡萄抗寒性的5个主要相关指标。用主成分分析法将5个指标转化为综合的主成分,确定指标权重,并结合隶属函数法建立葡萄抗寒的评价方法,运用隶属度函数值求得各葡萄抗寒性的综合评价指数Y。根据综合指数判断抗寒性强弱顺序为: 双优>左优红>北冰红>贝达>5BB>5C>黑比诺。结果表明:葡萄枝条可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量、丙二醛含量、相对电导率等与葡萄抗寒性有极显著或显著相关关系,因此,运用主成分分析法结合隶属度函数法,来综合评价葡萄的抗寒性将更加准确、科学。
Magsci [本文引用: 1]
为建立葡萄抗寒性的综合评价方法,以7个葡萄品种的枝条为试材,通过人工模拟低温的方法,分别于10、-5、-15、-25、-35、-45℃下处理葡萄枝条,测定枝条可溶性糖、可溶性蛋白、丙二醛、游离脯氨酸、组织含水量、相对电导率等抗寒相关指标,根据抗寒性指标在低温处理过程中的变化,确定葡萄枝条抗寒性指标的有效低温适用范围。并以相对电导率结合Logistic方程计算葡萄枝条的半致死温度(LT<sub>50</sub>),以LT<sub>50</sub>为葡萄抗寒能力的衡量指标,用相关分析法确定评价葡萄抗寒性的5个主要相关指标。用主成分分析法将5个指标转化为综合的主成分,确定指标权重,并结合隶属函数法建立葡萄抗寒的评价方法,运用隶属度函数值求得各葡萄抗寒性的综合评价指数Y。根据综合指数判断抗寒性强弱顺序为: 双优>左优红>北冰红>贝达>5BB>5C>黑比诺。结果表明:葡萄枝条可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量、丙二醛含量、相对电导率等与葡萄抗寒性有极显著或显著相关关系,因此,运用主成分分析法结合隶属度函数法,来综合评价葡萄的抗寒性将更加准确、科学。
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2014.01259Magsci [本文引用: 1]
<p>以15份国内胡麻栽培品种为材料, 设自然降雨和正常灌水2个处理, 考察与抗旱性相关的7个农艺性状、8个生理生化指标及产量指标, 采用综合抗旱系数、因子分析、隶属函数、<span>聚类分析和灰色关联度分析相结合的方法</span>, 对其抗旱性进行综合评价、抗旱型划分和评价指标筛选。结果显示, 相关性状<span>指标对干旱胁迫的反应及关联程度各异</span>, 可优先选择与抗旱性关系密切的产量及其相关性状、光合作用因子、叶片抗氧化因子等相关生理生化性状;因子分析表明, 6个公因子可代表胡麻抗旱性90.89%的原始数据信息量。基于抗旱性度量值(drought resistance comprehensive evaluation values, D值)和加权抗旱系数(<span>weight drought resistance coefficient, WDC</span>值)的各品种抗旱性排序相近, 位居前6位的抗旱品种相同。各品种D值与综合抗旱系数(<span>comprehensive drought resistance coefficient, CDC</span>值)、WDC值、产量抗旱系数(yield <span>drought resistance coefficient, Y</span>值)之间均呈极显著正相关, 而各品种Y值与CDC、WDC值间极显著正相关;据D值将供试品种划分为5个抗旱级别, 可较好地反映品种的选育条件及适应地区。试验结果说明以D值为主要参数, 以WDC为辅助评价参数, 评价以产量为主要考量目标的胡麻抗旱性是适宜且准确的;以抗旱性综合评价方法进行胡麻抗旱性综合评价、抗旱型划分、评价指标筛选是准确的。</p>
DOI:10.3724/SP.J.1006.2014.01259Magsci [本文引用: 1]
<p>以15份国内胡麻栽培品种为材料, 设自然降雨和正常灌水2个处理, 考察与抗旱性相关的7个农艺性状、8个生理生化指标及产量指标, 采用综合抗旱系数、因子分析、隶属函数、<span>聚类分析和灰色关联度分析相结合的方法</span>, 对其抗旱性进行综合评价、抗旱型划分和评价指标筛选。结果显示, 相关性状<span>指标对干旱胁迫的反应及关联程度各异</span>, 可优先选择与抗旱性关系密切的产量及其相关性状、光合作用因子、叶片抗氧化因子等相关生理生化性状;因子分析表明, 6个公因子可代表胡麻抗旱性90.89%的原始数据信息量。基于抗旱性度量值(drought resistance comprehensive evaluation values, D值)和加权抗旱系数(<span>weight drought resistance coefficient, WDC</span>值)的各品种抗旱性排序相近, 位居前6位的抗旱品种相同。各品种D值与综合抗旱系数(<span>comprehensive drought resistance coefficient, CDC</span>值)、WDC值、产量抗旱系数(yield <span>drought resistance coefficient, Y</span>值)之间均呈极显著正相关, 而各品种Y值与CDC、WDC值间极显著正相关;据D值将供试品种划分为5个抗旱级别, 可较好地反映品种的选育条件及适应地区。试验结果说明以D值为主要参数, 以WDC为辅助评价参数, 评价以产量为主要考量目标的胡麻抗旱性是适宜且准确的;以抗旱性综合评价方法进行胡麻抗旱性综合评价、抗旱型划分、评价指标筛选是准确的。</p>
DOI:10.11686/cyxb20130528Magsci [本文引用: 1]
<p>对9种暖季型草坪草进行遮阴逆境胁迫研究,测定了叶面积、叶片长度、叶片长宽比、节间距离、地上生物量、叶绿素a含量、叶绿素b含量、叶绿素总量、可溶性糖含量、POD 活性、Fv/Fm、Fv/Fo等12 个形态和生理生化指标,并把各个指标数据转化为遮阴条件下与对照(全光照)的相对值,采用相关分析和主成分分析,将12个指标转化为4个相互独立的综合变量,求得各品种的每一个综合指标值及相应的隶属函数值,经过综合加权进行综合评价,得出9种草坪草的耐阴性强弱排序是:中大1号地毯草、钝叶草、金边钝叶草、兰引Ⅲ号结缕草、海滨雀稗、普通地毯草、近缘地毯草、假俭草、沟叶结缕草。利用逐步回归建立最优回归方程,筛选出叶片长宽比、节间距离、叶绿素a含量、叶绿素b含量和过氧化物酶活性等对草坪草耐阴性有显著影响的5个指标,可作为评价草坪草耐阴性的指标,使评价工作简单化。本研究采用生理形态指标的数理分析对草坪草耐阴性进行综合评价,该方法较为客观全面,可以为草坪草的抗逆性研究提供参考和借鉴。</p>
DOI:10.11686/cyxb20130528Magsci [本文引用: 1]
<p>对9种暖季型草坪草进行遮阴逆境胁迫研究,测定了叶面积、叶片长度、叶片长宽比、节间距离、地上生物量、叶绿素a含量、叶绿素b含量、叶绿素总量、可溶性糖含量、POD 活性、Fv/Fm、Fv/Fo等12 个形态和生理生化指标,并把各个指标数据转化为遮阴条件下与对照(全光照)的相对值,采用相关分析和主成分分析,将12个指标转化为4个相互独立的综合变量,求得各品种的每一个综合指标值及相应的隶属函数值,经过综合加权进行综合评价,得出9种草坪草的耐阴性强弱排序是:中大1号地毯草、钝叶草、金边钝叶草、兰引Ⅲ号结缕草、海滨雀稗、普通地毯草、近缘地毯草、假俭草、沟叶结缕草。利用逐步回归建立最优回归方程,筛选出叶片长宽比、节间距离、叶绿素a含量、叶绿素b含量和过氧化物酶活性等对草坪草耐阴性有显著影响的5个指标,可作为评价草坪草耐阴性的指标,使评价工作简单化。本研究采用生理形态指标的数理分析对草坪草耐阴性进行综合评价,该方法较为客观全面,可以为草坪草的抗逆性研究提供参考和借鉴。</p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7505/j.issn.1007-9084.2015.04.003Magsci [本文引用: 1]
<p>为了解轮回选择对白菜型冬油菜抗寒性及经济性状的影响,以改良北方白菜型冬油菜抗寒性和产量。以4个抗寒性不同的白菜型冬油菜品种在不同生态条件下轮回选择后代为材料,调查越冬率,鉴定农艺性状,分析生态环境与抗寒性之间的关系以及轮回选择的改良效果。结果表明,随轮回选择世代的升高,越冬率明显增加,轮回选择代数与越冬率存在显著的线性正相关,但轮选效果因材料而显著差异,抗寒性强的品种经轮回选择后越冬率的增幅与遗传增益幅度都要比抗寒性弱的品种小。株高、主花序角果数、角粒数、千粒重、单株产量均随轮选世代的增加呈显著增加,而单株角果数降低,抗寒性差的品种遗传增益增幅较大。生态环境对轮回选择效果有较大影响,在气候条件更为严酷的酒泉试点,轮回选择的遗传增益幅度更大。相关性和逐步回归分析结果表明,株高和主花序角果数均与轮回选择群体单株产量呈显著正相关。通径分析结果表明,在C0群体中,各性状对产量均有直接正向效应,但在C1、C2和C3群体中发生了变化,尤其单株角果数,在强抗寒品种陇油6号C1和C2群体中和耐寒品种天油2号C1和C3群体中通经系数均为负值,对单株产量有较大的负向作用。选择更寒冷的生态环境和抗寒性强的基因型进行轮回选择是改良北方冬油菜抗寒性及经济性状的有效途径。</p><p> </p>
DOI:10.7505/j.issn.1007-9084.2015.04.003Magsci [本文引用: 1]
<p>为了解轮回选择对白菜型冬油菜抗寒性及经济性状的影响,以改良北方白菜型冬油菜抗寒性和产量。以4个抗寒性不同的白菜型冬油菜品种在不同生态条件下轮回选择后代为材料,调查越冬率,鉴定农艺性状,分析生态环境与抗寒性之间的关系以及轮回选择的改良效果。结果表明,随轮回选择世代的升高,越冬率明显增加,轮回选择代数与越冬率存在显著的线性正相关,但轮选效果因材料而显著差异,抗寒性强的品种经轮回选择后越冬率的增幅与遗传增益幅度都要比抗寒性弱的品种小。株高、主花序角果数、角粒数、千粒重、单株产量均随轮选世代的增加呈显著增加,而单株角果数降低,抗寒性差的品种遗传增益增幅较大。生态环境对轮回选择效果有较大影响,在气候条件更为严酷的酒泉试点,轮回选择的遗传增益幅度更大。相关性和逐步回归分析结果表明,株高和主花序角果数均与轮回选择群体单株产量呈显著正相关。通径分析结果表明,在C0群体中,各性状对产量均有直接正向效应,但在C1、C2和C3群体中发生了变化,尤其单株角果数,在强抗寒品种陇油6号C1和C2群体中和耐寒品种天油2号C1和C3群体中通经系数均为负值,对单株产量有较大的负向作用。选择更寒冷的生态环境和抗寒性强的基因型进行轮回选择是改良北方冬油菜抗寒性及经济性状的有效途径。</p><p> </p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}