 , 范方军
, 范方军Genetic Analysis of Imazethapyr Resistance in Rice and the Closely Linked Marker Selection and Application
FEIYun-Yan, FANFang-Jun通讯作者:
收稿日期:2017-07-6
接受日期:2018-01-8
网络出版日期:2018-01-23
版权声明:2018作物学报编辑部作物学报编辑部
基金资助:
作者简介:
-->
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (1348KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
草害是影响水稻产量的主要因素之一。杂草不仅与作物争夺水分、光照等, 同时易于寄生病虫害, 影响作物生长发育, 降低作物产量、品质, 影响收获效率, 造成巨大经济损失[1,2]。近年, 由于免耕、直播、抛秧、旱育秧等栽培技术的推广, 使稻田杂草危害加剧[3]。化学除草剂作为一种快速、有效、经济、实用的除草措施, 为全球农作物高产做出了巨大贡献。而除草剂对常规作物同样具有杀害性, 一定程度上限制了其使用范围, 培育和利用抗除草剂作物是推动化学除草剂发展的动力, 除草剂和抗性作物的配合使用是目前最为经济有效解决稻田杂草问题的方式。
培育抗除草剂作物的方法主要包括自然选择、人工诱变、杂交育种及转基因育种等。目前, 在抗不同种类除草剂的作物中, 只有抗草甘膦、草铵膦及抗咪唑啉酮类除草剂作物获得大面积的种植[4], 最受人们青睐的为用非转基因手段获得的抗咪唑啉酮类除草剂作物, 商业化的该类抗性作物主要有水稻、玉米、油菜、小麦和向日葵[5]。咪唑啉酮类除草剂由美国氰胺公司开发, 已有6个商业化品种, 包括咪唑烟酯、咪唑乙烟酸、咪唑喹啉酸、甲氧咪草烟、甲基咪草烟和咪草酸。这是一类广谱性除草剂, 通过抑制靶标酶——乙酰乳酸合酶(acetolactate synthase, ALS), 影响植物体内支链氨基酸合成, 进而影响植物生长, 具有活性高、杀草谱广、用量少、对哺乳动物低毒等特点[6]。咪草烟是咪唑乙烟酸家族成员, 可播前混土使用, 也可苗前、出苗后及插秧后使用, 适用于水稻整个生育期[7]。抗咪草烟水稻品种主要包括XL729、XL745、XP756和XP534[8], 美国通过推广抗咪草烟水稻获得巨大收益, 至2011年抗咪草烟水稻及其杂交稻已占美国水稻的50%[8], 该类水稻品种应用潜力巨大。
分子标记辅助选择与传统表型选择相比能精确跟踪目标性状, 减少育种的盲目性, 选择过程不受环境及人为因素影响; 减少田间工作, 节约育种成本; 且可在作物早世代进行目标性状的选择, 节约时间, 缩短育种周期[9]。分子标记辅助育种与传统育种手段结合在各作物的抗病、优质、高产等选育中发挥着重要作用, 抗除草剂作为一种优良性状, 越来越多的研究者着眼于筛选相关分子标记。胡茂龙等[10]开发了AP15F/AP18R、AP15F/AP19R标记用于抗咪唑啉酮类除草剂油菜的育种; 李志江等[11]筛选了水稻抗除草剂“拿捕净”的SSR标记并选育了抗除草剂材料; Bulos等[12]开发了向日葵抗除草剂的分子标记; Mithila等[13]筛选了芥子抗植物生长素类除草剂的分子标记, 这些标记为作物抗除草剂育种提供了便利。
本研究对筛选得到的抗咪草烟水稻品种金粳818进行遗传分析, 利用获得的连锁标记进行分子标记辅助育种, 并验证其在长江中下游常规粳稻中的应用潜力。该研究为粳稻抗除草剂分子标记辅助选择育种奠定了基础。
1 材料与方法
1.1 试验材料
金粳818、苏垦118、南粳9108及91份粳稻(表1)均为长江中下游主要推广的粳稻品种, 具有高产、稳产、优质等显著特点, 其中, 金粳818是本团队鉴定的具有抗咪草烟特性的粳稻品种。Table 1
表1
表1长江中下游主要推广的91份粳稻品种
Table 1Ninety-one commercial japonica rice varieties in the middle and lower reaches of Yangtze River
| 生育特性 Maturing type | 品种 Cultivar |
|---|---|
| 中熟中粳 Medium-maturing medium japonica rice | 连粳4号, 连粳7号, 连粳9号, 宁粳4号, 徐稻3号, 镇稻88, 镇稻99, 连粳10号, 连粳11, 华粳5号, 华粳6号, 淮稻8号, 淮稻11, 淮优粳2号, 盐稻11, 盐粳11, 武运粳21, 武运粳27, 华瑞稻1号, 淮糯11, 淮粳096, 扬中稻1号, 扬辐粳1号, 扬粳4308, 泗稻12 Lianjing 4, Lianjing 7, Lianjing 9, Ningjing 4, Xudao 3, Zhendao 88, Zhendao 99, Lianjing 10, Lianjing 11, Huajing 5, Huajing 6, Huaidao 8, Huaidao 11, Huaiyoujing 2, Yandao 11, Yanjing 11, Wuyunjing 21, Wuyunjing 27, Huaruidao 1, Huainuo 11, Huaijing 096, Yangzhongjing 1, Yangfujing 1, Yangjing 4308, Sidao 12 |
| 迟熟中粳 Late-maturing medium japonica rice | 淮稻5号, 盐稻8号, 盐稻9号, 南粳49, 南粳40, 南粳41, 南粳45, 扬育粳2号, 扬辐粳8号, 苏沪香粳, 武陵粳1号, 武育粳3号, 武运粳24, 华粳3号, 华粳4号, 华粳7号, 淮稻7号, 淮稻9号, 淮稻10号, 淮稻13, 盐粳9号, 盐粳10号, 宁粳5号, 武运粳4号, 武运粳11 Huaidao 5, Yandao 8, Yandao 9, Nanjing 49, Nanjing 40, Nanjing 41, Nanjing 45, Yangyujing 2, Yangfujing 8, Suluxiangjing, Wulingjing 1, Wuyunjing 3, Wuyunjing 24, Huajing 3, Huajing 4, Huajing 7, Huaidao 7, Huaidao 9, Huaidao 10, Huaidao 13, Yanjing 9, Yanjing 10, Ningjing 5, Wuyunjing 4, Wuyunjing 11 |
| 早熟晚粳 Early-maturing late japonica rice | 宁粳1号, 南粳5055, 南粳44, 武香粳14, 南粳47, 武运粳7号, 镇稻18, 淮香稻15, 宁粳3号, 南粳42, 通粳981, 武粳13, 武粳15, 武香粳9号, 武运粳19, 武运粳23, 武运粳29, 常农粳4号, 常农粳5号, 常农粳7号, 镇稻7号, 镇稻10号, 镇稻15, 镇稻16, 镇糯19, 宁粳2号, 常粳144, 苏粳5号, 镇稻1号, 镇稻9424 Ningjing 1, Nanjing 5055, Nanjing 44, Wuxiangjing 14, Nanjing 47, Wuyunjing 7, Zhendao 18, Huaixiangdao 15, Ningjing 3, Nanjing 42, Tongjing 981, Wujing 13, Wujing 15, Wuxiangjing 9, Wuyunjing 19, Wuyunjing 23, Wuyunjing 29, Changnongjing 4, Changnongjing 5, Changnongjing 7, Zhendao 7, Zhendao 10, Zhendao 15, Zhendao 16, Zhennuo 19, Ningjing 2, Changjing 144, Sujing 5, Zhendao 1, Zhendao 9424 |
| 中熟晚粳 Medium-maturing late japonica rice | 南粳46, 扬粳4227, 武育粳18, 常农粳3号, 常农粳6号, 苏粳8号, 苏香粳2号, 镇稻12, 镇稻13, 镇稻17, 苏粳9号 Nanjing 46, Yangjing 4227, Wuyujing 18, Changnongjing 3, Changnongjing 6, Sujing 8, Suxiangjing 2, Zhendao 12, Zhendao 13, Zhendao 17, Sujing 9 |
新窗口打开
1.2 表型鉴定
于江苏省农业科学院试验基地种植所有材料, 采用完全随机区组设计, 每行10株, 株行距为18 cm× 22 cm, 以常规方法栽培。水稻种子催芽后播种, 在幼苗长至二叶一心时调查群体抗性, 利用咪草烟(70 g ai hm-2)喷洒筛选, 2周后调查表型, 将正常生长的幼苗记为抗咪草烟的个体, 将死亡的幼苗记为对咪草烟敏感的个体。1.3 遗传分析
以金粳818与苏垦118杂交获得的F2群体作为遗传分析群体, 采用分群法定位基因[14]。根据F2群体的表型, 分别随机选取死亡与存活单株各5株, 取等量叶片, 构建抗性基因池与感性基因池, 以CTAB法提取DNA[15]。参照Gramene数据库(http://www.gramene.org/)中的SSR引物序列, 随机选择500对均匀分布于水稻12对染色体上的SSR分子标记, 筛选多态性标记。PCR体系(20 μL)含2×Taq PCR Green Mix, 10 μL; SSR标记引物对(2 μmol L-1), 各2 μL; 模板DNA, 2 μL; dH2O, 4 μL。PCR条件为预变性94℃, 5 min; 变性94℃, 30 s; 退火60~50℃ (温度与引物相关), 30 s; 延伸72℃, 30 s; 30个循环; 延伸72℃, 5 min; 保温15℃。PCR产物经非变性聚丙烯酰胺凝胶电泳, 快速银染后观察[16]。利用多态性标记随机检测F2群体中的100株敏感单株, 构建遗传图谱。查询定位区间内所有基因信息, 筛选候选基因, 以日本晴的DNA序列设计引物, 扩增候选基因并测序, 分析测序结果。1.4 连锁标记在育种中的应用
以金粳818为供体, 南粳9108为受体, 配制杂交组合, 杂交种自交得到F2群体, 利用1.3中连锁标记对F2群体单株进行鉴定并选择。对选择的植株连续自交并进行表型及标记筛选, 在F7获得稳定材料。随机选择F7群体中若干单株进行标记及表型鉴定, 确定连锁标记在抗咪草烟辅助育种中的可行性。进一步利用连锁标记对91份常规粳稻品种(表1)及金粳818进行多态性分析。若存在多态性, 则标记可应用于辅助改良常规粳稻的抗咪草烟性状, 获得抗咪草烟粳稻新品种。2 结果与分析
2.1 表型鉴定
粳稻品种金粳818为抗咪草烟品种, 在幼苗期喷洒除草剂后仍能够正常生长, 叶片无枯萎变黄现象。常规粳稻品种为咪草烟敏感品种, 喷洒除草剂后, 植株停止生长, 叶片变黄并逐渐枯萎死亡(图1)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1抗咪草烟水稻和咪草烟敏感水稻的表型
R: 抗咪草烟水稻金粳818; S: 感咪草烟水稻苏垦118。
-->Fig. 1Phenotype of imazethapyr-resistant and sensitive rice
R: imazethapyr-resistant rice Jinjing 818; S: imazethapyr-sensitive rice Suken 118.
-->
2.2 遗传分析
以金粳818为供体, 苏垦118为受体, 收获F1种子, F1幼苗长至二叶一心时喷洒咪草烟, 结果显示F1植株全部存活, 表明杂种F1抗咪草烟。F2幼苗以相同的方式处理, 580株F2植株中, 430株抗性苗能够正常生长, 150株敏感苗立即枯萎死亡(图2), 抗、感植株符合3∶1理论分离比例(χ2=0.1862< χ20.05,1=3.84)。以上结果说明金粳818抗咪草烟性状由一对显性核基因控制。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2亲本金粳818、苏垦118和F2群体中咪草烟抗性及感性苗表型
R: 抗咪草烟幼苗; S: 感咪草烟幼苗。
-->Fig.2Phenotype of imazethapyr-resistant and sensitive seedlings in Jinjing 818, Suken 118, and F2 population
R: imazethapyr-resistant seedlings; S: imazethapyr-sensitive seedlings.
-->
2.3 基因定位
随机选择500对均匀分布于水稻12对染色体上的SSR分子标记, 对金粳818、苏垦118、抗、感DNA混合池进行多态性分析, 发现位于第2染色体上的SSR标记RM7632、RM7413、RM7426、RM5221在样本中主条带清晰, 多态性明显, 重复性好, 能够清楚分辨不同基因型。进一步利用这4对标记筛选F2群体中100株咪草烟敏感(隐性)单株, 发现重组个体分别为5、1、6和11, 将目标基因定位于RM7413和RM7426之间(图3)。经相关软件预测基因, 在定位区间内存在咪草烟靶基因ALS, 测序分析发现该基因在重要功能位点第1880位碱基处G突变为A, 即第627位氨基酸由丝氨酸(Ser, S)突变为天冬酰胺(Asn, N)(图3)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3SSR标记RM7326、RM7413、RM7426、RM5221在咪草烟敏感植株中的基因型(部分图)
1: 抗性对照金粳818的基因型; 2: 敏感对照苏垦118的基因型; 其余为咪草烟敏感植株中基因型。
-->Fig. 3Genotype of SSR markers RM7326, RM7413, RM7426, and RM5221 in imazethapyr-sensitive seedlings (parts of the figures)
1: the genotype of Jinjing 818; 2: the genotype of Suken 118; others are the genotype of imazethapyr-sensitive seedlings.
-->
2.4 标记在实际育种中的应用
分子标记辅助育种为目前重要的育种手段, 因此本实验拟利用获得的抗咪草烟标记用于实际育种, 验证其利用价值, 鉴于RM7413距离靶基因最近, 因此选其为目的标记。以金粳818为供体, 南粳9108为受体, 杂交组配收获F1, F1自交获得F2群体。选取F2个体50株, 利用标记RM7413(F: 5°-GTCTGG TTGGCAGCTCTCTC-3°; R: 5°-CGACACACATCCA CGCAC-3°)分析单株基因型, 选择与抗性亲本金粳818带型完全一致的单株, 选择的单株连续自交并通过表型及标记选择, 在F7获得稳定材料。为验证标记在抗咪草烟辅助育种中的可行性, 对F7群体进行表型分析, 同时对随机个体进行基因型鉴定, 结果表明F7植株在咪草烟处理下全部存活且生长良好, 利用标记RM7413随机鉴定的10株苗均呈现与抗性亲本金粳818完全一致的带型(图5)。上述结果表明, SSR分子标记RM7413能够辅助选育抗咪草烟粳稻新品种。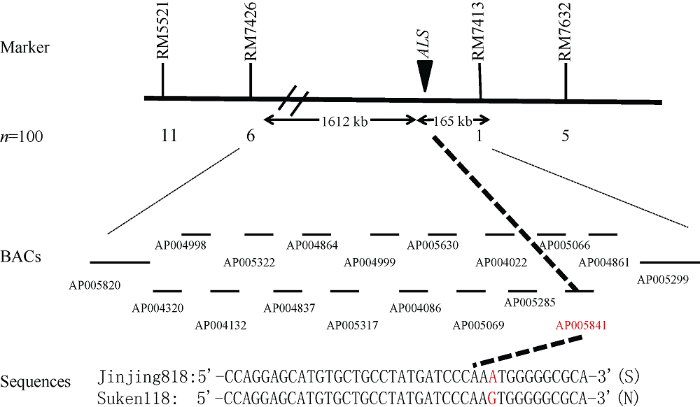 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4抗咪草烟基因的标记连锁图
-->Fig. 4A linkage group of four markers surrounding imazethapyr-resistance gene
-->
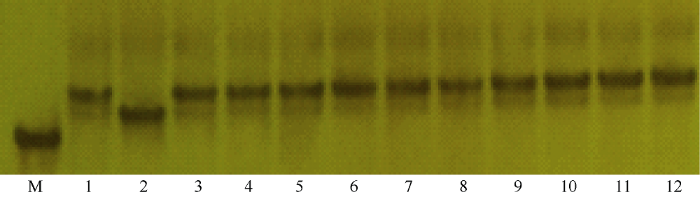 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5SSR标记RM7413在F7的随机10个个体中的基因型
1: 抗性对照金粳818的基因型; 2: 敏感对照南粳9108的基因型; M: DNA marker; 3~12: 10个随机个体的基因型。
-->Fig. 5Genotype of SSR marker RM7413 in ten F7 seedlings
1: the genotype of Jinjing 818; 2: the genotype of Nanjing 9108; M: DNA marker; 3-12: the genotype of ten seedlings.
-->
若标记RM7413在常规粳稻及金粳818中存在多态, 则该标记可用于常规粳稻的除草剂抗性改良。本试验选取91份近年来在长江中下游推广的主要粳稻品种进行多态性鉴定。鉴定结果如图6所示, 91份推广品种均与金粳818存在多态性, 表明分子标记RM7413在长江中下游常规粳稻抗咪草烟改良中具有非常大的应用潜力。
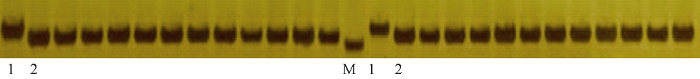 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6SSR标记RM7413在常规粳稻中的基因型(部分图)
1: 抗性对照金粳818的基因型; 2: 敏感对照苏垦118的基因型; M: DNA marker; 其余为常规粳稻的基因型。
-->Fig. 6Genotype of SSR marker RM7413 in conventional japonica rice (parts of the Figures)
1: the genotype of Jinjing 818; 2: the genotype of Suken 118; M: DNA marker; others are the genotype of conventional japonica rice.
-->
3 讨论
长江中下游粳稻生产方式近年来向机械化、轻简化、规模化方向发展, 直播稻面积逐年扩大, 伴随直播的杂草稻危害也日趋严重, 本试验鉴定获得的抗咪草烟粳稻品种金粳818为直播稻田杂草稻防除提供了基因资源, 同时丰富了作物抗除草剂类型, 降低了单一使用草甘膦等除草剂的生态风险[17]。长江中下游地区是我国粳稻主要生产区之一, 然而在由于育种过程中长期依赖于少数核心亲本, 导致江苏地区粳稻间亲缘关系较近, 遗传相似性较高[18,19]; 另有分析表明黑龙江、宁夏地区, 甚至全国范围内的粳稻材料同样存在该现象[20,21,22]; Viviana等[23]对智利的249份粳稻分析表明驯化瓶颈导致粳稻遗传多样性较低。这些研究都表明同一生态区粳稻遗传背景相近, 因此在开展粳-粳杂交品种后代的分子标记辅助选择育种中, 可用的多态性分子标记较少。本试验发现金粳818的咪草烟抗性与RM7413紧密连锁, 该标记与长江中下游主要推广的粳稻品种间存在多态性, 为长江中下游粳稻的抗咪草烟分子育种奠定了基础。
自1996年转基因作物正式商品化以来, 转基因技术被广泛应用于作物育种, 性状涉及抗除草剂、抗虫、抗干旱等, 其中在抗除草剂相关领域最为成功[24], 但转基因作物一直存在较大的争议使其不能够被广泛接受, 同时通过传统育种手段获得的抗除草剂作物极少, 导致中国在抗除草剂作物育种方面存在较大的空白。本试验结果证明粳稻资源金粳818与连锁标记RM7413能够用于抗咪草烟粳稻常规育种, 可极大简化育种过程, 为中国抗除草剂作物育种奠定基础。
4 结论
金粳818抗咪草烟特性是由单基因控制的显性性状, 初步确定乙酰乳酸合酶基因(ALS)的突变是金粳818产生抗性的主要原因。筛选获得的SSR分子标记RM7413与抗性基因紧密连锁, 该标记与金粳818在抗咪草烟粳稻新品种的选育中应用潜力巨大。The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . Saflufenacil is a new herbicide for broadleaf weed control in various crops. In rice, saflufenacil can be a prospective combination partner to complement the broadleaf weed control of herbicides such as imazethapyr. This study was established to evaluate 1) rice tolerance and 2) weed control of red rice ( Oryza sativa L.) and hemp sesbania ( Sesbania exaltata P. Mill.) to saflufenacil tank-mixed with imazethapyr. The experiment was conducted during 2009 and 2010 at the Texas A&M AgriLife Research and Extension Center located at Beaumont, TX. Treatments included a check, an imazethapyr treatment alone (7002g a.i.02ha 611 at the 1- to 2-leaf stage (EPOST) plus 7002g a.i.02ha 611 at the 4- to 6-leaf stage (LPOST)), and four saflufenacil doses (12.5, 18.75, 25, and 5002g a.i.02ha 611) applied at EPOST and LPOST. Imazethapyr treatment was applied to all saflufenacil treatments. Rice injury increased with doses of saflufenacil and injury up to 83% was observed at 7 DAA when 5002g02ha 611 was applied EPOST. Subsequent evaluations indicated less injury over time demonstrating rice recovery from saflufenacil treatments. No injury was observed in the imazethapyr treatment alone. Rice plants seemed to recover faster from LPOST application injury than EPOST. Hemp sesbania control was ≥88% in all saflufenacil treatments in evaluations conducted before harvest indicating effective control throughout the growing season. Red rice control was 100% for all treated plots containing imazethapyr in both years. Therefore, imazethapyr control of red rice was not adversely affected by tank-mixing with saflufenacil. Although injury was significantly higher in the highest doses of saflufenacil, rice yield was not adversely altered by the herbicide treatments. In summary, rice was injured at the highest doses of saflufenacil, but injury did not reduce rice yield. Hemp sesbania was effectively controlled by saflufenacil (>90% on average). No antagonistic interactions between saflufenacil and imazethapyr were observed with red rice control. Saflufenacil appears to be an effective herbicide candidate for broadleaf control in rice. |
| [2] | . Research was conducted in 2007 and 2008 to evaluate weed-control options in an imazethapyr-resistant rice production system. Raised beds were formed, and imidazolinone-resistant hybrid rice 'CL 730' was drill-seeded on beds. Five herbicide programs applied up to the four- to six-leaf stage of rice were evaluated with and without additional "as-needed" herbicide at later stages. All the herbicide combinations and as-needed herbicides tested in this research were labeled for rice, and only minor transient injury (< 5%) was initially observed. Weeds emerged throughout the growing season, and as-needed herbicides were applied after the four- to six-leaf stage of rice to control these late-emerging weeds and weeds not effectively controlled with earlier applications, primarily Palmer amaranth. Most of the Palmer amaranth at this site was insensitive to imazethapyr (possibly acetolactate synthase resistant). Therefore, application of as-needed herbicides with different modes of action, such as 2,4-D, were used to improve Palmer amaranth control. Rice yields were often numerically higher in plots that received additional herbicide after the six-leaf stage of rice, but yields were not significantly improved. Se llevó al cabo una investigación en 2007 y 2008 para evaluar las opciones de control de maleza en un sistema de producción de arroz resistente al imazethapyr. Se hicieron camas elevadas y en ellas se sembró a chorro corrido la semilla de arroz híbrido CL 730, resistente a las imidazolinonas. Se evaluaron cinco programas de herbicida aplicados al arroz hasta la etapa de 4 a 6 hojas con y sin herbicida adicional según la necesidad en las etapas posteriores. Todas las combinaciones de herbicida y de herbicidas adicionales según la necesidad que se evaluaron en esta investigación fueron etiquetados para usar en el arroz y solamente se observaron inicialmente da09os menores pasajeros (<5%). Las malezas emergieron a lo largo de la estación de crecimiento y se aplicaron herbicidas adicionales después de la etapa de cuatro a seis hojas para controlar las malezas de emergencia tardía, así como también, las que no se controlaron efectivamente con las aplicaciones anteriores, principalmente, Amaranthus palmeri. La mayor parte de dicha maleza en este sitio fue insensible al imazethapyr (posiblemente resistente al acetolactato sintasa). Debido a esto, para mejorar el control de Amaranthus palmeri se aplicaron herbicidas adicionales según la necesidad con diferentes modos de acción, tales como el 2,4-D. Frecuentemente los rendimientos del arroz resultaron ser numéricamente más altos en las parcelas que recibieron herbicida adicional después de la etapa de seis hojas, pero los rendimientos no mejoraron significativamente. |
| [3] | . Shortages of labor, water and the adverse effects of puddling on soil health are forcing farmers to switch rice production systems from puddled transplant to direct seeding. In recent years, resource conservation technologies (RCTs) like zero-tillage (ZT) and bed-planting are being promoted in the rice–wheat cropping systems of South Asia to address these concerns. However, weed control is a serious challenge in non-puddled, direct-seeded ZT/bed-planted rice. In the absence of weed control, rice yields are reduced by 35–100% in direct-seeded/RCTs systems. Herbicide-resistant rice (HR-rice) may facilitate adoption of RCTs by improving weed management options. The three major direct benefits of introducing HR-rice are to: (1) improve control of weeds specifically associated with rice, e.g. weedy Oryza species; (2) substitute currently used herbicides with new ones that are more efficient and that have better environmental profiles; and (3) provide new tools for managing weeds that have already developed resistance to current herbicides. In this paper, we discuss the current issues associated with the rice–wheat systems of India and role of HR-rice in addressing weed management constraints of direct-seeded rice. Direct-seeded ZT/bed-planted rice will save water, labor cost and drudgery involved in transplanting. Other potential benefits include improvement in soil health, timely sowing of wheat, and reduction in greenhouse gases. |
| [4] | . . |
| [5] | . Imidazolinone herbicides, which include imazapyr, imazapic, imazethapyr, imazamox, imazamethabenz and imazaquin, control weeds by inhibiting the enzyme acetohydroxyacid synthase (AHAS), also called acetolactate synthase (ALS). AHAS is a critical enzyme for the biosynthesis of branched-chain amino acids in plants. Several variant AHAS genes conferring imidazolinone tolerance were discovered in plants through mutagenesis and selection, and were used to create imidazolinone-tolerant maize (Zea mays L), wheat (Triticum aestivum L), rice (Oryza sativa L), oilseed rape (Brassica napus L) and sunflower (Helianthus annuus L). These crops were developed using conventional breeding methods and commercialized as Clearfield* crops from 1992 to the present. Imidazolinone herbicides control a broad spectrum of grass and broadleaf weeds in imidazolinone-tolerant crops, including weeds that are closely related to the crop itself and some key parasitic weeds. Imidazolinone-tolerant crops may also prevent rotational crop injury and injury caused by interaction between AHAS-inhibiting herbicides and insecticides. A single target-site mutation in the AHAS gene may confer tolerance to AHAS-inhibiting herbicides, so that it is technically possible to develop the imidazolinone-tolerance trait in many crops. Activities are currently directed toward the continued improvement of imidazolinone tolerance and development of new Clearfield* crops. Management of herbicide-resistant weeds and gene flow from crops to weeds are issues that must be considered with the development of any herbicide-resistant crop. Thus extensive stewardship programs have been developed to address these issues for Clearfield* crops. |
| [6] | . A novel mutant acetolactate synthase gene from rice cells, which confers resistance to ALS-inhibiting herbicides KAWAI Kiyoshi , KAKU Koichiro , IZAWA Norihiko , SHIMIZU Tsutomu , FUKUDA Atsunori , TANAKA Yoshiyuki J. Pestic. Sci. 32(2), 89-98, 2007-05-20 |
| [7] | . A study was established to evaluate weed control and crop response of imidazolinone-tolerant (IMI-tolerant) rice in water-seeded culture with imazethapyr at 70, 105, and 140 g ai/ha at four different soil application timings with or without 70 g/ha imazethapyr postemergence (POST). Application timings included preplant incorporated (PPI), surface application prior to seeding (SURFACE), following seeding (SEED), and at pegging (PEG). The PEG treatments were applied when green leaf tissue had emerged from the seed and the root had begun to extend downward into the soil. Response of barnyardgrass, red rice, and rice injury was not affected by the addition of an imazethapyr POST application. Barnyardgrass control was above 90% at 28 d after treatment (DAT) for both years. At 42 DAT, barnyardgrass control decreased to 88% in 1998, compared with 94% control in 1999. Averaged over years, Indian jointvetch control ranged from 44 to 74% at 28 DAT and 41 to 71% at 42 DAT. Indian jointvetch control was inconsistent and lacked uniformity over all factors. Rice injury increased as application timing was delayed from PPI to PEG in 1998 at both rating dates. In 1999, injury was not observed with imazethapyr at 28 DAT, and 1% injury was observed at 42 DAT with all imazethapyr application timings. |
| [8] | . The ecological risk of transgenic herbicide-resistant rice(Oryza sativa) has been seriously concerned by professional and public. Evaluating the size of ecological risk and the amount of benefit has important reference value in risk precaution, distinguishing right from wrong, weight the advantage and disadvantage, and scientific policy-making. The previous researches emphasised on individual risk analysis and mainly adopted the simulated environment, but there was less on comparison of risk and benefit and deficient on global and integrated analysis. This paper has comprehensively analysed the ecological risk and benefit of transgenic herbicide-resistant rice, and put forward technical approaches to control the ecological risk, that has vital pratical significance in promoting healthy and steady development of transgenic herbicide-resistant rice in China. . The ecological risk of transgenic herbicide-resistant rice(Oryza sativa) has been seriously concerned by professional and public. Evaluating the size of ecological risk and the amount of benefit has important reference value in risk precaution, distinguishing right from wrong, weight the advantage and disadvantage, and scientific policy-making. The previous researches emphasised on individual risk analysis and mainly adopted the simulated environment, but there was less on comparison of risk and benefit and deficient on global and integrated analysis. This paper has comprehensively analysed the ecological risk and benefit of transgenic herbicide-resistant rice, and put forward technical approaches to control the ecological risk, that has vital pratical significance in promoting healthy and steady development of transgenic herbicide-resistant rice in China. |
| [9] | . . |
| [10] | . . |
| [11] | . 为探究谷子抗病育种的理论基础,采用Blast比对的方法,利用已知的谷子抗"拿捕净"除草剂的基因组序列,对谷子抗"拿捕净"基因进行SSR分子连锁标记筛选研究。利用抗"拿捕净"基因的已知序列找到了在谷子基因组中第7和第9条染色体上均存在同源序列,分别在这些同源序列上下游寻找SSR引物,在谷子F2群体中进行SSR标记的筛选,进行共分离分析。结果表明:位于第7染色体上的2个SSR标记SIMS13569和SIMS13512与谷子抗"拿捕净"基因性状共分离,即表现为连锁遗传,该标记可用于分子标记辅助选择,对谷子抗"拿捕净"基因育种具有重要的现实意义;而第9染色体上的序列不具有抗"拿捕净"的作用。 . 为探究谷子抗病育种的理论基础,采用Blast比对的方法,利用已知的谷子抗"拿捕净"除草剂的基因组序列,对谷子抗"拿捕净"基因进行SSR分子连锁标记筛选研究。利用抗"拿捕净"基因的已知序列找到了在谷子基因组中第7和第9条染色体上均存在同源序列,分别在这些同源序列上下游寻找SSR引物,在谷子F2群体中进行SSR标记的筛选,进行共分离分析。结果表明:位于第7染色体上的2个SSR标记SIMS13569和SIMS13512与谷子抗"拿捕净"基因性状共分离,即表现为连锁遗传,该标记可用于分子标记辅助选择,对谷子抗"拿捕净"基因育种具有重要的现实意义;而第9染色体上的序列不具有抗"拿捕净"的作用。 |
| [12] | . IMISUN, SURES, and CLPlus are three herbicide tolerance traits in sunflower (Helianthus annuusL.) which are determined by the expression of different alleles at the same locus, Ahasl1.Introgression of these genes into high yielding sunflower germplasm should be complemented through the use of molecular markers in order to unambiguously identify the different alleles involved in tolerance. The aim of this work was to present a set of allele specific molecular markers for the different herbicide resistant traits of sunflower, which can be used with various technological platforms, and applied to different objectives in a breeding program. The Ahasl1gene sequences from lines carrying different alleles for susceptibility or resistance showed single nucleotide polymorphisms and length variations for a simple sequence repeat. These differences were utilized to develop three types of PCR markers (SSRs, CAPS and SNPs) which allow the precise identification of each allele at the Ahasl1 locus. Usefulness of these markers with respect to the application of phenotypic selection for herbicide tolerance, and the strategic implementation of these techniques for developing inbred lines with herbicide resistance is discussed. |
| [13] | . |
| [14] | . |
| [15] | . |
| [16] | . A dynamically rolled leaf mutant (rl10) was identified from a spontaneous mutation in an Oryza sativa L. subsp. indica line, II-32B. The leaf chlorophyll content of rl10 is higher than that of the wild type. Genetic analysis using 3 F2 segregating populations derived from crosses between rl10 and the rice lines Mian5B, II-32B, and D62B, respectively, confirmed that the rolled leaf trait of rl10 is controlled by a single recessive gene. Of 719 SSR primer pairs that showed polymorphism between D62B and rl10, 151 were adopted to map the RL10(t) gene using an F2 segregating population of the cross rl10000201— D62B, which contained 352 recessive plants. RL10(t) was primarily mapped on the long arm of chromosome 9, 5.09 cM from marker RM105 and 5.13 cM from marker RM3912. Using a novel set of 22 primer pairs between RM105 and RM3912, RL10(t) was further mapped between markers rlc3 (0.72 cM in distance) and rlc12 (0.1 cM in distance) using an F2/F3 population containing 1172 recessive individuals. Mapped position analysis and homology analysis of the 20 genes within the 194-kb region between these 2 markers both indicated that a gene encoding a Myb-like domain transcription factor with homology to Arabidopsis KANADI (annotated in PAC clone AP005904) is the most probable candidate for RL10(t). This study enables further investigation of whether KANADI-like Myb genes are involved in leaf polarity modeling in monocots, as they are in dicots. Un mutant 0102 enroulement foliaire actif (rl10) a 0108t0108 identifi0108 suite 0102 une mutation spontan0108e chez la lign0108e II-32B de l0964Oryzasativa L. subsp. indica. La teneur en chlorophylle foliaire chez rl10 est plus 0108lev0108e que chez le type sauvage. Une analyse g0108n0108tique, effectu0108e sur trois populations F2 d0108riv0108es de croisement entre rl10 et Mian5B, II-32B et D62B, a confirm0108 que l0964enroulement foliaire actif pr0108sente une h0108r0108dit0108 monog0108nique r0108cessive. Les 151 microsatellites (parmi 719) qui 0108taient polymorphes entre D62B et rl10 ont 0108t0108 employ0108s pour situer le g01¨ne RL10(t) au sein d0964une population F2rl10 01— D62B comptant 352 individus r0108cessifs. RL10(t) a d0964abord 0108t0108 situ0108 sur le bras long du chromosome 9, 0102 5,09 cM de RM105 et 0102 5,13 cM de RM3912. 01 l0964aide d0964un nouveau jeu de 22 microsatellites situ0108s entre RM105 et RM3912, RL10(t) a 0108t0108 positionn0108 entre rlc3 (0102 0,72 cM) et rlc12 (0102 0,1 cM) en utilisant une population F2/F3 comprenant 1172 individus r0108cessifs. L0964analyse de la position cartographique et de l0964homologie des 20 g01¨nes situ0108s dans l0964intervalle de 194 kb qui s0108pare ces deux marqueurs sugg01¨rent tous deux qu0964un g01¨ne codant pour un facteur de transcription 0102 domaine Myb et pr0108sentant une homologie avec le g01¨ne KANADI (annot0108 dans le clone PAC AP005904) serait le candidat le plus probable pour RL10(t). Cela permettra d0964examiner si des g01¨nes Myb ressemblant 0102 KANADI sont 0108galement impliqu0108s dans l09640108tablissement de la polarit0108 foliaire chez les monocotyl0108dones comme chez les dicotyl0108dones. |
| [17] | . |
| [18] | . 利用48对SSR分子标记对2007—2013年江苏省审定的65份不同类型常规粳稻品种和7份对照品种进行遗传多样性分析。结果表明:42对分子标记在供试材料间存在多态性,42对SSR标记共检测到101个等位基因,变化范围为2~4个,平均每个位点2.40个。基因多样性指数变异范围为0.03~0.64,平均0.21。多态信息含量(PIC)变异范围为0.03~0.58,平均0.18。65份江苏省常规粳稻品种间的遗传相似系数变异范围在0.78~0.97之间,平均0.91,95.4%的供试品种其遗传相似系数在0.87~0.97之间。3个不同类型的粳稻品种群体内,迟熟中粳检测出的等位基因数量最高,迟熟中粳的平均位点PIC值和基因多样性指数均高于晚粳品种和中熟中粳。UPGMA聚类结果表明,以遗传相似系数0.87为界,可将除‘通鉴981’、‘扬农稻1号’和‘盐稻10号’以外的62份江苏省常规粳稻品种分为2类;江苏省大面积生产主推粳稻品种的遗传多样性不够丰富,品种间的遗传距离较小,同一育种单位选育的品种间遗传距离更小。 . 利用48对SSR分子标记对2007—2013年江苏省审定的65份不同类型常规粳稻品种和7份对照品种进行遗传多样性分析。结果表明:42对分子标记在供试材料间存在多态性,42对SSR标记共检测到101个等位基因,变化范围为2~4个,平均每个位点2.40个。基因多样性指数变异范围为0.03~0.64,平均0.21。多态信息含量(PIC)变异范围为0.03~0.58,平均0.18。65份江苏省常规粳稻品种间的遗传相似系数变异范围在0.78~0.97之间,平均0.91,95.4%的供试品种其遗传相似系数在0.87~0.97之间。3个不同类型的粳稻品种群体内,迟熟中粳检测出的等位基因数量最高,迟熟中粳的平均位点PIC值和基因多样性指数均高于晚粳品种和中熟中粳。UPGMA聚类结果表明,以遗传相似系数0.87为界,可将除‘通鉴981’、‘扬农稻1号’和‘盐稻10号’以外的62份江苏省常规粳稻品种分为2类;江苏省大面积生产主推粳稻品种的遗传多样性不够丰富,品种间的遗传距离较小,同一育种单位选育的品种间遗传距离更小。 |
| [19] | . . |
| [20] | . Despite extensive studies on cultivated rice, the genetic structure and subdivision of this crop remain unclear at both global and local scales. Using 84 nuclear simple sequence repeat markers, we genotyped a panel of 153 global rice cultivars covering all previously recognized groups and 826 cultivars representing the diversity of Chinese rice germplasm. On the basis of model-based grouping, neighbour-joining tree and principal coordinate analysis, we confirmed the widely accepted five major groups of rice cultivars (indica, aus, aromatic, temperate japonica and tropical japonica), and demonstrated that rayada rice was unique in genealogy and should be treated as a new (the sixth) major group of rice germplasm. With reference to the global classification of rice cultivars, we identified three major groups (indica, temperate japonica and tropical japonica) in Chinese rice germplasm and showed that Chinese temperate japonica contained higher diversity than that of global samples, whereas Chinese indica and tropical japonica maintained slightly lower diversity than that present in the global samples. Particularly, we observed that all seasonal, drought-tolerant and endosperm types occurred within each of three major groups of Chinese cultivars, which does not support previous claims that seasonal differentiation exists in Indica and drought-tolerant differentiation is present in Japonica. It is most likely that differentiation of cultivar types arose multiple times stemming from artificial selection for adaptation to local environments. |
| [21] | . 利用SSR分子标记,对来自于 不同选育单位的粳稻品种间的遗传多样性及群体遗传结构进行分析,推断品种间的亲缘关系,为未来育种的亲本选配提供依据。选取均匀分布于水稻12个连锁群的 48对引物,对来自不同选育单位的36份水稻材料进行遗传多样性和遗传结构分析。遗传多样性结果表明:48对引物中27对具有多态,占引物总数的56%。 共检测出65个等位变异,有多态性的单个引物检测等位位点数为2~5个,平均等位基因数(Na)为2.4个。平均遗传多样性指数(He)为0.307,变 幅为0.050~0.662。平均多态性信息含量(PIC)为0.263,变幅为0.053~0.588,平均Shannon-weiner指数为 0.514,变幅为0.127~1.168。供试品种间的遗传相似系数介于0.38~1.00之间,平均为0.77。遗传相似系数在0.71~0.86占 总数的68.4%;0.71以下占总数的19%;0.86以上占总数的12.5%。并绘制聚类树状图。基于Structure模型的遗传结构分析把供试材 料分为5个遗传结构群:群Ⅰ、群Ⅱ、群Ⅲ、群Ⅳ和群Ⅴ,类群间的遗传一致度变幅为0.7642(群Ⅰ和群Ⅳ)~0.9303(群Ⅲ和群Ⅴ),遗传距离变幅 为0.0719(群Ⅲ和群Ⅴ)~0.269(群Ⅰ和群Ⅳ)。群组间聚类将5个类群分为3大类,群Ⅲ和群Ⅴ以及群Ⅱ聚为一大类,群Ⅲ和群Ⅴ首先聚为一类,群 Ⅰ与群Ⅳ各自成为一类。可见,同一单位选育的品种,亲缘关系较近,材料间总体遗传基础狭窄。建议黑龙江水稻育种者在未来亲本选配时尽量选择其他单位或地区 育成的品种,以丰富选育品种的遗传多样性。 . 利用SSR分子标记,对来自于 不同选育单位的粳稻品种间的遗传多样性及群体遗传结构进行分析,推断品种间的亲缘关系,为未来育种的亲本选配提供依据。选取均匀分布于水稻12个连锁群的 48对引物,对来自不同选育单位的36份水稻材料进行遗传多样性和遗传结构分析。遗传多样性结果表明:48对引物中27对具有多态,占引物总数的56%。 共检测出65个等位变异,有多态性的单个引物检测等位位点数为2~5个,平均等位基因数(Na)为2.4个。平均遗传多样性指数(He)为0.307,变 幅为0.050~0.662。平均多态性信息含量(PIC)为0.263,变幅为0.053~0.588,平均Shannon-weiner指数为 0.514,变幅为0.127~1.168。供试品种间的遗传相似系数介于0.38~1.00之间,平均为0.77。遗传相似系数在0.71~0.86占 总数的68.4%;0.71以下占总数的19%;0.86以上占总数的12.5%。并绘制聚类树状图。基于Structure模型的遗传结构分析把供试材 料分为5个遗传结构群:群Ⅰ、群Ⅱ、群Ⅲ、群Ⅳ和群Ⅴ,类群间的遗传一致度变幅为0.7642(群Ⅰ和群Ⅳ)~0.9303(群Ⅲ和群Ⅴ),遗传距离变幅 为0.0719(群Ⅲ和群Ⅴ)~0.269(群Ⅰ和群Ⅳ)。群组间聚类将5个类群分为3大类,群Ⅲ和群Ⅴ以及群Ⅱ聚为一大类,群Ⅲ和群Ⅴ首先聚为一类,群 Ⅰ与群Ⅳ各自成为一类。可见,同一单位选育的品种,亲缘关系较近,材料间总体遗传基础狭窄。建议黑龙江水稻育种者在未来亲本选配时尽量选择其他单位或地区 育成的品种,以丰富选育品种的遗传多样性。 |
| [22] | . 选用分布于水稻12条染色体上的47对引物对宁夏89份粳稻种质材料进行SSR分析,以探讨宁夏粳稻品种的遗传多样性水平.结果表明,(1)47个位点上 共检测到204个具有多态性的等位片段,每对引物检测出2~9个多态性片段,平均为4.16个;(2)聚类分析显示,89份材料的相似系数为 0.63~0.91,平均为0.79.在相似系数0.718处聚为8个类群.研究表明,宁夏粳稻种质之间的相似性较高,遗传差异较小,遗传背景比较单一. . 选用分布于水稻12条染色体上的47对引物对宁夏89份粳稻种质材料进行SSR分析,以探讨宁夏粳稻品种的遗传多样性水平.结果表明,(1)47个位点上 共检测到204个具有多态性的等位片段,每对引物检测出2~9个多态性片段,平均为4.16个;(2)聚类分析显示,89份材料的相似系数为 0.63~0.91,平均为0.79.在相似系数0.718处聚为8个类群.研究表明,宁夏粳稻种质之间的相似性较高,遗传差异较小,遗传背景比较单一. |
| [23] | . |
| [24] | . 61Herbicide use is increasing due to glyphosate-resistant weeds selected in GMHR crops.61Cultivars with stacked-HR traits are required in the short-term for their management.61Robust crop stewardship plans need to be developed and monitored for compliance.61The only sustainable path is herbicide-use reduction achieved by new or emerging tools. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}