 ,河南省农业科学院经济作物研究所/国家大豆改良中心郑州分中心/农业部黄淮海油料作物重点实验室,郑州 450002
,河南省农业科学院经济作物研究所/国家大豆改良中心郑州分中心/农业部黄淮海油料作物重点实验室,郑州 450002Distribution of Soybean Cyst Nematode Resistance Allele Rhg1 and Rhg4 in Huang-Huai Soybean Varieties
LIAN Yun, LI HaiChao, LI JinYing, WANG JinShe, WEI He, LEI ChenFang, WU YongKang, LU WeiGuo,Institute of Industrial Crops, Henan Academy of Agricultural Sciences/Zhengzhou Subcenter of National Soybean Improvement Center/Key Laboratory of Oil Crops in Huanghuaihai Plains of Ministry of Agriculture, Zhengzhou 450002通讯作者:
收稿日期:2019-01-16接受日期:2019-04-30网络出版日期:2019-08-01
| 基金资助: |
Received:2019-01-16Accepted:2019-04-30Online:2019-08-01
作者简介 About authors
练云,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (382KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
练云, 李海朝, 李金英, 王金社, 魏荷, 雷晨芳, 武永康, 卢为国. 大豆胞囊线虫抗性位点Rhg1和Rhg4优异等位变异在黄淮育成品种中的分布[J]. 中国农业科学, 2019, 52(15): 2559-2566 doi:10.3864/j.issn.0578-1752.2019.15.001
LIAN Yun, LI HaiChao, LI JinYing, WANG JinShe, WEI He, LEI ChenFang, WU YongKang, LU WeiGuo.
0 引言
【研究意义】大豆胞囊线虫(soybean cyst nematode,SCN)病是一种世界性大豆病害,给大豆生产造成严重经济损失[1,2]。种植抗病品种是抵御SCN病最经济有效的措施。但是多年来有限抗源PI88788、Peking、PI437654的过度使用,加之SCN是混居群体,极易产生基因突变和重新组合,已经导致越来越多的SCN个体克服有限抗病品种的抗性,致使部分大豆主产区优势生理小种毒性升级[3,4],甚至有新生理小种出现[5]。因此,有必要明确大豆胞囊线虫优异等位变异在大豆主产区育成品种中的分布,以有效指导抗病品种的合理应用;同时加速抗性资源筛选,挖掘新的抗病基因,以拓宽抗病品种的遗传基础,培育新的抗病品种。【前人研究进展】抗性资源的筛选及合理利用,可以加速抗病品种培育,但传统的SCN抗性鉴定方法,在计数方法上经过多次改进[6],仍然耗时费力,因为鉴定一个品种抗性水平的依据就是寄生在根上的胞囊数目,而胞囊繁殖一代需要25—30 d,周期较长,且繁殖极易受环境温度和湿度影响。因此,目前抗病资源筛选进展缓慢。2012—2015年调查研究发现,在黄淮海地区采集的土样约有70%感染了SCN,且与2001—2003调查研究相比[7],大豆胞囊线虫生理小种组成及分布均有一定的改变,优势生理小种由1号小种演变为2号小种[4];此次调查还发现了新的生理小种X12[5];据报道,美国密苏里州2005年调查研究发现,在采集的土样中,约有50%的样本感染了SCN[8],而在2015—2016年的调查中,88%的样本感染了SCN,且在最广泛使用的2个抗源PI88788和Peking上能够成功寄生的胞囊数目显著增加[9],以上研究结果表明,SCN分布范围有扩大趋势且致病性有增强趋势。大豆胞囊线虫的抗性是受多个位点控制的数量性状且抗性位点之间相互作用,目前已鉴定出rhg1、rhg2、rhg3、rhg4等多个抗性位点[10,11]。其中,rhg1和rhg4是SCN病最重要的抗性位点,rhg1位于G连锁群上,控制着大豆对多个SCN生理小种的抗性,该位点与抗病基因紧密连锁,能够解释50%以上的SCN抗性变异[12,13]。rhg4位于A连锁群上,对3号生理小种和14号生理小种有抗性作用[14,15]。位于rhg4位点的丝氨酸羟甲基转移酶基因和位于rhg1位点的氨基酸转运体α-SNAP和WI12参与植物对SCN产生抗性,但抗性机制不同[12,14,16]。20世纪90年代以来,不同****研究、定位了SCN抗性相关QTL,这些QTL分布在除第2、7、12和13染色体之外的其他16条染色体上[10]。在抗大豆胞囊线虫研究方面,已初步明确抗SCN的主效基因位点rhg1和rhg4的作用机理[14,16-18],发现了微效基因GmSNAP11[19]。大豆基因组测序的完成促进了抗病、耐逆分子标记的鉴定。KADAMA等[20]利用基因组信息发掘了rhg1和Rhg4位点的SNP,并开发成KASP(Kompetitive Allele-Specific PCR)标记。田宇等[21]针对与SCN3抗性显著关联的SNP位点,开发了KASP标记,该位点对抗病材料的选择效率达到92%。史学晖等[22]针对Rhg4位点开发了CAPS和dCAPS标记,其中Rhg4-389鉴定的抗性等位变异对抗病种质的选择效率为97.1%。【本研究切入点】黄淮地区是中国两大大豆主产区之一,SCN发生普遍,但对该地区育成大豆品种中抗SCN的基因型分析尚未见报道,明确该地区育成大豆品种中所含的主要SCN抗性位点,用于有目的地指导大豆生产、减缓由SCN危害带来的经济损失。【拟解决的关键问题】本研究以黄淮地区育成的170份大豆品种为材料,研究大豆胞囊线虫抗性位点Rhg1和Rhg4优异等位变异在黄淮育成品种中的分布,并对含有多个优异等位变异的品种进行表型抗性鉴定,为抗病品种在育种中的合理应用提供依据。1 材料与方法
试验于2018年3月—8月在河南农业科学院经济作物研究所完成。1.1 试验材料
试验材料共189份。源自河南省农业科学院经济作物研究所种质库的黄淮地区育成品种170份(电子附表1),其中,111份来自河南,13份来自北京,19份来自山东,6份来自江苏,9份来自河北,3份来自山西,6份来自安徽,其他3份;Riggs鉴别模式和HG type鉴别寄主、兴县灰皮支(ZDD2315)、邯6192、沧豆6号、中黄57(ZDD24656)、PI567516C、科丰1号、山西小黑豆等已知抗性的优异抗源16份;感病材料Willams82、Essex和Lee68共3份。SCN 2号、4号、5号和X12号小种由河南省农业科学院经济作物研究所SCN生理小种库保存。Table 1
附表1
附表1供检测的黄淮大豆资源名称
Table 1
 |
新窗口打开|下载CSV
1.2 大豆基因组DNA的提取及检测
将参试品种每份播种4—6粒,在蛭石中培养,收集新鲜叶片。采用生工生物工程(上海)股份有限公司的植物基因组DNA快速抽提试剂盒,从大豆叶片中提取基因组DNA。用1%琼脂糖凝胶电泳,确定DNA的完整性,用NanoDrop 2000超微量紫外分光光度计检测DNA浓度,-20℃条件下保存DNA。1.3 标记的选择与反应程序
参照KADAMA等[20]报道的KASP标记,选择其开发的Rhg1位点的Rhg1-2(特异性等位位点1:5′-TCTAATGCATTGGTTATAGCAACAACG-3′;特异性等位位点2:5′-TCTAATGCATTGGTTATAGCAAC AACC-3′;通用引物:5′-TGCTGGCATCTGCCAACTC TGTAAA-3′)和Rhg1-5(特异性等位位点1:5′-GAA AGCCAAAGAACTTGAGGAGC-3′;特异性等位位点2:5′-GAAAGCCAAAGAACTTGAGGAGG-3′;通用引物:5′-CCAACCACCAGGAATATTAAAGGTACA AT-3′);Rhg4位点的Rhg4-3(特异性等位位点1:5′-TCGTTGTGTGATTGTTTTGCAGGGA-3′;特异性等位位点2:5′-TCGTTGTGTGATTGTTTTGCAGGGT -3′;通用引物:5′-CAGAGATCACAGAGTTTCTCCA CCTT-3′)和Rhg4-5(特异性等位位点1:5′-GAGGTG GCCGCCGGAGG-3′;特异性等位位点2:5′-GAGGTG GCCGCCGGAGC-3′;通用引物:5′-CGACCGCATC ATGGGGCTAGAT-3′)共4个标记。PCR反应体系按照KBiosciences(Herts,UK)(http://www.ksre.ksu.edu/ igenomics)操作说明进行。反应程序为94℃ 15 min;94℃ 20 s,65℃—67℃ 60 s,10个循环,每循环降低0.8℃;94℃ 20 s,57℃ 60 s,23个循环。使用Roche LightCycler(LC)荧光阅读仪读取荧光信号,进行样品SNP分型。1.4 SCN抗性鉴定
不同生理小种的繁殖、接种方法及胞囊计数方法参考练云等[4]报道。简述之,将Lee68播种在病土中并在适当的环境下繁殖,1株/杯,5个重复,约25 d后,采用淘洗-过筛法从根系中分离胞囊,并将胞囊接种至刚萌发出新根的待鉴定材料中,每杯按不同处理接种虫卵(2 000虫卵或6 000虫卵),待胞囊处于显囊盛期时,将植株拔出,利用河南省农业科学院大豆课题组自主研发的PDS软件[9],计数根部胞囊数目。2 结果
2.1 Rhg1和Rhg4位点的优异等位变异在已知抗性资源中的频率分布
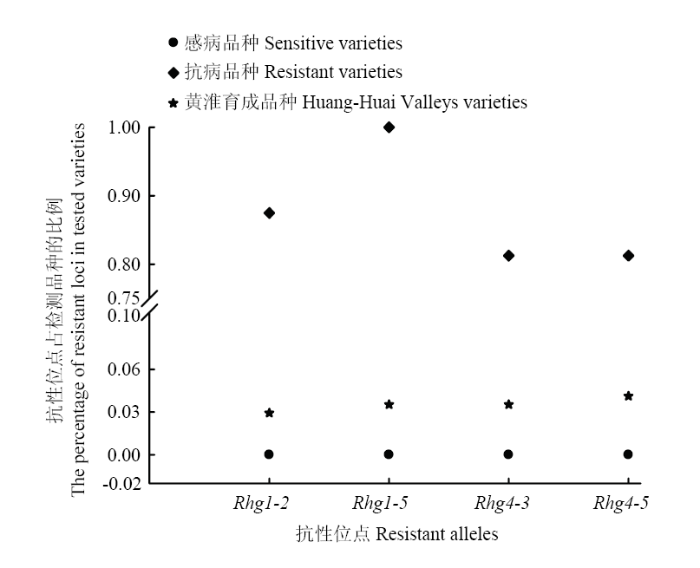
依据抗SCN的Rhg1和Rhg4位点开发的4个KASP标记,选用16份已知的抗性资源和3份感病材料进行基因分型,以验证开发位点的有效性。在抗病材料中,优异等位变异在Rhg1-2(CC)、Rhg1-5(CC)的分布比例分别为87.5%和100%;在Rhg4-3(TT)、Rhg4-5(CC)的分布比例均是81.25%;在感病材料中,优异等位变异在这4个位点中的分布比例均是0(图1);且2种纯合基因型呈明显的簇状分离,此结果验证了所选择4个KASP的有效性,这些位点为功能性优异等位变异,可以用作优异等位变异在资源中分布检测的依据。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1抗SCN优异等位变异在待检测品种中的分布
Fig. 1The distribution of SCN resistance loci in tested varieties (%)
2.2 Rhg1和Rhg4位点的优异等位变异在黄淮育成品种中的频率分析
鉴于所选用KASP的有效性,选取了黄淮地区170份育成品种,系统检测了抗SCN的优异等位变异在黄淮育成品种中的频率分布。结果表明,功能标记Rhg1-2、Rhg1-5、Rhg4-3、Rhg4-5的优异等位变异在黄淮育成品种中的分布频率分别为2.94%(5份)、3.53%(6份)、3.53%(6份)和4.12%(7份)(图1),鉴定出同时携带2个或2个以上优异等位变异的种质6份,占受检测品种的3.53%。说明在黄淮地区育成的大豆品种中,引入SCN优异等位变异的品种偏少。2.3 含优异等位变异种质的抗性评价
根据基因分型结果,选择携带优异等位变异的开豆4号、商豆1201、鲁0305-2、漯4903、潍豆12和潍豆91861,其中,漯4903含Rhg4的2个优异等位变异,另外5个品种含Rhg1和Rhg4 2个位点的4个优异等位变异。利用黄淮地区常见的2号、4号、5号和新发现的具有较强侵染力的X12号小种,进行SCN抗性鉴定。在接种2 000个虫卵/杯的浓度结合接种不同的生理小种,Lee68根系上的平均胞囊数目均大于100个。这6个品种对2号小种均表现高感,对4号小种有1个表现感病、5个表现高感,对5号小种有2个表现感病,1个表现抗病(商豆1201,FI=23.52),2个表现高抗(鲁0305-2,FI=8.98和潍豆91861,FI= 6.28),对X12有2个表现感病,4个表现高感(表1)。综合以上结果,借助KASP标记分型辅助分析,在参与鉴定的6份材料中,快速筛选到了3份抗5号小种的资源。其中,商豆1201表现抗病,鲁0305-2和潍豆91861表现高抗。Table 1
表1
表1携带优异等位变异的种质对不同SCN生理小种的抗性鉴定(接种2 000虫卵/杯)
Table 1
| 品种 Varieties | 2号小种 Race 2 | 4号小种 Race 4 | 5号小种 Race 5 | X12号小种 Race X 12 | ||||
|---|---|---|---|---|---|---|---|---|
| 胞囊数目 No. of Cysts | 胞囊指数 FI | 胞囊数目 No. of Cysts | 胞囊指数 FI | 胞囊数目 No. of Cysts | 胞囊指数 FI | 胞囊数目 No. of Cysts | 胞囊指数 FI | |
| Lee68 | 179.40±9.57 | — | 195.40±6.36 | — | 111.40±8.08 | — | 380.00±18.99 | — |
| 开豆4号 Kaidou 4 | 299.67±23.19 | 167.04 | 234.00±14.06 | 119.75 | 35.00±2.48 | 31.42 | 263.80±6.75 | 69.42 |
| 商豆1201 Shangdou 1201 | 264.25±48.70 | 147.30 | 210.40±16.65 | 107.68 | 26.20±0.91 | 23.52 | 231.50±21.00 | 60.92 |
| 鲁0305-2 Lu 0305-2 | 311.40±38.23 | 173.58 | 185.80±7.88 | 95.09 | 10.00±0.48 | 8.98 | 186.75±20.03 | 49.14 |
| 漯4903 Luo 4903 | 223.60±19.52 | 124.64 | 237.20±17.80 | 121.39 | 64.80±3.91 | 58.17 | 231.80±20.74 | 61.00 |
| 潍豆12 Weidou 12 | 134.80±14.95 | 75.14 | 228.40±12.90 | 116.89 | 58.20±2.19 | 52.24 | 321.80±13.89 | 84.68 |
| 潍豆91861 Weidou 91861 | 192.00±12.96 | 107.02 | 108.60±7.72 | 55.58 | 7.00±0.63 | 6.28 | 206.80±18.57 | 54.42 |
新窗口打开|下载CSV
通过提高接种浓度进一步进行抗性鉴定,在2次接种虫卵浓度均提升至6 000虫卵/杯的情况下,潍豆91861对5号小种均表现为中抗(FI=22.29和FI=18.09),商豆1201(FI=59.13)和鲁0305-2(FI=37.04)表现为感病(表2)。这三个品种的抗性受接种浓度影响,当接种浓度由2 000虫卵/杯提升到6 000虫卵/杯时,商豆1201由抗病表型转变为感病表型;鲁0305-2由高抗表型转变为感病表型;潍豆91861由高抗表型转变为中抗表型。
Table 2
表2
表2接种高浓度SCN虫卵的抗性鉴定(接种6 000虫卵/杯)
Table 2
| 重复 | 品种Varieties | 胞囊数目No. of Cysts | 胞囊指数FI |
|---|---|---|---|
| Lee68 | 487.25±28.59 | — | |
| I | 商豆1201 Shangdou 1201 | 288.13±11.18 | 59.13 |
| 鲁0305-2 Lu0305-2 | 180.50±11.88 | 37.04 | |
| 潍豆91861 Weidou 91861 | 64.38±2.82 | 22.29 | |
| II | Lee68 | 288.86±11.78 | — |
| 潍豆91861 Weidou 91861 | 52.25±1.89 | 18.09 |
新窗口打开|下载CSV
试验结果表明,用2号小种接种中黄57,当接种浓度为2 000虫卵/杯或6 000虫卵/杯,抗性表型均表现高抗;用4号小种接种中黄57,当接种浓度为2 000虫卵/杯时,抗性表型表现为抗病,当接种浓度提升至6 000虫卵/杯时,抗性表型表现为高感(表3)。
Table 3
表3
表3中黄57接种不同浓度虫卵和不同生理小种的抗性鉴定
Table 3
| 生理小种 Race | 接种量(虫卵/杯) Inoculation density (Eggs/cup) | Lee68 | 中黄57 Zhonghuang57 | 胞囊指数 FI |
|---|---|---|---|---|
| 2号小种 Race 2 | 2000 | 108.00±59.87 | 20.20±0.80 | 9.44 |
| 6000 | 194.20±41.90 | 2.40±0.51 | 1.24 | |
| 6000 | 281.60±69.90 | 20.60±2.40 | 7.32 | |
| 4号小种 Race 4 | 2000 | 182.60±36.40 | 22.20±3.31 | 12.16 |
| 6000 | 242.14±44.82 | 331.80±45.45 | 137.03 | |
| 6000 | 278.13±30.62 | 250.57±55.28 | 90.09 |
新窗口打开|下载CSV
3 讨论
大豆的抗病性是与大豆产量、品质相关的重要性状,加速抗性资源筛选可为加速抗病品种培育提供材料支撑。目前,对大豆种质进行传统的SCN抗性鉴定,主要依靠田间表型鉴定或室内表型鉴定,但该鉴定易受环境影响、且费时费力,鉴定的准确性和时效性也较难保证。与传统抗性鉴定方法相比,分子标记技术具有周期短、可进行高通量分析、不受环境条件影响等优点,该技术已经在大豆、水稻、小麦、玉米、马铃薯等[23,24,25,26,27]抗性资源筛选及品种鉴定方面得到了一定范围的应用。目前,已有关于利用SSR标记鉴定Rgh1的报道,比如Satt309标记已被应用于育种程序[28,29]。然而,Satt309标记仅能检测到含3个拷贝的Rgh1。在已有的分子标记技术中,SNP较其他遗传标记相比,SNP技术分析系统具有自动化程度高、通量大、速度快、适合大规模操作的特点,已有利用SNP标记鉴定Rhg1和Rhg4的报道[30,31,32]。而近几年发展起来的新一代KASP技术,具有高通量、低错误率和成本低廉的特点,为加速优异种质资源筛选,提高种质利用效率创造了条件。本研究用到的4个KASP标记是利用SoySNP50K iSelect BeadChip对19 652份大豆种质(来自USDA种质库)进行基因分型[33],结合对大豆胞囊线虫Rhg1和Rhg4位点进行遗传多样性评价而开发的多态性SNP分子标记。利用这些标记在95份大豆种质和3个重组自交系群体进行基因型鉴定,结果表明,来自Rhg1位点的2个标记(Rhg1-2和Rhg1-5)可以区分多拷贝型(PI88788-type)、低拷贝型(Peking-type)和单拷贝型(Williams 82)3种拷贝数的变异;来自Rhg4位点的标记能够检测到Peking-type 的抗病基因型[20]。在大豆更多研究方面:2018年,叶俊华等[19]利用KASP技术对中国引进的1 489份大豆种质的多个性状(包括大豆胞囊线虫病、大豆花叶病毒病、耐盐性等)进行了基因型鉴定,发现携带2个或2个以上优异等位变异的种质有36份,标记的有效性在所用已知抗源中得到了验证,但该研究未对新发现的抗源做进一步的抗性鉴定;田宇等[21]利用抗大豆胞囊线虫SCN3-11位点开发的KASP标记GmSNAP11-5149,鉴定了来自8个国家的202份大豆资源;PATIL等[23]建立了基于耐盐基因GmCHX1的高通量SNP marker,并开发了与该基因结构变异相关的6个KASP标记,对耐盐性状的表型鉴定效率达91%。
目前未见对田间虫卵浓度进行调查的报道,但在实际生产中,田间虫卵密度达不到室内试验接种浓度,理由:一是在前期的取样调查中,约有2/3的地块感染了SCN病,但是没有繁殖到足够用于对病土进行生理小种鉴定的胞囊;二是依据经验,即便是接种2 000虫卵/杯,也已超过田间实际的虫卵浓度。所以,推测本次用于鉴定的商豆1201和鲁0305-2是抗病品种。前期研究数据也表明,有些品种的抗性不受接种浓度的影响,比如中黄57对2号小种:在接种浓度2 000虫卵/杯或6 000虫卵/杯情况下,胞囊指数均小于10,表现高抗(表3),兴县灰皮支(ZDD2315)也稳定高抗1—6号小种[5];还有一些品种的抗性表型受接种浓度的影响,比如中黄57对4号小种:在接种浓度为2 000虫卵/杯情况下,胞囊指数为12.16,表现抗病,但是当接种浓度提升至6 000虫卵/杯情况下,胞囊指数大于60,对4号小种表现为高感(表3)。目前抗性鉴定标准一般是依据对照品种Lee68上的胞囊数目≥30个/株,即认为本次试验结果有效,但在实际应用中,部分种质的抗性水平确实受接种浓度影响[34],因此,抗性鉴定依据有待进一步探讨。考虑到田间感染SCN地块的实际虫卵浓度达不到2 000虫卵/杯这样的接种浓度,因此建议接种2 000—2 500虫卵/杯,且Lee68上的胞囊数目≥30个/株,暂可作为鉴定抗性依据。根据这个标记,本试验所用材料已验证表型的共有25份,其中22份标记基因型跟表型结果一致,正确率达到88.0%。
从对黄淮地区育成品种的鉴定结果看,黄海地区育成品种中,含有大豆胞囊线虫优异等位变异的品种偏少,常年种植感病品种导致不仅影响大豆产量,而且SCN群体内异交机率增加,这可能是导致优势生理小种发生变化及新生理小种出现的一个潜在原因。鉴于SCN生理小种毒性在黄淮地区有增加趋势,5号小种是仅次于2号小种的优势生理小种,建议在抗SCN育种中,利用筛选出来的抗源,通过基因聚合培育兼抗2号和5号小种的大豆品种。
4 结论
黄淮育成品种中含Rhg1或Rhg4优异等位变异的品种偏少,应加大抗病品种培育力度。利用文中的KASP标记可以进行大豆胞囊线虫抗性资源的快速筛选,提升抗性资源筛选效率。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2016.01479URLMagsci [本文引用: 3]

<p>大豆胞囊线虫(SCN)在黄淮地区普遍发生,调查小种分布情况,确定优势小种对抗病育种有重要意义。2012—2015年,取样调查黄淮地区6个省份土样,利用Riggs模式鉴定生理小种,绘制黄淮地区SCN生理小种分布图,并与文献报道结果对比,探讨黄淮地区SCN生理小种类型及其分布规律。结果表明,该病害在黄淮大豆主产区均有分布,在采集受SCN感染的322份土样中,112份被鉴定出生理小种类型,包括1号、2号、3号、4号、5号、6号和11号小种。其中,57份为2号小种,占样本总体的50.9%;26份土样为5号小种,占23.2%;11份土样为4号小种,占9.8%,1号、3号、6号和11号小种分别占总体的4.5%、5.4%、4.5%和1.8%。依据不同生理小种在各省发生频率由高到低的顺序,河南分布5号、2号、3号、11号小种;河北分布2号、5号、6号、3号、4号小种;安徽分布2号、5号、6号、3号小种;山西分布2号、4号、5号、1号、3号、11号小种;山东分布2号、3号、5号、1号、6号小种;江苏分布2号、5号、1号小种。以上结果表明,2号小种是目前黄淮海地区的优势小种,其次是5号小种,致病力最强的4号小种主要分布在山西省。在黄淮海地区,抗线虫育种目标应以抗2号生理小种为主,兼抗5号小种,部分地区应以兼抗2号和4号小种为主。在黄淮地区3号、6号和11号小种是新发现的小种。与2001-2003年调查结果比较,黄淮海地区大豆胞囊线虫生理小种组成及分布有一定的改变。</p>
DOI:10.3724/SP.J.1006.2016.01479URLMagsci [本文引用: 3]
<p>大豆胞囊线虫(SCN)在黄淮地区普遍发生,调查小种分布情况,确定优势小种对抗病育种有重要意义。2012—2015年,取样调查黄淮地区6个省份土样,利用Riggs模式鉴定生理小种,绘制黄淮地区SCN生理小种分布图,并与文献报道结果对比,探讨黄淮地区SCN生理小种类型及其分布规律。结果表明,该病害在黄淮大豆主产区均有分布,在采集受SCN感染的322份土样中,112份被鉴定出生理小种类型,包括1号、2号、3号、4号、5号、6号和11号小种。其中,57份为2号小种,占样本总体的50.9%;26份土样为5号小种,占23.2%;11份土样为4号小种,占9.8%,1号、3号、6号和11号小种分别占总体的4.5%、5.4%、4.5%和1.8%。依据不同生理小种在各省发生频率由高到低的顺序,河南分布5号、2号、3号、11号小种;河北分布2号、5号、6号、3号、4号小种;安徽分布2号、5号、6号、3号小种;山西分布2号、4号、5号、1号、3号、11号小种;山东分布2号、3号、5号、1号、6号小种;江苏分布2号、5号、1号小种。以上结果表明,2号小种是目前黄淮海地区的优势小种,其次是5号小种,致病力最强的4号小种主要分布在山西省。在黄淮海地区,抗线虫育种目标应以抗2号生理小种为主,兼抗5号小种,部分地区应以兼抗2号和4号小种为主。在黄淮地区3号、6号和11号小种是新发现的小种。与2001-2003年调查结果比较,黄淮海地区大豆胞囊线虫生理小种组成及分布有一定的改变。</p>
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
【目的】探讨黄淮地区大豆胞囊线虫(Heterodera glycines Ichinohe)的种类及其分布。【方法】2001~2003年在黄淮大豆产区依据Riggs的鉴别模式对38个地点大豆胞囊线虫(SCN)生理小种作抽样调查,并结合文献资料,绘制出黄淮地区大豆胞囊线虫生理小种分布图。【结果】大豆胞囊线虫主要分布在山东、河北、北京、山西大部分地区、河南东部与北部、安徽北部;在山西南部及河南西南部地区的抽样调查中未检测到大豆胞囊线虫。其中1号生理小种主要分布在山东济南以南及以东地区,河南北部与河北南部交界地区,河南漯河、周口及安徽阜阳地区。4号生理小种主要分布在河南、山东、安徽交界地区,山西、北京地区,以及山东黄河三角洲地区。2号生理小种主要分布在山东聊城、德州地区,河北石家庄地区,河南焦作、获嘉地区。7号生理小种主要分布在山东半岛和河南开封、滑县、温县等地。5号生理小种在河南和河北有零星分布。另外,在河南商丘地区新发现有9号生理小种。【结论】黄淮地区的优势小种是1号和4号生理小种,抗线虫育种应该以兼抗1号和4号生理小种为主要目标。各生理小种的分布没有明显分界,优势小种分布区域中存在其他生理小种。在过去的10年中,该地区生理小种的组成相对稳定,本研究结果可供大豆抗线虫育种参考。
URLMagsci [本文引用: 1]
【目的】探讨黄淮地区大豆胞囊线虫(Heterodera glycines Ichinohe)的种类及其分布。【方法】2001~2003年在黄淮大豆产区依据Riggs的鉴别模式对38个地点大豆胞囊线虫(SCN)生理小种作抽样调查,并结合文献资料,绘制出黄淮地区大豆胞囊线虫生理小种分布图。【结果】大豆胞囊线虫主要分布在山东、河北、北京、山西大部分地区、河南东部与北部、安徽北部;在山西南部及河南西南部地区的抽样调查中未检测到大豆胞囊线虫。其中1号生理小种主要分布在山东济南以南及以东地区,河南北部与河北南部交界地区,河南漯河、周口及安徽阜阳地区。4号生理小种主要分布在河南、山东、安徽交界地区,山西、北京地区,以及山东黄河三角洲地区。2号生理小种主要分布在山东聊城、德州地区,河北石家庄地区,河南焦作、获嘉地区。7号生理小种主要分布在山东半岛和河南开封、滑县、温县等地。5号生理小种在河南和河北有零星分布。另外,在河南商丘地区新发现有9号生理小种。【结论】黄淮地区的优势小种是1号和4号生理小种,抗线虫育种应该以兼抗1号和4号生理小种为主要目标。各生理小种的分布没有明显分界,优势小种分布区域中存在其他生理小种。在过去的10年中,该地区生理小种的组成相对稳定,本研究结果可供大豆抗线虫育种参考。
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
URL [本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
DOI:10.3724/SP.J.1006.2015.01463URLMagsci [本文引用: 1]
<div ><span new=""><span><span new=""><span><span new=""><span><span >大豆胞囊线虫(soybean cyst nematode,SCN)严重危害世界大豆生产,Rhg4 (resistance to Heterodera glycines 4)是控制大豆SCN抗性的2个主效位点之一。本研究针对Rhg4 (GmSHMT)上的2个单核苷酸多态性(single nucleotide polymorphisms, SNP)位点开发了快速、经济、简便易行的CAPS (Rhg4-389)和dCAPS标记(Rhg4-1165),并用开发的2个标记鉴定了以大豆胞囊线虫应用核心种质为主的193份代表性抗感种质。结果表明,Rhg4-389和Rhg4-1165位点间存在显著连锁不平衡(P=0.0001,r2=0.87),可形成4种单倍型。Rhg4-389-G/Rhg4-1165-T和Rhg4-389-C/Rhg4-1165-A为优势单倍型,稀有单倍型Rhg4-389-G/Rhg4-1165-A和Rhg4-389-C/Rhg4-1165-T是在中国抗源中新发现的单倍型。结合193份种质对SCN 3号小种抗性鉴定分析发现,Rhg4-389-G和Rhg4-1165-T主要存在于抗病种质,它们形成的单倍型对抗病种质鉴定效率可达94.1%。本研究开发了可用于辅助大豆SCN抗性鉴定且方便育种家利用的CAPS/dCAPS标记,且用其摸清了应用核心种质等重要抗源在Rhg4位点的“本底”,为育种家有效利用这些优异抗源提供了重要信息。</span></span></span></span></span></span></span></div>
DOI:10.3724/SP.J.1006.2015.01463URLMagsci [本文引用: 1]
<div ><span new=""><span><span new=""><span><span new=""><span><span >大豆胞囊线虫(soybean cyst nematode,SCN)严重危害世界大豆生产,Rhg4 (resistance to Heterodera glycines 4)是控制大豆SCN抗性的2个主效位点之一。本研究针对Rhg4 (GmSHMT)上的2个单核苷酸多态性(single nucleotide polymorphisms, SNP)位点开发了快速、经济、简便易行的CAPS (Rhg4-389)和dCAPS标记(Rhg4-1165),并用开发的2个标记鉴定了以大豆胞囊线虫应用核心种质为主的193份代表性抗感种质。结果表明,Rhg4-389和Rhg4-1165位点间存在显著连锁不平衡(P=0.0001,r2=0.87),可形成4种单倍型。Rhg4-389-G/Rhg4-1165-T和Rhg4-389-C/Rhg4-1165-A为优势单倍型,稀有单倍型Rhg4-389-G/Rhg4-1165-A和Rhg4-389-C/Rhg4-1165-T是在中国抗源中新发现的单倍型。结合193份种质对SCN 3号小种抗性鉴定分析发现,Rhg4-389-G和Rhg4-1165-T主要存在于抗病种质,它们形成的单倍型对抗病种质鉴定效率可达94.1%。本研究开发了可用于辅助大豆SCN抗性鉴定且方便育种家利用的CAPS/dCAPS标记,且用其摸清了应用核心种质等重要抗源在Rhg4位点的“本底”,为育种家有效利用这些优异抗源提供了重要信息。</span></span></span></span></span></span></span></div>
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}