Disease Resistance of Rice stripe virus NS3-Transgenic Nicotiana benthamiana
WU GenTu, CHEN GuangXiang, ZHANG JiaYuan, HU Qiao, MA MingGe, DOU YanXia, LI MingJun, QING Ling,Chongqing Key Laboratory of Plant Disease Biology, College of Plant Protection, Southwest University, Chongqing 400716
Abstract 【Objective】The protein encoded by NS3 is a suppressor of RNA silencing of Rice stripe virus (RSV). In previous study, it was found that NS3 expression enhanced the resistance of Nicotiana benthamiana to RSV using transgenic technology. In order to reveal the detailed function of NS3 in host plants, the resistance of NS3-transgenic N. benthamiana to other tobacco diseases will be analyzed in this study.【Method】Wild-type and NS3-transgenic N. benthamiana were used to study the infection of Potato virus X (PVX), Ralstonia solanacearum, and Phytophthora parasitica var. nicotianae. PVX infectious clone was inoculated by agro-infiltration, viral symptoms were observed. The total RNA of infected leaves was extracted, and viral relative accumulation was detected by RT-qPCR. The method of filling root was used to inoculate R. solanacearum and P. parasitica var. nicotianae, respectively, disease symptoms were observed, and the incidence and disease index were analyzed.【Result】The main symptoms of PVX in N. benthamiana leaves were mosaic and chlorosis, and the NS3-transgenic N. benthamiana plants had the same PVX symptoms as those of wild-type plants. At 10 days past inoculation (dpi), the results of RT-qPCR showed that NS3 expression inhibited the accumulation of PVX in N. benthamiana, the relative accumulation of PVX in NS3-6 line and NS3-9 line was 68.17% and 81.01% of that of wild-type N. benthamiana, respectively. After the infection of R. solanacearum, the symptoms both in wild-type and NS3-transgenic N. benthamiana plants were wilting and damping-off. In the early stage of disease, the expression of NS3 was beneficial to the infection of R. solanacearum to a certain extent, which showed that it was more sensitivity to bacterial wilt. At 14 dpi, the incidence and disease index of bacterial wilt of wild-type N. benthamiana were 100.00% and 78.67, while the incidence of NS3-6, NS3-9 line was 95.00%, 98.33%, and the disease index was 70.00, 73.00, respectively. After 14 dpi, the incidence and disease index increased gradually, while the disease index of NS3-transgenic lines was lower than that of wild-type plants. The black-brown necrotic patches at the base of stem were observed both in wild-type and transgenic plants after P. parasitica var. nicotianae infection, and the expression of NS3 did not affect the symptoms of N. benthamiana. At the early stage of P. parasitica var. nicotianae infection, the incidence of black shank disease in NS3-6 was higher than that of wild-type N. benthamiana, however, during the whole process of black shank disease, the disease index of NS3-transgenic lines was lower than that of wild-type N. benthamiana. At 12 dpi, the incidence and disease index of black shank disease in wild-type N. benthamiana were 96.67% and 77.50, while the incidence of NS3-6, NS3-9 line was 90.00%, 86.67%, and the disease index was 64.17, 62.08, respectively.【Conclusion】The observation of pathogen infection process showed that the expression of NS3 affected the infection and severity of tobacco diseases on N. benthamiana to a certain extent, and the effects on different pathogens were also different. Keywords:Rice stripe virus (RSV);NS3;transgenic Nicotiana benthamiana;disease resistance
PDF (2613KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 吴根土, 陈广香, 张珈源, 胡桥, 马明鸽, 窦彦霞, 李明骏, 青玲. 转水稻条纹病毒NS3本氏烟的抗病性[J]. 中国农业科学, 2019, 52(10): 1710-1720 doi:10.3864/j.issn.0578-1752.2019.10.005 WU GenTu, CHEN GuangXiang, ZHANG JiaYuan, HU Qiao, MA MingGe, DOU YanXia, LI MingJun, QING Ling. Disease Resistance of Rice stripe virus NS3-Transgenic Nicotiana benthamiana[J]. Scientia Agricultura Sinica, 2019, 52(10): 1710-1720 doi:10.3864/j.issn.0578-1752.2019.10.005
0 引言
【研究意义】植物RNA沉默(RNA silencing)不仅调节内源基因的表达,也是抵御病原物侵染的重要机制[1]。然而,几乎所有的植物病毒均会编码RNA沉默抑制子(viral suppressors of RNA silencing,VSR)来干扰寄主的RNA沉默,进而影响植物的正常生长发育[2,3,4,5]。同时,VSR还可协助病毒积累,加重病毒病症状[6,7,8]。前期研究发现,水稻条纹病毒(Rice stripe virus,RSV)NS3同时具有作为VSR协助RSV侵染和作为寄主防卫反应诱导因子抑制病害发生的双重功能[9,10]。本研究继而以转RSV NS3本氏烟(Nicotiana benthamiana)为材料,分析其对烟草重要病害的抗性,对深入揭示VSR在寄主体内的双重功能具有重要意义,并可为植物抗病育种提供理论依据。【前人研究进展】由RSV引发的水稻条纹叶枯病在我国稻区时有发生,造成严重的经济损失[11]。RSV基因组由4条RNA单链(RNA1、RNA2、RNA3、RNA4)组成,共编码7个蛋白。除RNA1反义编码RNA依赖RNA聚合酶(RNA dependent RNA polymerase,RdRp)外,其他3条RNA链均以双义编码策略编码病毒蛋白,分别含有2个不重叠的开放阅读框(open reading frame,ORF),这2个ORF由具有转录终止功能的非编码区(intergenic region,IR)间隔[12,13]。NS3是RNA3正义链编码的蛋白,被鉴定为RNA沉默抑制子[14]。通过遗传转化方法获得转NS3水稻(Oryza sativa)和本氏烟,表型观察发现NS3表达的转基因植株未呈现生长异常,因此认为其不是症状决定因子[9,14]。但是,NS3蛋白在维持病毒与传播介体昆虫,病毒与寄主植物的互作过程中起到很大的作用。NS3可以与介体灰飞虱(Laodelphax striatellus)体内蛋白酶体途径中的重要因子LsRPN3(L. striatellus regulatory-particle non-ATPase subunit 3)互作,进而抑制26S蛋白酶体途径,帮助昆虫介体传播病毒[15]。NS3与OsDRB1(O. sativa dsRNA-binding protein)互作,增加microRNA(miRNA)的积累,从而有利于RSV在水稻中的侵染和积累[16]。姜良良[17]研究发现,NS3蛋白与本氏烟核定位蛋白NbPip3(N. benthamiana protein interacting with p3)互作,从而促进自噬途径对NS3降解,参与植物抗RSV侵染的作用途径。因此,NS3在RSV传播、侵染及在寄主体内积累等过程中均起到一定的作用,其在病原与寄主互作过程中起着多重作用。随着气候逐渐变暖和植物产品调动日益频繁,植物病原物不断扩散及危害加重的可能性增大,为抗病育种工作者带来挑战。在抗植物病毒研究中,人们通常是以转入病毒基因来获取抗病毒材料[18,19,20]。研究发现,VSR除了作为RNA沉默抑制子协助病毒侵染,还可以促使寄主对其他病原物产生抗性,例如转入烟草蚀纹病毒(Tomato etch virus,TEV)HC-Pro的植物对番茄黑斑病毒(Tomato black ring virus,TBRV)和烟草霜霉菌(Peronospora tabacina)抗性增强[21]。【本研究切入点】在前期工作中,笔者通过农杆菌介导遗传转化获得转NS3水稻和本氏烟。转基因水稻接种RSV后发现,在RSV侵染早期,水稻对RSV敏感性增强,表现为症状加重、病毒积累量增多;而在侵染后期,与野生型植株上的病毒相比,转NS3水稻上病毒症状较轻,而病毒积累量却无明显变化。进一步抗性分析发现,转NS3水稻对稻瘟病菌(Magnaporthe oryzae)的抗性增强[9];在转NS3本氏烟上接种RSV后发现病毒症状较轻,且病毒的发病率和积累量显著降低,通过转录组测序分析发现,NS3表达影响了寄主内源抗性基因表达[10]。因此,NS3表达是否会通过影响本氏烟抗性基因表达而增强对其他病原菌的抗性有待验证。【拟解决的关键问题】针对该问题,本研究在转基因本氏烟上分别接种马铃薯X病毒(Potato virus X,PVX)、烟草青枯病菌(Ralstonia solanacearum)、烟草黑胫病菌(Phytophthora parasitica var. nicotianae),观察发病情况并统计发病率和病情指数,为揭示NS3在寄主体内的作用机制和抗病育种提供理论依据。
1 材料与方法
试验于2017年4月至2018年6月在西南大学植物保护学院植物病理学温室完成。
1.1 试验材料
供试烟草:野生型本氏烟(wild-type N. benthamiana)、转NS3本氏烟(NS3-transgenic N. benthamiana)由笔者实验室保存,种于西南大学植物保护学院植物温室中,温室条件为16 h光照/8 h黑暗,温度24℃。植株培养至4—6叶期备用。在前期工作中,已对转NS3本氏烟的生长发育与野生型植株进行了比较,NS3表达对本氏烟发芽速率、生长速率、株型、叶型及株高均未造成影响,与野生型植株生长发育相一致;同时,利用PCR技术及Northern杂交技术检测了阳性植株和基因转录情况[10],利用Western杂交技术检测了阳性植株NS3蛋白表达[22]。
PCR检测:用抽提的总DNA作为模板,以已构建好的pCV/NS3质粒作为阳性对照,野生型植株DNA作为阴性对照,ddH2O作为空白对照,用RSV NS3基因特异性引物进行PCR扩增,引物序列为NS3 det F:5′- ATGGAAGGGAAGACTATGTTC-3′,NS3 det R:5′-TTAACCCATGCTGGGAGTATC-3′,检测转NS3本氏烟的阳性率。
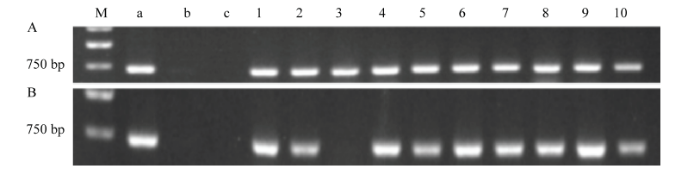
对提取成功的本氏烟植株DNA进行PCR扩增,获得大小为636 bp RSV NS3扩增产物(图1)。阳性样品均能扩增到目标大小的DNA片段,而阴性对照和空白对照均无扩增产物,说明PCR体系和反应程序没有问题,NS3引物没有污染。NS3-6株系的10个植株均能获得目标大小的DNA片段(图1-A),而NS3-9株系的10个植株中只有1个植株未能获得目标大小的DNA片段(图1-B),因此,所检测的NS3-6、NS3-9株系的阳性率分别为100%、90%。
A:转基因本氏烟NS3-6株系NS3-transgenic N. benthamiana line 6;B:转基因本氏烟NS3-9株系NS3-transgenic N. benthamiana line 9。M:Marker;a:阳性对照Positive control;b:阴性对照Negative control;c:ddH2O;1—10:1—10号样品The ten plants of transgenic N. benthamiana Fig. 1The identification of NS3-transgenic N. benthamiana by PCR
A:野生型本氏烟未接种PVX The health wild-type N. benthamiana plants; B:野生型本氏烟接种PVX Wild-type N. benthamiana inoculated with PVX;C:转基因本氏烟NS3-6株系接种PVX NS3-transgenic N. benthamiana line 6 inoculated with PVX;D:转基因本氏烟NS3-9株系接种PVX NS3-transgenic N. benthamiana line 9 inoculated with PVX Fig. 2The symptom of PVX-infecting N. benthamiana
COGONIC, MACINOG . Post-transcriptional gene silencing across kingdoms Current Opinion in Genetics and Development, 2000,10(6):638-643. DOI:10.1016/S0959-437X(00)00134-9URL [本文引用: 1]
AZEVEDOJ, GARCIAD, PONTIERD, OHNESORGES, YUA, GARCIAS, BRAUNL, BERGDOLLM, HAKIMI MA, LAGRANGET, VOINNETO . Argonaute quenching and global changes in Dicer homeostasis caused by a pathogen-encoded GW repeat protein Genes and Development, 2010,24(9):904-915. DOI:10.1101/gad.1908710URL [本文引用: 1]
CHENJ, LI WX, XIED, PENG JR, DING SW . Viral virulence protein suppresses RNA silencing-mediated defense but upregulates the role of microRNA in host gene expression The Plant Cell, 2004,16(5):1302-1313. DOI:10.1105/tpc.018986URL [本文引用: 1]
CHIU MH, CHEN IH, BAULCOMBE DC, TSAI CH . The silencing suppressor P25 of Potato virus X interacts with Argonaute1 and mediates its degradation through the proteasome pathway. Molecular Plant Pathology, 2010,11(5):641-649. [本文引用: 1]
MALLORY AC, ELYL, SMITH TH, MARATHER, ANANDALAKSHMIR, FAGARDM, VAUCHERETH, PRUSSG, BOWMANL, VANCE VB . HC-Pro suppression of transgene silencing eliminates the small RNAs but not transgene methylation or the mobile signal The Plant Cell, 2001,13(3):571-583. DOI:10.1105/tpc.13.3.571URL [本文引用: 1]
DUNOYERP, LECELLIER CH, PARIZOTTO EA, HIMBERC, VOINNETO . Probing the microRNA and small interfering RNA pathways with virus-encoded suppressors of RNA silencing The Plant Cell, 2004,16(5):1235-1250. DOI:10.1105/tpc.020719URL [本文引用: 1]
JAYF, WANGY, YUA, TACONNATL, PELLETIERS, COLOTV, RENOU JP, VOINNETO . Misregulation of AUXIN RESPONSE FACTOR 8 underlies the developmental abnormalities caused by three distinct viral silencing suppressors in Arabidopsis. PLoS Pathogens, 2011,7(5):e1002035. [本文引用: 1]
SHEN WJ, RUAN XL, LI XS, ZHAOQ, LI HP . RNA silencing suppressor Pns11 of Rice gall dwarf virus induces virus-like symptoms in transgenic rice. Archives of Virology, 2012,157(8):1531-1539. [本文引用: 1]
WU GT, WANG JY, YANGY, DONGB, WANG YL, SUN GC, YAN CQ, YANF, CHEN JP . Transgenic rice expressing Rice stripe virus NS3 protein, a suppressor of RNA silencing, shows resistance to rice blast disease. Virus Genes, 2014,48(3):566-569. [本文引用: 4]
WU GT, ZHENG GX, HUQ, MA MG, LI MJ, SUN XC, YANF, QINGL . NS3 protein from Rice stripe virus affects the expression of endogenous genes in Nicotiana benthamiana. Virology Journal, 2018,15:105. [本文引用: 4]
YANF, ZHANG HM, ADAMS MJ, YANGJ, PENG JJ, ANTONIW JF, ZHOU YJ, CHEN JP . Characterization of siRNAs derived from Rice stripe virus in infected rice plants by deep sequencing. Archive of Virology, 2010,155(6):935-940. [本文引用: 1]
ZHUY, HAYAKAWAT, TORIYAMAS . Complete nucleotide sequence of RNA 4 of Rice stripe virus isolate T, and comparison with another isolate and with Maize stripe virus. The Journal of General Virology, 1992,73(5):1309-1312. [本文引用: 1]
WU GT, LU YW, ZHENG HY, LINL, YANF, CHEN JP . Transcription of ORFs on RNA2 and RNA4 of Rice stripe virus terminate at an AUCCGGAU sequence that is conserved in the genus Tenuivirus. Virus Research, 2013,175(1):71-77. [本文引用: 1]
XIONG RY, WU JX, ZHOU YJ, ZHOU XP . Characterization and subcellular localization of an RNA silencing suppressor encoded by Rice stripe tenuivirus. Virology, 2009,387(1):29-40. [本文引用: 3]
XUY, WU JX, FUS, LI CY, ZHU ZR, ZHOU XP . Rice stripe tenuivirus nonstructural protein 3 hijacks the 26S proteasome of the small brown planthopper via direct interaction with regulatory particle non-ATPase subunit 3 Journal of Virology, 2015,89(8):4296-4310. DOI:10.1128/JVI.03055-14URL [本文引用: 1]
ZHENG LJ, ZHANGC, SHI CN, WANGY, ZHOUT, SUNF, WANGH, ZHAO SS, QIN QQ, QIAOR, DING ZM, WEI CH, XIE LH, WU JG, LIY . Rice stripe virus NS3 protein regulates primary miRNA processing through association with the miRNA biogenesis factor OsDRB1 and facilitates virus infection in rice PLoS Pathogens, 2017,13(10):e1006662. [本文引用: 1]
JIANG LL . Studies on interaction between RSV p3 protein and a host nucleus-localized protein Nbpip3 [D]. Nanjing: Nanjing Agricultural University, 2017. (in Chinese) [本文引用: 1]
DASGUPTAI, MALATHI VG, MUKHERJEE SK . Genetic engineering for virus resistance Current Science, 2003,84(3):341-354. [本文引用: 1]
RUBINOL, CAPRIOTTIG, LUPPOR, RUSSOM . Resistance to cymbidium ringspot tombusvirus infection in transgenic Nicotiana benthamiana plants exprssing the virus coat protein gene. Plant Molecular Biology, 1993,21(4):665-672. [本文引用: 1]
YU XH, ZHU YX, YING HC, CHEN ZL . Molecular cloning, nucleotide sequencing and plant transformation of a cDNA coding for the Tobacco mosaic virus movement protein. Chinese Journal of Biotechnology, 1996,12(2):211-214. (in Chinese) [本文引用: 1]
PRUSS GJ, LAWRENCE CB, BASST, LI QQ, BOWMAN LH, VANCEV . The potyviral suppressor of RNA silencing confers enhanced resistance to multiple pathogens Virology, 2004,320(1):107-120. DOI:10.1016/j.virol.2003.11.027URL [本文引用: 3]
ZHENG GX . The study about prokaryotic expression of p3 from Sichuan RSV isolate and p3 affects on endogenous gene expression of Nicotiana benthamiana [D]. Chongqing: Southwest University, 2018. (in Chinese) [本文引用: 1]
WANGY, XIAO CG . Isolation of avirulent strains of bacterial wilt of tomato (Ralstonia solanacearum) and its application in biocontrol of the disease. Journal of Southwest Agricultural University (Natural Science), 2004,26(4):426-428. (in Chinese) [本文引用: 1]
WANG HB . Study on mutation breeding of two antagonistic bacterium strains against Phytophthora parasitica and Ralstonia solanacearum [D]. Chongqing: Southwest University, 2009. (in Chinese) [本文引用: 1]
马国胜, 高智谋 . 烟草黑胫病菌培养性状的研究 中国农业科学, 2007,40(3):512-517. Magsci [本文引用: 1] 【目的】对烟草黑胫病菌的若干培养性状进行研究,进一步了解烟草黑胫病菌的培养特性的稳定性,为烟草黑胫病的相关研究和综合治理提供试验依据。【方法】从安徽10个主要生产烟草的市(县)多点采集发生烟草黑胫病的烟株,采用组织分离法进行病原菌分离,经纯化后获得69个烟草黑胫病菌(Phytophthora nicotianae Breda de Haan var. nicotianae Waterh.)菌株,对其进行游动孢子囊的诱导、配对培养、生长特性测定等进行研究。【结果】烟草黑胫病菌的主要培养性状为:异宗配合,与标准相对交配型菌株配对培养可以产生有性器官;卵孢子球形,满器或不满器,雄器围生,藏卵器小;孢子囊多近球形,乳突明显,有双乳突现象,孢子囊具脱落性,且孢子囊柄短,不对称孢子囊较常见,孢子囊外着生有长短不一的附属丝;菌丝最适生长温度为20~30℃,最高生长温度大于35℃,小于40℃,最低生长温度大于8℃,小于10℃。【结论】供试烟草黑胫病菌的培养性状与前人的研究结论基本一致,初步认为10多年来生态环境和农业措施的变化没有导致烟草黑胫病菌的生长性状发生较大的改变,表明烟草黑胫病菌的培养性状是较为稳定的。 MA GS, GAO ZM . Cultural characteristics of Phytophthora nicotianae var. nicotianae pathogen of tobacco blank shank. Scientia Agricultura Sinica, 2007,40(3):512-517. (in Chinese) Magsci [本文引用: 1] 【目的】对烟草黑胫病菌的若干培养性状进行研究,进一步了解烟草黑胫病菌的培养特性的稳定性,为烟草黑胫病的相关研究和综合治理提供试验依据。【方法】从安徽10个主要生产烟草的市(县)多点采集发生烟草黑胫病的烟株,采用组织分离法进行病原菌分离,经纯化后获得69个烟草黑胫病菌(Phytophthora nicotianae Breda de Haan var. nicotianae Waterh.)菌株,对其进行游动孢子囊的诱导、配对培养、生长特性测定等进行研究。【结果】烟草黑胫病菌的主要培养性状为:异宗配合,与标准相对交配型菌株配对培养可以产生有性器官;卵孢子球形,满器或不满器,雄器围生,藏卵器小;孢子囊多近球形,乳突明显,有双乳突现象,孢子囊具脱落性,且孢子囊柄短,不对称孢子囊较常见,孢子囊外着生有长短不一的附属丝;菌丝最适生长温度为20~30℃,最高生长温度大于35℃,小于40℃,最低生长温度大于8℃,小于10℃。【结论】供试烟草黑胫病菌的培养性状与前人的研究结论基本一致,初步认为10多年来生态环境和农业措施的变化没有导致烟草黑胫病菌的生长性状发生较大的改变,表明烟草黑胫病菌的培养性状是较为稳定的。
WANGY . Screening and biocontrol effect of antagonistic actinomycetes against Phytophthora parasitica and Ralstonia solannacearum [D]. Chongqing: Southwest University, 2009. (in Chinese) [本文引用: 1]
KASSCHAU KD, XIEZ, ALLENE, LLAVEC, CHAPMAN EJ, KRIZAN KA, CARRINGTON JC . P1/HC-Pro, a viral suppressor of RNA silencing, interferes with Arabidopsis development and miRNA function. Developmental Cell, 2003,4(2):205-217. [本文引用: 1]
TAKAGIK, NISHIZAWAK, HIROSEA, KURAUCHIT, SENDAM, MASUTAC, ISHIMOTOM . Seed coat pigmentation in transgenic soybean expressing the silencing suppressor 2b gene of Cucumber mosaic virus. Plant Cell Reports, 2013,32(12):1903-1912. [本文引用: 1]
LIU YL, WANG ZD, ZHUF, KANG LY, TIANB . Tolerance of the transgenic tobacco expressing satellite RNA of Cucumber mosaic virus to Tobacco mosaic virus. Acta Microbiologica Sinica, 1994,34(5):403-405. (in Chinese) [本文引用: 1]
刘晓玲, 宋云枝, 刘红梅, 温孚江, 朱常香, 白庆荣 . 马铃薯X病毒25 kD运动蛋白基因和外壳蛋白基因介导的抗病性研究 作物学报, 2005,31(7):827-832. Magsci [本文引用: 1] 以马铃薯X病毒(potato virus X,PVX)的RNA为模板,应用反转录-聚合酶链式反应(RT-PCR)方法分别扩增出长度为681 bp的非翻译马铃薯X病毒25 kD运动蛋白基因(PVX-p25)和长度为714 bp的非翻译马铃薯X病毒外壳蛋白基因(PVX-CP)。并分别构建植物表达载体pROKⅡ-p25和pROKⅡ-CP。利用农杆菌介导方法转化烟草NC89。经卡那霉 LIU XL, SONG YZ, LIU HM, WEN FJ, ZHU CX, BAI QR . Virus resistance mediated by the cDNAs encoding for the movement and coat proteins of Potato Virus X. Acta Agronomica Sinica, 2005,31(7):827-832. (in Chinese) Magsci [本文引用: 1] 以马铃薯X病毒(potato virus X,PVX)的RNA为模板,应用反转录-聚合酶链式反应(RT-PCR)方法分别扩增出长度为681 bp的非翻译马铃薯X病毒25 kD运动蛋白基因(PVX-p25)和长度为714 bp的非翻译马铃薯X病毒外壳蛋白基因(PVX-CP)。并分别构建植物表达载体pROKⅡ-p25和pROKⅡ-CP。利用农杆菌介导方法转化烟草NC89。经卡那霉
GUO XQ, HAN HY, ZHANG JD, WANG HG . The viral resistance of transgenic tobacco plants containing untranslatable PVY N coat protein gene Acta Biologiae Experimentalis Sinica, 2003,36(3):176-184. (in Chinese) [本文引用: 1]
ZHANGG, CHENM, LIL, XUZ, CHENX, GUOJ, MAY . Overexpression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco Journal of Experimental Botany, 2009,60(13):3781-3796. DOI:10.1093/jxb/erp214URL [本文引用: 1]
ZHANGG, CHENM, CHENX, XUZ, LIL, GUOJ, MAY . Isolation and characterization of a novel EAR-motif-containing gene GmERF4 from soybean(Glycine max L.). Molecular Biology Reports, 2010,37(2):809-818. [本文引用: 1]
JAYNES JM, NAGPALAP, DESTEFANO-BELTRANL, HUANG JH, KIMJ, DENNYT, CETINERS . Expression of a Cecropin B lytic peptide analog in transgenic tobacco confers enhanced resistance to bacterial wilt caused by Pseudomonas solanacearum. Plant Science, 1993,89(1):43-53. [本文引用: 1]
 ,西南大学植物保护学院植物病害生物学重庆市高校级重点实验室,重庆 400716
,西南大学植物保护学院植物病害生物学重庆市高校级重点实验室,重庆 400716
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT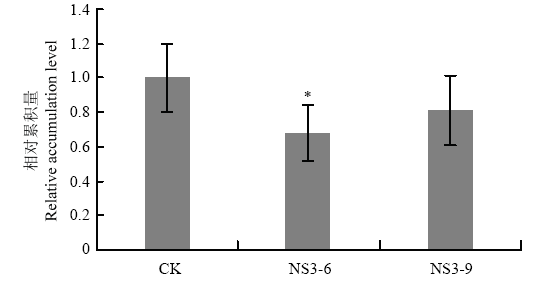 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT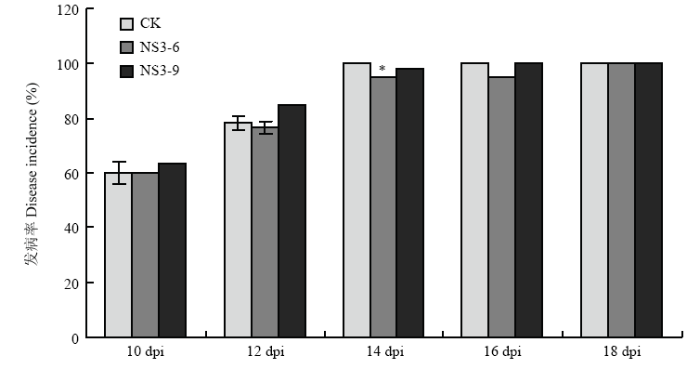 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT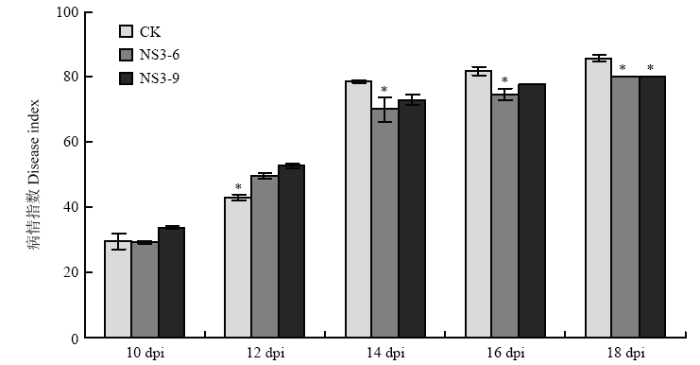 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT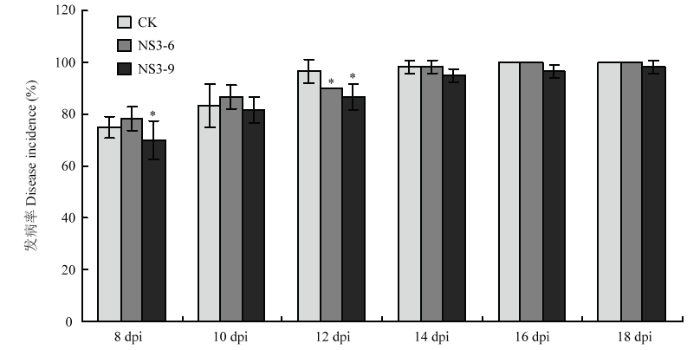 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT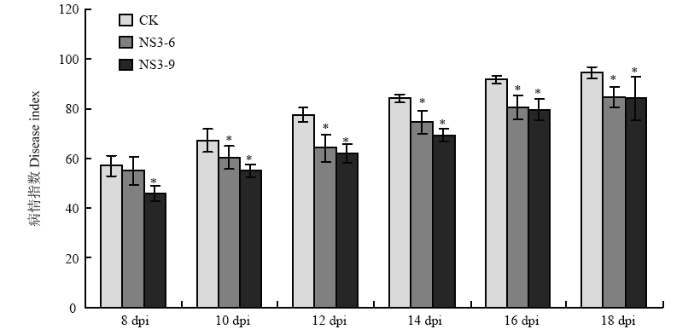 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}