 ,1,2
,1,2Screening and Identification of Candidate Proteins Interacting with BmHSP60 in the Silkworm (Bombyx mori)
DONG ZhanQi1, JIANG YaMing1, PAN MinHui,1,2通讯作者:
责任编辑: 岳梅
收稿日期:2018-08-8接受日期:2018-09-6网络出版日期:2019-01-16
| 基金资助: |
Received:2018-08-8Accepted:2018-09-6Online:2019-01-16
作者简介 About authors
董战旗,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1322KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
董战旗, 蒋亚明, 潘敏慧. 家蚕热休克蛋白HSP60相互作用蛋白筛选与鉴定[J]. 中国农业科学, 2019, 52(2): 376-384 doi:10.3864/j.issn.0578-1752.2019.02.016
DONG ZhanQi, JIANG YaMing, PAN MinHui.
0 引言
【研究意义】热休克蛋白(heat shock protein,HSP)最早在果蝇(Drosophila melanogaster)中被发现[1]。生物体不仅在热激条件下能够产生热休克反应,在受到其他物理、化学或生物刺激下,也能够发生相关应答,提高热休克蛋白的表达[2]。热休克蛋白可帮助蛋白质正确折叠、在各种应激条件下调节蛋白质稳态,因此在保护细胞免受氧化压力、极端温度和凋亡损伤中发挥重要作用,在组织中也可消除由疾病引起的损伤[1]。热休克蛋白广泛存在于各种生物中,根据分子大小,可分为HSP110(HSPH)、HSP90(HSPC)、HSP70(HSPA)、HSP40(DNAJ)和小分子热休克蛋白(HSPB)以及伴侣家族HSPD/E(HSP60/HSP10)[3]。HSP60是热休克家族重要成员,广泛参与先天性免疫和细胞凋亡过程,前期研究证明BmHSP60作为线粒体蛋白在家蚕核型多角体病毒(Bombyx mori nucleopolyhedrovirus,BmNPV)侵染过程中为病毒提供能量促进病毒增殖复制,但BmHSP60在家蚕个体发育及BmNPV入侵过程中具体的作用机制仍然未知[4]。因此,鉴定BmHSP60相互作用蛋白对解析其在先天性免疫中的功能以及病毒和宿主相互作用机制解析具有重要意义。【前人研究进展】HSP60功能强大,参与宿主很多途径,并参与病原入侵的过程。人乙肝病毒(Hepatitis B virus,HBV)聚合酶已被证明能够与HSP60发生相互作用,利用了HSP60分子伴侣的功能,确保聚合酶折叠成活化形态,ATP也辅助参与了这个过程,抑制HSP60表达能够有效抑制HBV的复制。在此前的研究中,HBV转录反式激活蛋白也被证明能够与线粒体HSP60形成一个复合物,促进病毒的感染[5]。同时,在对人乳头瘤病毒(Human papilloma virus,HPV)引起的宫颈癌的治疗中,DNA疫苗编码的HSP60能够与HPV16的E6和E7抗原结合,并将其呈递到特异型的CD8+ T细胞,免疫效果比单纯的抗原呈递明显,促进了体液免疫[6]。除了直接与病毒蛋白质发生相互作用,HSP60也能够直接与病毒的基因组发生相互作用,在小鼠肝炎病毒(Murine hepatitis virus,MHV)侵染过程中,HSP60连同HSP70、HSP40共同作用,与MHV的RNA 3′末端42个核苷酸结合,形成RNA-蛋白质复合结构,该结构被认为与RNA病毒的复制相关[7]。此外,病毒也能够利用HSP60所在的通路,如人丙肝病毒(Hepatitis C virus,HCV)核心蛋白能够与HSP60相互作用,提高了细胞活性氧含量,使之对肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)变得更加敏感,反之,过表达HSP60能够在一定程度恢复细胞对TNF-α的耐受力[8]。热休克蛋白HSP60广泛存在于各种昆虫中,在昆虫各个时期、各种组织中均有表达[1]。HSP60与昆虫个体发育息息相关,伴随着组织的分解与再生,对保持蛋白质的正确折叠和维持细胞蛋白质稳态非常重要[2]。西方蜜蜂(Apis mellifera)在社会化发展过程中,蜂王幼虫中HSP60表达高于工蜂幼虫,从而造成蜂王和工蜂的卵巢发育程度不同[9];在果蝇中,HSP60参与了Caspase介导的细胞凋亡,作为果蝇细胞线粒体抗氧化、抗衰老以及重建蛋白质平衡的标志,为昆虫适应极端环境提供了一定的分子基础[10]。【本研究切入点】笔者课题组前期研究已经证明杆状病毒晚期表达因子11(late expression factor 11,LEF-11)蛋白能够劫持宿主ATPase家族BmATAD3A和伴侣蛋白BmHSP60促进病毒增殖复制,干涉BmHSP60后能够显著抑制病毒增殖复制[4]。但是BmHSP60促进病毒增殖复制的调控机制仍然未知。【拟解决的关键问题】通过筛选和鉴定BmHSP60具有直接相互作用的蛋白,找到与BmHSP60在病毒感染过程中相互作用蛋白,为其在家蚕先天性免疫中作用机制研究打下基础,为家蚕抗病育种研究提供靶标基因。1 材料与方法
试验于2015—2017在西南大学家蚕基因组生物学国家重点实验室完成。1.1 细胞和培养基
家蚕胚胎细胞系BmN-SWU1细胞和BmNPV病毒为西南大学家蚕基因组生物学国家重点实验室保存,BmN-SWU1细胞用含10%(V/V)胎牛血清TC-100培养基,在27℃条件下恒温培养[11,12]。1.2 质粒构建
以pIZ/V5-His为空载,通过PCR获得家蚕BmHSP60、BmANT1、BmTubulin、BmEF-1α和BmHSP90开放阅读框(open reading frame,ORF),分别通过酶切、连接构建不同的带有Flag和HA标签的质粒。所有构建质粒都经测序分析,所用引物见表1。Table 1
表1
表1本试验所用引物
Table 1
| 引物名称Primer name | 引物系列Primer sequence (5′-3′) |
|---|---|
| BmHSP60 Flag-BamH I | CGCGGATCCATGGACTACAAAGACGATGACGACAAGTTGCGTCTACCTCGTGTT |
| BmHSP60Flag-Not I | ATAAGAATGCGGCCGCTCACTTGTCGTCATCGTCTTTGTAGTC CATCATGCCTCCCATACCA |
| BmANT1HA-KpnI-F | GGGGTACCATGTACCCATACGACGTCCCAGACTACGCTTCGAACCTCGCCGA |
| BmANT1 HA-SpeI-R | GGACTAGTGGCGTAGTCGGGCACGTCGTAGGGGTACAGGACCTTCTTGATCTCAT |
| BmTubulin HA-KpnI-F | GGGGTACCTACCCATACGACGTCCCAGACTACGCTATGAGGGAATGCATCTCC |
| BmTubulin HA-SpeI-R | GGACTAGTGGCGTAGTCGGGCACGTCGTAGGGGTAATATTCATTTTCTTCGTCCAACT |
| BmEF-1αHA-KpnI-F | GGGGTACCTACCCATACGACGTCCCAGACTACGCTATGGGTAAAGAGAAGACTCATATCA |
| BmEF-1αHA-SpeI-R | GGACTAGTGGCGTAGTCGGGCACGTCGTAGGGGTATTTTTTCTTCTGGGCCTTCT |
| BmHSP90 HA-SpeI-F | GGACTAGTTACCCATACGACGTCCCAGACTACGCTATGAAGGTTGAAACCTTCAC |
| BmHSP90 HA-XhoI-R | CCGCTCGAGGGCGTAGTCGGGCACGTCGTAGGGGTACTGAATCGGCTCATCTT |
新窗口打开|下载CSV
1.3 免疫共沉淀
当细胞铺满80%左右时,按照4 μg质粒,12 μL转染试剂,在离心管中轻柔逐滴混匀50次左右,静止30 min,然后滴加到细胞瓶中。转染48 h,添加BmNPV 48 h后,用PBS轻轻润洗两次,在细胞瓶中加入1 mL含有PMSF的Western及IP细胞裂解液,冰上裂解30 min。取两只灭菌的1.5 mL离心管,分别加入50 μL DynabeadsTM Protein A磁珠,安置在磁力架上,弃去液体,加入少许PBS混匀清洗,重复一次,然后加入200 μL PBS,一只加入3 μL Flag标签抗体,另外一只加入等量的小鼠IgG,做好标记,并安装在翻转仪上,在4℃环境中,与磁珠孵育1 h。将抗体孵育完成的磁珠安装在磁力架上,弃去液体,同时将细胞裂解产物取100 μL作为Input样品,放在冰上,剩下产物各取450 μL,分装到两管,与磁珠轻柔混匀后,放置在翻转仪上,4℃环境中孵育1 h。将与磁珠孵育完成的样品从翻转仪上取下,安装在磁力架上,弃去液体后,加入300 μL左右PBS,轻轻混匀,清洗磁珠,并在磁力架上分离液体与磁珠,弃去液体,如此重复两次,最后弃去洗液。然后每只离心管加入60 μL含有PMSF的Western及IP细胞裂解液,再加入15 μL的5×SDS-PAGE蛋白上样缓冲液,最后将样品沸水浴10 min。将沸水浴后的样品安装在磁力架上,分离液体和磁珠,将液体转移至新的离心管中,做好标记,和Input样品一同保存在-20℃。1.4 Western blot
制备12%的SDS-PAGE胶,将蛋白样品轻轻加入到点样孔中,同时点好相应的蛋白Marker,开始时浓缩蛋白用7 mA/块胶,待到蛋白完全通过浓缩胶后,将电流提高到15 mA/块,保持电流稳定电泳3 h左右,待蓝色染料刚好跑出即可停止电泳[13,14]。电泳完成后,把浓缩胶及以上部分切掉,并将分离胶从玻璃板上剥下,放置在事先加入dH2O的玻璃皿中轻摇,清洗一次。倒掉洗液,向玻璃皿中加入适量银染固定液,使其能够浸没胶块,在摇床上慢速轻摇1 h。倒掉固定液,向玻璃皿中加入适量银染清洗液,脱色摇床上轻摇1 h。倒掉清洗液,避光条件下,向玻璃皿中加入适量银染致敏液,增感2 min,然后倒掉致敏液,再向里面加入适量dH2O,润洗20 s后倒掉,重复dH2O清洗3次。避光条件下,向玻璃皿中加入适量银染染色液,在水平摇床上轻摇30 min,然后倒掉染色液,如上方法,用dH2O润洗20 s,重复3次。清洗完成后,向玻璃皿中倒入适量银染显色液,时刻观察胶块上条带染色情况,若染色完成,立即将胶块转移至装有适量银染终止液的玻璃皿中,终止显色反应。将显色完成后的胶块平放在扫描仪上,轻轻赶走产生的气泡,根据软件提示,扫面胶块并保存结果。根据差异条带位置,用甲醛消毒过的刀切掉差异条带,-80℃保存。1.5 质谱分析
通过硝酸银染色获取的凝胶样品管中加入200—400 μL脱色液(30% ACN/100 mmol·L-1 NH4HCO3),清洗脱色至透明,吸弃上清,冻干;加入100 mmol·L-1 DTT,56℃孵育30 min;吸弃上清,加入200 mmol·L-1 IAA,暗处孵育20 min。吸弃上清,加入100 mmol·L-1 NH4HCO3,室温孵育15 min。吸弃上清,加入100% ACN,5 min后吸弃冻干,加入2.5—10 ng·μL-1 Trypsin溶液,在37℃条件下反应20 h左右。转移酶解原液至新的EP管中,之后凝胶中加入100 μL抽提液(60% ACN/0.1% TFA),超声15 min后将提取液与酶解原液合并,冻干后取出。加入0.1% FA水溶液60 μL进行复溶,再用0.22 μm过滤管过滤后待用。色谱柱以95%的0.1%甲酸的水溶液平衡后,样品由自动进样器上样至Trap柱。色谱梯度:0—50 min,0.1%甲酸的乙腈水溶液(乙腈为84%)线性梯度从4%—50%;50—54 min,0.1%甲酸的乙腈水溶液线性梯度从50%—100%;54—60 min,0.1%甲酸的乙腈水溶液维持在100%。多肽和多肽碎片的质量电荷比按照下列方法采集:每次全扫描(full scan)后采集10个碎片图谱(MS2 scan)。质谱测试原始文件(raw file)用Mascot2.2软件检索相应的数据库,最后得到鉴定的蛋白质结果。1.6 免疫荧光
按照1.3所述方法转染48 h后,用1×PBST轻洗细胞3次,每次5 min。加入4%的多聚甲醛溶液,室温固定15 min,1×PBST同上轻洗。然后加入300 μL 1% Triton X-100,室温打孔15 min。PBS清洗两次后,加入500 μL封闭液,于37℃培养箱中封闭1 h。吸去封闭液,每孔加入配好的1﹕200的Flag/HA一抗,37℃孵育1 h。回收一抗,每孔加入适量PBS,在水平摇床上轻摇清洗,5 min更换一次PBS,重复6次。根据一抗来源,向孔里加入500 μL 1﹕500稀释的荧光标记二抗,37℃孵育1 h。向孔中加入100 μL DAPI染色液或者1﹕500稀释的 Mito-Tracker Green,37℃孵育15 min。吸取染色液,重复清洗步骤。用带钩的针头挑出扒片,将细胞一面平铺到滴有甘油的载玻片上,用指甲油封片。用荧光共聚焦显微镜进行观察。2 结果
2.1 免疫共沉淀钓取BmHSP60相互作用候选蛋白
将新提取的pIZ-BmHSP60Flag质粒转染到生长状态良好的细胞中,转染后加入BmNPV 48 h后收取蛋白。根据免疫共沉淀收方法用Flag抗体孵育蛋白细胞提取液,然后进行蛋白质电泳和银染。生物学重复两次,选取两次试验同时具有差异的条带,分别在110、90、75、60和35 kD位置处选取5个差异条带进行切胶,命名为差异条带A、B、C、D和E(图1)。把各差异条带混在一起-80℃保存,以备质谱分析。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1免疫共沉淀钓取BmHSP60相互作用候选蛋白
免疫共沉淀检测BmHSP60相互作用蛋白Co-IP assays of BmHSP60- associated protein analyzed by SDS-PAGE。Marker:蛋白分子量标记Protein molecular weight marker;Input:细胞裂解物Input cell lysates;IgG:免疫共沉淀对照小鼠抗体IP with control mouse IgG;α-Flag:免疫共沉淀用Flag抗体IP with Flag antibody。特异条带用箭头表示The specific bands are represented by the arrows
Fig. 1Co-immunoprecipitation for BmHSP60 interaction candidate proteins
2.2 LC-MS/MS分析BmHSP60候选相互作用蛋白
为了筛选不同分子量的BmHSP60相互作用蛋白,将两次重复同时具有差异条带的5条特异条带混为蛋白池,进行LC-MS/MS质谱分析,将质谱分析多肽与家蚕数据库以及NCBI数据库进行比对。根据候选相互作用蛋白相对分子质量大小和多肽的数量进行排序,共找到5个潜在的相互作用蛋白,分别为ADP/ATP translocase (ANT)、Actin、Alpha-tubulin、Elongation factor 1-alpha(EF1α)和HSP90,命名为BmANT1、BmActin、BmTubulin、BmEF-1α和BmHSP90(表2)。Table 2
表2
表2LC-MS/MS分析BmHSP60相互作用蛋白
Table 2
| 蛋白ID Protein ID | 描述 Description | 分子量Molecular weight (kD) | 多肽数量 Peptide number | 功能 Function |
|---|---|---|---|---|
| NP_001037072.1 | ADP/ATP translocase (ANT1) | 32.9 | 13 | 参与ADP/ATP的转运、线粒体稳态以及细胞凋亡Involved in ADP/ATP transport, mitochondrial homeostasis and apoptosis |
| NP_001119727.1 | Actin | 41.8 | 12 | 肌动蛋白是高度保守的蛋白质,参与各种类型的细胞运动,并在所有核细胞中普遍表达Actin is a highly conserved protein that is involved in various types of cell motility and ubiquitously expressed in all eukaryotic cells |
| NP_001036884.1 | Alpha-tubulin | 50.0 | 8 | 微管蛋白是微管的主要成分,它结合两个摩尔的GTP,一个在β链上的可交换位点,另一个在α链上的不可交换位点Tubulin is the major constituent of microtubules. It binds two moles of GTP, one at an exchangeable site on the beta chain and one at a non-exchangeable site on the alpha chain |
| NP_001037510.1 | Elongation factor 1-alpha (EF1α) | 50.3 | 7 | 促进蛋白质生物合成过程中氨基酰-tRNA与核糖体A位点的GTP依赖性结合Promoting the GTP-dependent binding of aminoacyl-tRNA to the A-site of ribosomes during protein biosynthesis |
| AEB39782.1 | HSP90, partial | 90.0 | 4 | 促进特定靶蛋白的成熟、结构维持和适当调节的分子伴侣,例如参与细胞周期控制和信号转导Molecular chaperone that promotes the maturation, structural maintenance and proper regulation of specific target proteins, for instance involved in cell cycle control and signal transduction |
新窗口打开|下载CSV
2.3 免疫共沉淀验证BmHSP60相互作用蛋白
克隆BmHSP60相互作用候选蛋白基因,构建到含有HA标签的pIZ/V5-His载体上,与pIZ-BmHSP60Flag共同转染到家蚕细胞,48 h后收取蛋白。因为Actin可能为细胞骨架蛋白,在多次免疫共沉淀中均有发现,因此暂不考虑。免疫共沉淀结果显示immunoprecipitated(IP)用α-Flag抗体孵育,immunoblotting(IB)再用α-Flag抗体孵育,均能够检测到BmHSP60表达,说明免疫共沉淀试验操作成功。如果IB用HA抗体进行孵育,均不能测到BmTubulin、BmEF-1α,但是能够检测到BmHSP90、BmANT1(图2)。上述结果说明BmHSP60不与BmTubulin、BmEF-1α结合,但可与BmHSP90、BmANT1结合。图2
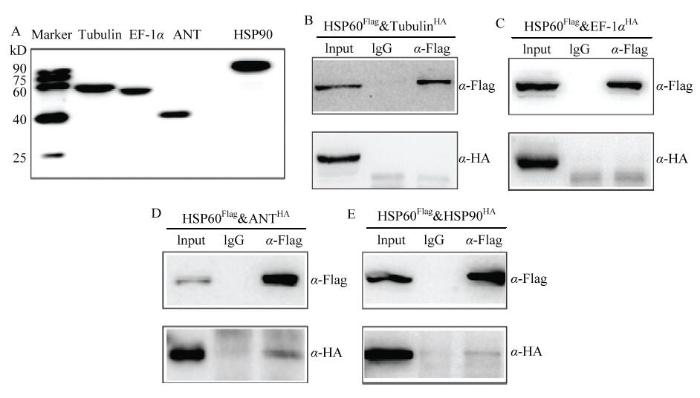 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2免疫共沉淀验证BmHsp90、BmANT1与BmHsp60相互作用
A:通过Western blot分析BmHSP60相互作用蛋白表达The expression of candidate interaction proteins of BmHSP60 by Western blot;B—E:通过Western blot分析BmHSP60相互作用蛋白。BmN-SWU1细胞共同转染BmHSP60和BmTubulin、BmEF-1α、BmHSP90、BmANT1。转染96 h后,收集细胞用Flag抗体共沉淀蛋白,靶蛋白用HA抗体检测。每个相互作用顶部表示免疫沉淀所用抗体,右侧标记显示Western blot分析所用抗体
Fig. 2Western blot analysis determining the interaction between BmHsp60 with BmHsp90 and BmANT1
Co-immunoprecipitation of BmHSP60 examined by Western blot. BmN-SWU1 cells are co-transfected with BmHSP60 and candidate BmTubulin, BmEF-1α, BmHSP90, BmANT1 protein. At 96 hours after transfection, cells are lysed and immunoprecipitation performed with α-FLAG, and use α-HA to detect the bound of target protein. The label on the top of each panel shows the antibodies used for immunoprecipitation. The label on the right of each panel shows the antibodies used for analysis of Western blot
2.4 荧光共定位验证BmHSP60相互作用蛋白
为了进一步探究BmHsp60及其相互作用蛋白BmHsp90、BmANT1在细胞中共定位情况,共转后通过免疫荧光步骤进行处理,荧光共定位结果显示,过表达BmANT1、BmHSP90 与BmHSP60在细胞中能够完全重叠,绿色代表HA标签,红色代表Flag标签,蓝色DAPI代表细胞核,共定位显示红色几乎和绿色完全重合,并且全部分布在细胞质中(图3),进一步说明它们之间可能存在相互作用。图3
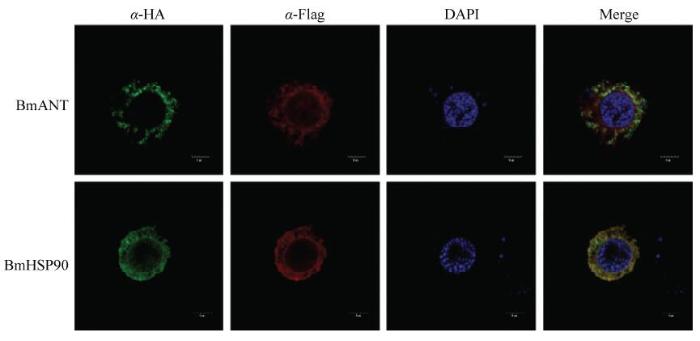 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3免疫荧光验证BmHSP60和BmHSP90、BmANT1相互作用
在BmN-SWU1细胞中转染BmHSP60和BmHSP90、BmANT1 48 h后,用Alexa 555标记抗Flag,FITC标记抗HA和Hoechst33258染色。红色荧光表示Flag,绿色荧光表示HA,蓝色荧光表示细胞核。比例尺Scale bar:5 μm
Fig. 3Immunofluorescence verification of interaction between BmHSP60 with BmHSP90 and BmANT1
Co-localization of BmHSP60 with BmHSP90 and BmANT1 in BmN-SWU1 cells. Alexa 555 labels anti-Flag, FITC labels anti-HA and Hoechst33258 at 48 h post-transfection in the BmN-SWU1 cells. Red fluorescence represents Flag, green fluorescence represents HA, and blue fluorescence represents the nucleus
2.5 线粒体共定位分析
为了探究BmHSP60及其相互作用蛋白BmANT1和BmHSP90亚细胞定位情况,分别转染后,通过免疫荧光步骤进行处理,同时对线粒体用Mito-Tracker- Green染成绿色,目的蛋白染成红色,荧光结果显示,过表达BmHSP60、BmANT1细胞中,红色几乎和绿色共定位在一起;过表达BmHSP90中,红色与绿色大部分共定位在一起,说明而BmHSP60、BmANT1参与线粒体代谢过程,BmHSP90也与线粒体代谢相关(图4)。图4
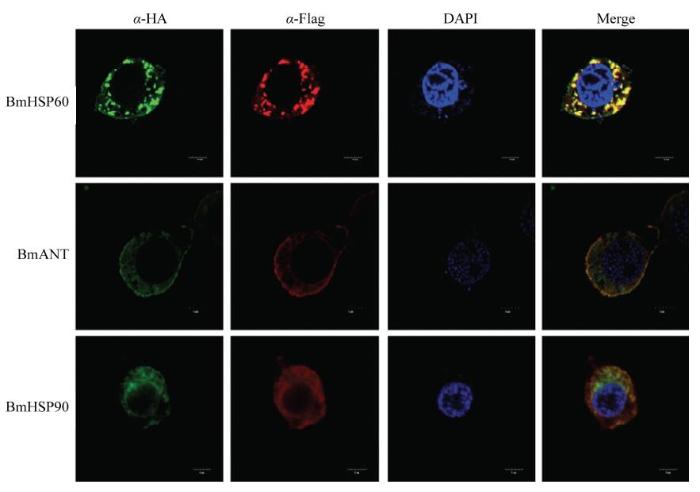 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4BmHSP60、BmANT1和BmHSP90线粒体共定位分析
在BmN-SWU1细胞中转染BmHSP60、BmANT1和BmHSP90 48 h后,用Alexa 555标记,线粒体示踪和Hoechst33258染色。红色荧光表示BmHSP60、BmANT1和BmHSP90,绿色荧光表示Mito-Tracker,蓝色荧光表示细胞核。比例尺Scale bar:5 μm
Fig. 4Mitochondria co-localization analysis of BmHSP60, BmANT1 and BmHSP90
BmHSP60, BmANT1 and BmHSP90 are stained with Alexa 555-labeled, mitochondria-tracker and Hoechst33258 at 48 h post-transfection in the BmN-SWU1 cells. Red fluorescence represents BmHSP60, BmANT1 and BmHSP90, green fluorescence represents Mito-Tracker, and blue fluorescence represents the nucleus
3 讨论
本研究采用免疫共沉淀和荧光共定位技术,筛选了BmHSP60在BmNPV感染过程中的相互作用候选蛋白,并验证了BmHSP60与BmHSP90和BmANT1具有相互作用,这为后期研究BmHSP60在家蚕个体发育中的功能及其在BmNPV增殖复制中作用机制提供了基础数据。以BmHSP60为诱饵蛋白钓取的相互作用蛋白中,BmHSP90和BmANT1均为机体的管家蛋白,HSP90作为另一类热休克蛋白,同其他热激蛋白家族一样,与病毒有着复杂的关系[15,16]。HSP90既能通过信号转导途径影响病毒的复制增殖,也能直接作用于病毒的蛋白[17,18]。研究表明,HSP90通过抑制牛痘病毒核心蛋白4a的活性抑制其复制,HSP90通过结合流感病毒PB2蛋白调节病毒聚合酶的活性促进其复制[19,20]。HSP90也能够帮助流感病毒DNA聚合酶的组装,并且能协助其入核[19,21]。目前报道与HSP90相关的病毒涉及双链DNA病毒、单链DNA病毒、RNA病毒等,对于不同的病毒HSP90所起作用也不尽相同,有促进病毒增殖的作用,也有起免疫作用的[17]。先前已被报道与HSP60能够一起作为分子伴侣形成一个大的复合物,抑制细胞线粒体凋亡[22],因此它们可能在病毒入侵时通过抑制宿主细胞凋亡,进而共同调控BmNPV入侵。笔者课题组前期研究也同样证明LEF-11能够劫持宿主ATPase家族BmATAD3A和BmHSP60促进自身增殖复制,这些结果都暗示BmHSP60在家蚕抗病毒免疫的热休克反应和线粒体代谢过程具有一定的作用[23]。
包括家蚕在内的很多昆虫都具有两个ANT同源基因,分别命名为ANT1和ANT2,ANT1在各组织中均有高量表达,而ANT2只特异地在精巢中表达,与生殖细胞的形成有关[24]。已有报道宿主的ANT1能够和病原的相关蛋白发生相互作用。猪繁殖与综合呼吸征病毒包膜蛋白E、对虾白斑综合症病毒VP12以及人类免疫缺陷病毒Vpr均能和宿主ANT1相互作用,控制线粒体膜通透性[25,26]。此外,研究表明病毒的RNA也能干涉宿主ANT基因,人类巨细胞病毒miR-UL36-5P与ANT-3的mRNA作用,下调ANT-3的表达从而抑制凋亡[27]。BmNPV IE-2相互作用蛋白鉴定中,同样发现IE-2相互作用候选蛋白包括ANT1,为ANT1参与BmNPV复制进一步提供了依据[28],因此,笔者推测BmANT1也很有可能与病毒蛋白直接相互作用调控病毒增殖复制。不同昆虫的ANT1存在相似的结构,即含有3个线粒体跨膜结构域,锚定在线粒体内膜中,与电压依赖性阴离子通道(voltage- dependent anion channel,VDAC)一同构成了线粒体膜孔复合物,参与调节线粒体内膜通透性,电化学平衡和氧化磷酸化链,甚至参与细胞凋亡途径[28],因此根据它们定位于线粒体,可以推测家蚕BmHSP60和BmANT1在BmNPV诱导线粒体凋亡信号通路中具有非常关键的作用,从而促进病毒增殖复制。
BmHSP90和BmANT1蛋白均能够定位在线粒体上,这与BmHSP60在线粒体中的功能相吻合[29]。下一步将集中于研究BmHSP90和BmANT1蛋白如何影响病毒增复制以及与病毒直接的调控关系,为BmNPV劫持宿主能量代谢提供更深入的研究,完善病毒和宿主相互博弈网络。
4 结论
通过免疫共沉淀和细胞共定位鉴定出BmHSP60相互作用蛋白为BmHSP90和BmANT1,这两个蛋白与BmHSP60共定位在细胞线粒体,为研究BmHSP60在家蚕抗病毒免疫中的作用机制打下了基础。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
DOI:10.1038/293311a0 [本文引用: 2]
[本文引用: 1]
[本文引用: 2]
DOI:10.1007/s00705-005-0521-1URL [本文引用: 1]
DOI:10.1016/j.ygyno.2007.06.031URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1371/journal.pone.0144930URL [本文引用: 1]
DOI:10.1016/j.antiviral.2014.01.017URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/(ISSN)1096-9071URL [本文引用: 2]
DOI:10.1016/j.virusres.2016.06.003URL [本文引用: 1]
DOI:10.1074/jbc.M206822200URL [本文引用: 2]
DOI:10.1128/JVI.76.3.1379-1390.2002URL [本文引用: 1]
DOI:10.1016/j.cimid.2015.10.001URL [本文引用: 1]
DOI:10.1016/j.fsi.2015.03.027URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00705-016-2845-4URL [本文引用: 1]
DOI:10.1371/journal.ppat.0020127URL [本文引用: 1]
DOI:10.1038/sj.cdd.4401089URL [本文引用: 1]
[本文引用: 2]
DOI:10.1016/j.bbrc.2016.10.153URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}