张宏军1,, 宿振起2,3, 柏贵华3,4, 张旭5, 马鸿翔5, 李腾1, 邓云6, 买春艳7, 于立强8, 刘宏伟1, 杨丽1, 李洪杰1,*,, 周阳1,*, 1 中国农业科学院作物科学研究所 / 农作物基因资源与基因改良国家重大科学工程, 北京1000812 河北省农林科学院粮油作物研究所, 河北石家庄0500353 Department of Agronomy, Kansas State University, Manhattan, KS 66506, USA4 USDA-ARS / Hard Winter Wheat Genetics Research Unit, Manhattan, KS 66506, USA5 江苏省农业科学院 / 江苏省农业生物学重点实验室, 江苏南京2100146 福建省南平市农业科学研究所, 福建建阳3542007 新乡矮败小麦育种技术创新中心, 河南新乡4537318 石家庄市农林科学研究院赵县试验基地, 河北赵县051530
Improvement of Resistance of Wheat Cultivars to Fusarium Head Blight in the Yellow-Huai Rivers Valley Winter Wheat Zone with Functional Marker Selection of Fhb1 Gene
ZHANGHong-Jun1,, SUZhen-Qi2,3, BAIGui-Hua3,4, ZHANGXu5, MAHong-Xiang5, LITeng1, DENGYun6, MAIChun-Yan7, YULi-Qiang8, LIUHong-Wei1, YANGLi1, LIHong-Jie1,*,, ZHOUYang1,*, 1 National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China2 Institute of Cereal and Oil Crops, Hebei Academy of Agriculture and Forestry Sciences, Shijiazhuang 050035, Hebei, China3 Department of Agronomy, Kansas State University, Manhattan, KS 66506, USA4 USDA-ARS / Hard Winter Wheat Genetics Research Unit, Manhattan, KS 66506, USA5 Jiangsu Academy of Agricultural Sciences / Jiangsu Provincial Key Laboratory of Agrobiology, Nanjing 210014, Jiangsu, China6 Nanping Institute of Agricultural Sciences of Fujian Province, Jianyang 354200, Fujian, China7 Xinxiang Innovation Center for Breeding Technology of Dwarf-Male-Sterile Wheat, Xinxiang 453731, Henan, China8 Zhaoxian Experiment Station, Shijiazhuang Academy of Agriculture and Forestry Sciences, Zhaoxian 515300, Hebei, China 通讯作者:* 通信作者(Corresponding authors): 李洪杰, E-mail: lihongjie@caas.cn; 周阳, E-mail: zhouyang@caas.cn* 通信作者(Corresponding authors): 李洪杰, E-mail: lihongjie@caas.cn; 周阳, E-mail: zhouyang@caas.cn* 通信作者(Corresponding authors): 李洪杰, E-mail: lihongjie@caas.cn; 周阳, E-mail: zhouyang@caas.cn 收稿日期:2017-07-19 接受日期:2018-01-8 网络出版日期:2018-01-29 版权声明:2018作物学报编辑部作物学报编辑部 基金资助:本研究由国家重点研发计划项目(2016YFD0101802, 2017YFD0100600)和作物分子育种国家工程实验室项目资助 作者简介: -->zhanghongjun01@caas.cn
关键词:小麦;赤霉病抗性;Fhb1基因;分子标记辅助选择 Abstract Fusarium head blight (FHB) becomes a major disease in the wheat production of the Yellow-Huai Rivers Valley Winter Wheat Zone of China. Improving FHB resistance is one of the major breeding targets in this region. The donors of Fhb1 gene, including Ningmai 9, Shengxuan 6, Jianyang 798, Jianyang 84, Sumai 3, and Ningmai 13, were moderately or highly resistant to FHB. Six backcrossing populations were developed by crossing these donors with a dwarfing and male-sterile wheat near isogenic line Zhoumai 16, and backcrossing with Zhoumai 16. The progenies were subjected to phenotypic evaluation of FHB by both the floret-inoculation method and natural infection in a field disease nursery. Fhb1 was detected by a functional Kompetitive Allele Specific PCR (KASP) marker. The backcrossing progenies with Fhb1 gene were moderately susceptible to FHB. Compared with the backcrossing progenies without Fhb1 gene, mean number of diseased spikelets and disease index reduced by 4.2 (P < 0.01) and 4.0 in the progenies with Fhb1 gene, respectively. The mean number of diseased spikelets and disease index for the backcrossing progenies with Fhb1 gene were 8.1 (P < 0.01) and 28.4 (P < 0.01) lower than those of the recurrent parent Zhoumai 16. There were significant differences in resistance among the backcrossing progenies from different donors. The progenies from Shengxuan 6 had better performance in number of diseased spikelets and disease index compared with those from any other donors. The results from this study indicate that Fhb1 gene can efficiently improve the FHB resistance of wheat cultivars grown in the Yellow-Huai Rivers Valley Winter Wheat Zone.
Keywords:Triticum aestivum L.;Fusarium head blight resistance;Fhb1 gene;molecular marker-assisted selection -->0 PDF (3899KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 张宏军, 宿振起, 柏贵华, 张旭, 马鸿翔, 李腾, 邓云, 买春艳, 于立强, 刘宏伟, 杨丽, 李洪杰, 周阳. 利用Fhb1基因功能标记选择提高黄淮冬麦区小麦品种对赤霉病的抗性[J]. 作物学报, 2018, 44(04): 505-511 https://doi.org/10.3724/SP.J.1006.2018.00505 ZHANGHong-Jun, SUZhen-Qi, BAIGui-Hua, ZHANGXu, MAHong-Xiang, LITeng, DENGYun, MAIChun-Yan, YULi-Qiang, LIUHong-Wei, YANGLi, LIHong-Jie, ZHOUYang. Improvement of Resistance of Wheat Cultivars to Fusarium Head Blight in the Yellow-Huai Rivers Valley Winter Wheat Zone with Functional Marker Selection of Fhb1 Gene[J]. Acta Agronomica Sinica, 2018, 44(04): 505-511 https://doi.org/10.3724/SP.J.1006.2018.00505 小麦赤霉病(Fusarium head blight, FHB)是由镰孢属(Fusarium spp.)引起的一种世界性真菌病害, 主要危害麦穗和籽粒, 不仅造成产量损失, 而且其引起的真菌毒素还严重危害人畜健康[1]。受气候变暖以及秸秆还田、免耕栽培等耕作制度改变的影响, 小麦赤霉病发生区域由长江中下游麦区迅速向黄淮冬麦区扩展, 目前已经由次要病害上升为黄淮冬麦区的一种主要病害[2]。在长江中下游麦区, 利用中感或感病品种间杂交, 从抗性超亲分离后代中选育出一批丰产性好、中抗或中感赤霉病的品种[2]。然而, 作为我国小麦主产区的黄淮冬麦区[3], 由于历史上赤霉病不是该麦区小麦的主要病害, 因此抗赤霉病育种没有受到广泛关注, 该麦区培育和推广的绝大多数小麦品种都高感赤霉病。发现和培育抗赤霉病小麦种质资源, 提高小麦生产品种对赤霉病的抗性已经成为黄淮冬麦区主要育种目标之一。 研究发现, 抗赤霉病基因/QTL主要有两类, 分别是抗扩展(Type II)相关基因, 包括Fhb1 [4,5]、Fhb2 [6]、Fhb3 [7]和Fhb7 [8], 以及抗侵入(Type I)相关基因, 包括Fhb4 [9]和Fhb5 [10]。其中, 位于3B染色体上的Fhb1是目前发现效应最大的、抗性最稳定的抗赤霉病基因 [11]。利用Fhb1分子标记辅助选择技术, 已在美国[12,13]、加拿大[14]、澳大利亚[15]、日本[16]等发达国家育成一批小麦新品种, 其赤霉病抗性显著提高。黄淮冬麦区小麦赤霉病的发生年度间变化很大, 在发病轻的年份, 难以在田间条件下对育种材料进行准确的抗病性表型鉴定。稳定的表型鉴定也是黄淮冬麦区小麦抗赤霉病育种遇到的难题, 分子标记辅助选择不仅有助于摆脱表型鉴定难的困境, 而且还可以加速育种进程。本研究利用黄淮冬麦区高感赤霉病小麦品种周麦16的矮败小麦近等基因系(以下简称矮败周麦16)做母本, 与6个携带Fhb1基因供体亲本杂交和回交, 构建了6个回交群体, 目的是分析黄淮冬麦区小麦品种遗传背景下Fhb1基因对赤霉病抗性的影响, 同时比较同一遗传背景下来自不同供体的Fhb1基因对赤霉病的抗性, 为分子标记辅助育种技术在黄淮冬麦区小麦抗赤霉病品种的培育提供实验依据和技术支持。
为了比较同一供体Fhb1基因对赤霉病抗性的影响, 以及在同一遗传背景下来自不同供体的Fhb1基因对赤霉病抗性的影响, 分别从上述相应的6个回交群体中选取11株携带Fhb1基因的可育株和11株不携带Fhb1基因的可育株, 按株系种植, 进行赤霉病抗性对比分析, 同时对携带Fhb1基因的可育株与轮回亲本周麦16、中感赤霉病对照淮麦20、高感赤霉病对照安农8455及中抗对照扬麦158的赤霉病抗性进行比较。人工接种鉴定的发病情况比较充分(图1), 来自6个供体携带Fhb1基因的11个株系的平均病小穗数和平均病情指数分别比轮回亲本周麦16小8.1 (P < 0.01)和28.4 (P < 0.01); 与不携带Fhb1的11个株系相比, 平均病小穗数和平均病情指数分别少4.8 (P < 0.01) 和4.0 (图2)。整体而言, 11个携带Fhb1基因株系的赤霉病抗性达到中感对照淮麦20水平。单花滴注接种鉴定结果表明, 除建阳798外的5个供体回交群体中携带Fhb1基因的株系, 病小穗数均显著低于不含有Fhb1基因的株系(P < 0.05或P < 0.01)。5个供体回交群体的田间病圃鉴定结果与人工接种鉴定结果趋势一致, 仅宁麦13表现不一致。其中, 携带宁麦9号和生选6号Fhb1基因的回交后代株系病情指数显著低于不携带Fhb1的回交后代株系(P < 0.05)。 显示原图|下载原图ZIP|生成PPT 图1单花滴注接种鉴定携带和不携带供体Fhb1基因的回交后代、轮回亲本及对照赤霉病抗性表现 携带宁麦9号(A)、生选6号(B)、建阳798 (C)、建阳84 (D)、苏麦3号(E)和宁麦13 (F) Fhb1基因回交后代的病小穗数明显少于不携带供体Fhb1基因的回交后代。轮回亲本周麦16较中感对照淮麦20 (G)、高感对照安农8455较中抗对照扬麦158 (H)具有更多的病小穗数。+: 携带Fhb1基因; –: 不携带Fhb1基因; RP: 轮回亲本; MS: 中感; HS: 高感; MR: 中抗。 -->Fig. 1Performance of FHB resistance in the backcrossing progenies with or without Fhb1 gene from different donors, recurrent parent and controls using the floret-inoculation method The number of diseased spikelets of backcrossing progenies with Fhb1 gene from donors Ningmai 9 (NM 9, A), Shengxuan 6 (SX 6, B), Jianyang 798 (JY 798, C), Jianyang 84 (JY 84, D), Sumai 3 (SM 3, E), and Ningmai 13 (NM 13, F) were obviously lower than the backcrossing progenies without Fhb1 gene. The recurrent parent Zhoumai 16 (ZM 16) and the highly susceptible control Annong 8455 (AN 8455) had more number of diseased spikelets than Huaimai 20 (HM 20), the moderately susceptible control (G) and Yangmai 158 (YM 158), the moderately resistant control (H). +: Fhb1 genotype; –: non-Fhb1 genotype; RP: recurrent parent; MS: moderately susceptible; HS: highly susceptible; MR: moderately resistant. -->
显示原图|下载原图ZIP|生成PPT 图2携带不同供体Fhb1基因回交后代与轮回亲本、中感对照及不携带Fhb1基因回交后代间病小穗数和病情指数比较 携带Fhb1基因的所有回交后代与轮回亲本周麦16 (A)、中感对照淮麦20 (B)和不携带Fhb1基因的所有回交后代(C)病小穗数和病情指数比较。携带宁麦9号Fhb1基因的回交后代(D)、携带生选6号Fhb1基因的回交后代(E)、携带建阳798 Fhb1基因的回交后代(F)、携带建阳84 Fhb1基因的回交后代(G)、携带苏麦3号Fhb1基因的回交后代(H)和携带宁麦13 Fhb1基因的回交后代(I)分别与其不携带Fhb1基因的回交后代间病小穗数和病情指数比较。NDS: 病小穗数(人工接种); DI: 病情指数(自然发病)。++: 携带Fhb1基因的所有回交后代; RP: 轮回亲本; MS: 中感对照; ––: 不携带Fhb1基因的所有回交后代; +: 携带Fhb1基因的回交后代; –: 不携带Fhb1基因的回交后代。误差线上*和**分别表示在P < 0.05和P < 0.01水平下差异显著。 -->Fig. 2Comparison of number of diseased spikelets and disease index between backcrossing progenies with Fhb1 gene from different donors and recurrent parent, moderately susceptible control and the backcrossing progenies without Fhb1 gene Comparisons of the number of diseased spikelets and disease index were carried out between all Fhb1 backcrossing progenies and the recurrent parent Zhoumai 16 (A), the moderately susceptible control Huaimai 20 (B) and all non-Fhb1 backcrossing progenies (C), and between non-Fhb1 and Fhb1 backcrossing progenies from the donor cultivars Ningmai 9 (D), Shengxuan 6 (E), Jianyang 798 (F), Jianyang 84 (G), Sumai 3 (H) and Ningmai 13 (I). NDS: number of diseased spikelets (floret-inoculation method); DI: disease index (natural infection nursery); ++: all Fhb1 backcrossing progenies; RP: recurrent parent; MS: moderately susceptible control; ––: all Fhb1 backcrossing progenies; +: Fhb1 backcrossing progenies; –: non-Fhb1 backcrossing progenies. * and ** above the error bars indicate significant difference at P < 0.05 and P < 0.01, respectively. -->
2.3 不同供体回交后代赤霉病抗性比较
尽管携带来自于6个供体Fhb1基因的回交后代对赤霉病均表现出一定的抗性, 但是不同供体回交后代在抗病性上表现出显著差异。根据单花滴注接种法鉴定结果, 抗病性由强到弱依次为生选6号、苏麦3号、建阳84、宁麦9号、宁麦13和建阳798的回交后代(表1), 生选6号回交后代对赤霉病抗性接近中抗水平; 根据田间病圃自然发病鉴定结果, 抗病性由强到弱排序为生选6号、建阳798、建阳84、苏麦3号、宁麦9号和宁麦13回交后代。两种鉴定方法的鉴定结果均表明, 生选6号的回交后代抗病性表现最好。 Table 1 表1 表1携带不同供体Fhb1基因的回交后代间病小穗数和病情指数比较 Table 1Comparison of number of diseased spikelets and disease index among the backcrossing progenies with Fhb1 gene from different donors
品种(系)或回交后代 Cultivars (lines) or backcrossing progenies
病小穗数 No. of diseased spikelets
病情指数Disease index
平均值±标准差 Mean±SD
范围 Range
平均值±标准差 Mean±SD
范围 Range
宁麦9号回交后代 Ningmai 9 backcrossing progenies
6.5±3.4 de
1.0-13.0
70.9±6.1 ab
61.8-78.8
生选6号回交后代 Shengxuan 6 backcrossing progenies
3.9±3.2 f
1.0-10.0
56.4±7.5 c
48.8-63.8
建阳798回交后代 Jianyang 798 backcrossing progenies
9.5±4.8 c
1.0-20.0
65.6±10.9 c
52.4-76.4
建阳84回交后代 Jianyang 84 backcrossing progenies
5.6±3.9 e
1.0-12.0
65.9±19.7 ab
34.4-86.1
苏麦3号回交后代 Sumai 3 backcrossing progenies
4.8±4.1 ef
1.0-13.0
70.8±12.3 ab
48.1-84.7
宁麦13回交后代 Ningmai 13 backcrossing progenies
7.4±5.6 d
1.0-20.0
81.5±16.4 ab
63.2-95.0
轮回亲本-周麦16 Recurrent parent-Zhoumai 16
14.4±6.0 b
7.0-20.0
96.1±1.4 a
95.0-97.6
中感对照-淮麦20 MS control-Huaimai 20
6.2±2.2 de
2.0-12.0
68.6±11.3 ab
56.9-81.0
中抗对照-扬麦158 MR control-Yangmai 158
3.2±2.2 f
1.0-10.0
—
—
高感对照-安农8455 HS control-Annong 8455
19.7±1.4 a
12.0-20.0
—
—
标准差后不同字母表示轮回亲本、对照品种和不同Fhb1供体的回交后代间差异显著(P < 0.05)。— 表示未测定。Different letters after deviations indicate significant difference amont recurrent parent, controls, and the backcrossing progenies from different Fhb1 donors (P < 0.05). — : data not available. MR: moderately resistant; MS: moderately susceptible; HS: highly susceptible. 新窗口打开
6个回交群体中携带Fhb1基因的回交后代在平均病小穗数和平均病情指数上均显著低于轮回亲本周麦16和不含有Fhb1基因的回交后代, 赤霉病抗性达到中感水平。不同供体回交后代在抗病性上表现出明显差异, 其中最好的是生选6号。通过Fhb1基因分子标记辅助选择技术改良黄淮冬麦区小麦品种的赤霉病抗性是可行的。 致谢 感谢中国农业科学院作物科学研究所夏先春博士在KASP基因分型方面提供的技术支持。 The authors have declared that no competing interests exist. 作者已声明无竞争性利益关系。
YinY, LiuX, LiB, MaZ.Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F .graminearum collected from wheat in China. Phytopathology, 2009, 99: 487-497 [本文引用: 1]
Cheng SH, ZhangY, Bie TD, Gao DR, Zhang BQ.Damage of wheat Fusarium head blight epidemics and genetic improvement of wheat for scab resistance in China .Jiangsu J Agric Sci, 2012, 28: 938-942 (in Chinese with English abstract) [本文引用: 2]
Liu XY, PeiL, Wei YZ, Zhang ZB, Gao HM, XuP.Agronomic traits variation analysis of Huanghuai dryland winter wheat under temperature change background in China: taking Linfen, Shanxi as an example .Sci Agric Sin, 2015, 48: 1942-1954 (in Chinese with English abstract) [本文引用: 1]
[4]
Anderson JA, Stack RW, LiuS, Waldron BL, Fjeld AD, CoyneC, Moreno-SevillaB, Fetch JM, Song QJ, Cregan PB, Frohberg RC.DNA markers for Fusarium head blight resistance QTLs in two wheat populations .Theor Appl Genet, 2001, 102: 1164-1168 [本文引用: 3]
[5]
Cuthbert PA, Somers DJ, ThomasJ, CloutierS, Brulé-BabelA.Fine mapping Fhb1, a major gene controlling Fusarium head blight resistance in bread wheat(Triticum aestivum L.) .Theor Appl Genet, 2006, 112: 1465-1472 [本文引用: 2]
[6]
Cuthbert PA, Somers DJ, Brulé-BabelA.Mapping of Fhb2 on chromosome 6BS: a gene controlling Fusarium head blight field resistance in bread wheat(Triticum aestivum L.) .Theor Appl Genet, 2007, 114: 429-437 [本文引用: 1]
[7]
Qi LL, Pumphre MO, FriebeB, Chen PD, Gill BS.Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to Fusarium head blight disease of wheat .Theor Appl Genet, 2008, 117: 1155-1166 [本文引用: 1]
[8]
GuoJ, ZhangX, HouY, CaiJ, ShenX, ZhouT, XuH, Ohm HW, WangH, LiA, HanF, WangH, KongL.High-density mapping of the major FHB resistance geneFhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection .Theor Appl Genet, 2015, 128: 2301-2316 [本文引用: 1]
[9]
XueS, LiG, JiaH, XuF, LinF, TangM, WangY, AnX, XuH, ZhangL, KongZ, MaZ.Fine mapping Fhb4, a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.) .Theor Appl Genet, 2010, 121: 147-156 [本文引用: 1]
[10]
XueS, XuF, TangM, ZhouY, LiG, AnX, LinF, XuH, JiaH, ZhangL, KongZ, MaZ.Precise mapping Fhb5, a major QTL conditioning resistance to Fusarium infection in bread wheat ( Triticum aestivum L.) .Theor Appl Genet, 2011, 123: 1055-1063 [本文引用: 1]
Liu YK, Tong HW, Zhu ZW, ChenL, ZouJ, Zhang YQ, Jiao CH, Gao BC.Review on improvement of Fusarium head blight resistance in wheat .J Triticeae Crops, 2016, 36: 51-57 (in Chinese with English abstract) [本文引用: 1]
[12]
Brown-GuediraG, GriffeyC, KolbF, McKendry A, Murphy J, Sanford D. Breeding FHB-resistant soft winter wheat: Progress and prospects .Cereal Res Commun, 2008, 36: 31-35 [本文引用: 2]
[13]
BernardoA, BaiG, YuJ, KolbF, BockusW, DongY.Registration of near-isogenic winter wheat germplasm contrasting in Fhb1 for Fusarium head blight resistance . J Plant Regist, 2013, 8: 106-108 [本文引用: 2]
[14]
Randhawa HS, AsifM, PozniakC, Clarke JM, Graf RJ, Fox SL, Humphreys DG, Knox RE, DePauw R M, Singh A K, Cuthbert R D, Hucl P, Spaner D. Application of molecular markers to wheat breeding in Canada .Plant Breed, 2013, 132: 458-471 [本文引用: 2]
[15]
Xie GQ, Zhang MC, ChakrabortyS, Liu CJ.The effect of 3BS locus of Sumai 3 on Fusarium head blight resistance in Australian wheats .Aust J Exp Agric, 2007, 47: 603-607 [本文引用: 2]
[16]
OdaS.Advances in Wheat Genetics:From Genome to Field . Berlin Heidelberg: Springer-Verlag Press, 2015. pp 311-318 [本文引用: 2]
ZhangX, Ren LJ, Tan XY, Zang YH, Ma HX, Qin JC, Lu WZ.Evaluation method of wheat Fusarium head blight resistance for genetic population based on molecular markers .J Nanjing Univ(Nat Sci), 2005, 41: 125-132 (in Chinese with English abstract) [本文引用: 2]
Ministry of Agriculture of the People’s Republic of China . Rules for Resistance Evaluation of Wheat to Diseases and Insect Pests—Part 4: Rules for Resistance Evaluation of Wheat to Wheat Scab (Fusarium graminearum Schwabe [Teleomorph Gibberella zeae (Schwein) Petch]), 2007. pp 1-6 (in Chinese) [本文引用: 1]
[19]
RasheedA, WenW, GaoF, ZhaiS, JinH, LiuJ, GuoQ, ZhangY, DreisigackerS, XiaX, HeZ.Development and validation of KASP assays for genes underpinning key economic traits in bread wheat .Theor Appl Genet, 2016, 129: 1843-1860 [本文引用: 1]
Lu WZ.Screening and application of molecular markers linked to wheat scab resistance .Jiangsu J Agric Sci, 2011, 27: 243-249 (in Chinese with English abstract) [本文引用: 1]
[23]
LiuS, PumphreyM, GillB, TrickH, ZhangJ, DolezelJ, ChalhoubB, AndersonJ.Toward positional cloning of Fhb1, a major QTL for Fusarium head blight resistance in wheat .Cereal Res Commun, 2008, 36: 195-201 [本文引用: 1]
[24]
Zhou WC, Kolb FL, Bai GH, Domier LL, Yao JB.Effect of individual Sumai 3 chromosomes on resistance to scab spread within spikes and deoxynivalenol accumulation within kernels in wheat . Hereditas, 2002, 137: 81-89 [本文引用: 1]
[25]
Pumphrey MO, BernardoR, Anderson JA.Validating the Fhb1 QTL for Fusarium head blight resistance in near-isogenic wheat lines developed from breeding populations .Crop Sci, 2007, 47: 200-206 [本文引用: 1]
Lu WZ, Ma HX.Breeding of wheat cultivar Shengxuan 6 for high-resistant Fusarium head blight .Jiangsu Agric Sci, 2010, 38(3): 153-154 (in Chinese) [本文引用: 1]
Yao JB, Lu WZ.Analysis of yield potential, yield stability and resistance to Fusarium head blight of new wheat cultivar Shengxuan 6 .J Triticeae Crops, 2012, 32: 793-798 (in Chinese with English abstract) [本文引用: 1]
ZhangX, JiangP, Ye RY, WuL, ZhangY, Ma HX.Evaluation and source tracing of resistance to Fusarium head blight in wheat variety Ningmai 9 and its derivatives .Mol Plant Breed, 2017, 15: 1053-1060 (in Chinese with English abstract) [本文引用: 1]
 , 宿振起
, 宿振起 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT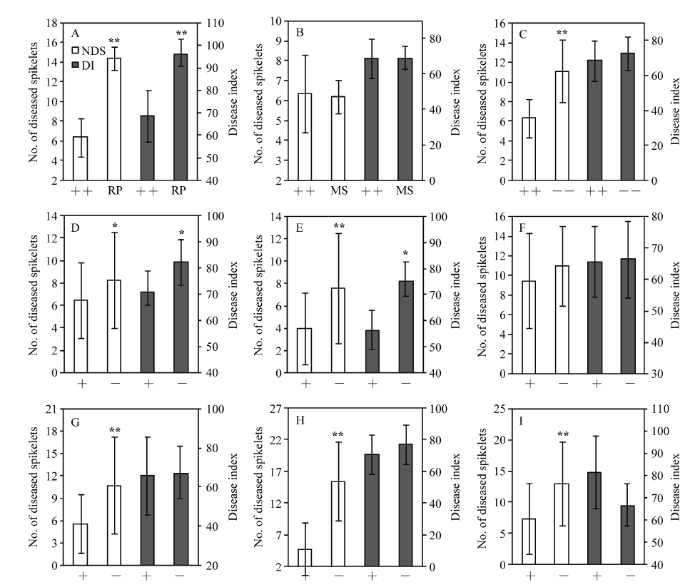 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}