 , ������, ������
, ������, ������Genetic Analysis and Molecular Characterization of a New Allelic Mutant of Vp15 Gene in Maize
WANGRui, ZHANGXiu-Yan, CHENYang-SongͨѶ����:
�ո�����:2017-08-7
��������:2017-11-21
�����������:2018-03-12
��Ȩ����:2018����ѧ���༭������ѧ���༭��
��������:
�����:
-->
չ��
ժҪ
�ؼ��ʣ�
Abstract
Keywords��
-->0
PDF (1593KB)Ԫ������ά��������������ղ�����
�������ø�ʽ����EndNoteRisBibtex�ղر���-->
���������뷢ѿ��ָ���ջ�ǰ������ĸ��ֲ������Ϸ�ѿ������, Ҳ��Ϊ���ȡ��뷢ѿ����Ӱ��С��ˮ��������IJ�����Ʒ�ʺ���������, �����������������صľ�����ʧ, ����뷢ѿ����һֱ���ܹ�����һЩֲ�����ּҺ�����ѧ�ҹ�ע��20����50��������Ͼ��Ѿ���ʼ�о��뷢ѿ����, ������ɸѡ�������뷢ѿ������Դ, �����뷢ѿ����Ҫ�����QTL [1,2,3], ����ȡ���ִ�ʩ��������ϵ��뷢ѿ���⡣
��������, ��Ҫͨ���뷢ѿͻ����(viviparous)�������о��뷢ѿ��ػ���Ŀǰ�ѱ����������뷢ѿ����������, �����뷢ѿͻ������Ҫͨ�����ABA������ϳɻͶ�ABA�ĸ�Ӧ�ȶ�ʹ������ĸ���������ǰ�ȷ�[4,5], ���¿ɽ���Щͻ�����Ϊ�����ࡣ��һ����ABAȱ����, ��ABA�ϳ����衣����ͻ������Ŀǰ�о��Ƚ��������vp2��vp5��vp7��vp8��vp9��vp10��vp12��vp14��vp15��w2��y9��[6,7,8,9,10,11,12,13], ���Dz��������ϳ�ABA, ��������������ABA�����ϵ�, �������������ȷ�, �Ӷ������뷢ѿ������, ����vp5��vp7ͻ����ͨ���������ܲ�������ϳ�;��ø�ĺϳɽ���Ӱ��ABA�ĺϳ�[6,7]; vp12ͻ��������������������ϳ�;����ø�ĺϳ�, �Ӷ�Ӱ��ABA�ĺϳ�[10]; vp14ͻ������ABA����ϳ�;���ĵ�һ���ؼ����豻���, ����������ܲ����ѽ�Ϊ����ȩ�Ĺ��̱����, ����ABA����ϳ�����[9]; vp10ͻ�������⸨���Ӻϳ�����, ʹ��ABAȩ��������ABA[9]���ڶ�����ABA�۸��͡�����ͻ����Ŀǰ�о����������vp1��vp6��[14], ����ͻ���岻Ӱ��ABA������ϳɻ�ABA���³´�л, ֻ�Ǹı��ABA����Ӧ, ���, �����뷢ѿ���Ե���Ӱ�����鷢����������ɫ, ����ԴABA�������������ȷ�������, ����vp1ͻ������ԴABA����������, ֻ�Ƕ���ԴABA�����Խ���, ��������ABA�ź������δ���[15]��
vp15�������뷢ѿͻ����, ������������ֲ�������(MPT)��øС�ǻ�, ��ø�����⸨����(MoCo)����ϳ�[16,17,18,19], ��MoCo�Dz���ABAǰ��——����ܲ����ѽ�ΪABA�����е�ø������ĸ�����[12]��MoCo�㷺������ԭ����������������, ����Ҳ�Ƕ���ø�ĸ�����[20]����ֲ����, ��������4��ø��Ҫ�⸨���Ӳ���ά�ֻ���, ����ȩ����ø������������ø�����ỹԭø������������ø[20], ͻ����vp15����ȱ�������⸨���ӵ���ȩ����ø������������ø������������ø, ���²��������ϳ�ABA, ʹ������ĸ������ǰ�ȷ�[12]�����, vp15ͻ�������о�MPT�ϳ�øС�ǻ����ܵ���Ҫ���ϡ�
���о���ʵ�����·��ֵ�һ�������뷢ѿͻ����vp-like4�������Ŵ������ͻ���λ, ���շ���vp-like4��һ���µ�Vp15�����λͻ���塣���о�Ϊ��һ��������������뷢ѿ�ķ��ӻ��ƺͻ������ṩ�˷ḻ��ʵ����ϡ�
1 �����뷽��
1.1 ʵ�����
�����뷢ѿͻ����(viviparous) vp-like4���ڷۺ�30 d����, ����ڹ����Ϲ۲쵽���Ե��뷢ѿ���͡�����λȺ��Ϊvp-like4�Ӻ������Խ�ϵMo17�ӽ����Խ���õ�F2Ⱥ�塣2��vp15����ͻ����505G (vp15-DR1126)��505I (vp15-umu1)������Maize Genetics Cooperation Stock Center (�����Ŵ�ѧ�����������, https://maizecoop.cropsci.uiuc.edu/request)��1.2 ͻ����ı��ͼ������Ŵ�����
�����뷢ѿ�������ڹ����ջ�ʱ, �ȷ�����ѿ�Ѹɿ�����, ������ѡȡ�뷢ѿ�����ϵ�����������ֲ, �Խ���������뷢ѿ����Ĺ���, Ȼ����ȡ����������ֲ, �Դ���������Խ��۲��ͻ������͡�����ͻ������͵��Ŵ�����, ����ÿһ�����Խ��ڷۺ�30 d����, ������Ҷ����뷢ѿ���, ѡȡ�뷢ѿ����, ����ͳ�������������뷢ѿ�����ķ���ȡ�1.3 Ŀ�Ļ���ij���λ
����BSR-Seq����[21]��Ŀ�Ļ�����г���λ�����ѡȡ���뷢ѿ���ͷ����F2 ����, ��ȡ40�������������뷢ѿ����, �ֱ��ȥ��Ƥ, ���Ϊ2��ʵ����Ʒ, Һ����ĥ, ����ֲ����RNA��ȡ�Լ���(������������Ƽ�����˾, Cat# DP432)��ȡ������RNA, �ڱ�������Ϳ����\������˾����HiSeq2000�������ת¼����ο�BSR-Seq������������[21], ��2�������IJ��������з���, ��ȡ2������������ķ��ӱ��, �����λ������2����ϳصķֲ�����, ������ó�vp-like4����ij���λ���䡣1.4 ��λ��������������λ����
����vp-like4����ij���λ����, ��MaizeGDB���ݿ���վ��(http://www.maizegdb.org/)���������ڵĻ���ע����Ϣ, �����Ѿ������������뷢ѿVp15����λ�ڸ�������[12]��Ϊ���vp-like4��vp15�Ƿ��λ, �ֱ��vp-like4��vp15-umu1��vp15-DR1126���Ӻ������������ѡ����������ֲ, ��ֲvp-like4 3��, vp15-umu1�� vp15-DR1126��2��, ÿ��25�ꡣ�����ڷ�ʱ, ��ѡvp-like4��һ�н��е����Խ�, ͬʱ�ռ���һ�е���Ļ��۽��л��, �ֱ��ڷ��ӽ�vp15-umu1�� vp15-DR1126������һ��ÿһ�����ꡣͬ��, vp15-umu1�� vp15-DR1126������һ��ÿ�������Խ�, ͬʱ�ֱ��ռ����еĻ���, ��۷ֱ��ӽ�vp-like4������2��ÿ�����ꡣ�ڷۺ�30 d, ������Ҷ, ����Ƿ����뷢ѿ�Ĺ��롣
1.5 ������DNA��ȡ��PCR���������з���
�ڷۺ�30 d����, �ֱ�ȡͻ����vp-like4��vp15-umu1��vp15-DR1126�����������뷢ѿ����, ��CTAB����ȡ��������DNA[22]������Vp15����������, ����Primer3����(http://primer3.ut.ee/)�������7������(VP15-G-F1/R1~VP15-G-F7/R7) (��1), �������︲��Vp15���������С���3��ͻ����(vp-like4��vp15-umu1��vp15-DR1126)������DNAΪģ��, �ֱ�����Vp15����Ƭ��, ��PCR�����ͱ��������ҵ���\������˾����ʹ��DNAMAN�����ȶԷ���������, ȷ������ͻ�������PCR������ϵΪ2 × KOD buffer 25 µL, ����(10 µmol L-1) 0.75 µL, ģ��DNA 2 µL, dNTPs (2.5 µmol L-1) 5 µL, KOD FXø(1.0 U µL-1) 1 µL, ������ˮ��50 µL��PCR��������Ϊ94°CԤ����5 min; 94°C����10 s, 59°C�˻�30 s, 68°C����1 min, 35��ѭ��; 68°C����7 min����1%��֬��������Ӿ���Ŀ��������Table 1
��1
��1���о����õ�����
Table 1Primers used in this study
| ���� Primer | �������� Forward sequence (5°-3°) | �������� Reverse sequence (5°-3°) |
|---|---|---|
| VP15-G-F1/R1 | AAGGACGCGAGAGTTTTTGA | GGTGAGCGAGTAACCCATGT |
| VP15-G-F2/R2 | ACATGGGTTACTCGCTCACC | CTTCCTGGTAGCGAGACGTG |
| VP15-G-F3/R3 | GCTCGTTCACCATCCAGAAT | AGCAAAGCCCTTTCTGAACA |
| VP15-G-F4/R4 | GGGCTTTGCTAAGATGTCTCC | GGACAATACATAACTGCTCCCTAAA |
| VP15-G-F5/R5 | TCTTCCACTCTTCCTACCAGCTA | AGGAGCCAACAACTGCTGAT |
| VP15-G-F6/R6 | GAACAATCGAGCCTTCTTCG | TCGTCCTCCTTGACCACTTC |
| VP15-G-F7/R7 | CACCAACTGCAAGATCGAAA | CCAACAGACGTCAACACCAG |
| VP15-CDS-F1/R1 | GGAACTCTCTGATACCTGGCTTT | CAGTTCCGTGATTAGCCGC |
| Zm-GAPDH-F/R | CCCTTCATCACCACGGACTAC | AACCTTCTTGGCACCACCCT |
�´��ڴ�
1.6 vp-like4��vp15�ٴε�λ����
���ݲ�����, ���vp-like4��vp15-umu1��vp15-DR1126 �ֱ�����������(VP15-G-F2/R2��VP15-G-F3/R3)ȷ�������Ӻ�ͻ����, ���ٴν��е�λ���顣ȡvp-like4�Ӻ�ͻ���廨�۷ֱ��ӽ�vp15-umu1��vp15-DR1126�Ӻ�ͻ����ֲ��; ͬ��, �ֱ�ȡvp15-umu1��vp15-DR1126�Ӻ�ͻ���廨���ӽ�vp-like4�Ӻ�ͻ����ֲ�ꡣ�ڷۺ�30 d, ������Ҷ, ���������뷢ѿ�������, ��ͳ�Ʒ���ȡ�1.7 RNA����ȡ���������������
���ڷۺ�30 d���ҵ��뷢ѿ����������, ��ȥ��Ƥ, ��ֲ����RNA��ȡ�Լ���(ͬ��)��ȡ����RNA�����÷�ת¼�Լ���(����ȫʽ�����\������˾, Cat# AU311-02)��ת¼ΪcDNA, ��cDNAΪģ��, �����������VP15-CDS-F1/R1, ����GAPDHΪ�ڲ�����(��1), ��ABI 7300ʵʱ����PCR���Ͻ���ʵʱ����������PCR��ϵΪdye I 0.4 µL, 10 × qMix 10 µL, ����(10 µmol L-1) 0.5 µL, ģ��cDNA 1 µL, ������ˮ��20 µL��PCR��������Ϊ95°CԤ����2 min, 95°C����10 s, 60°C�˻�15 s, 72°C����30 s, ѭ����Ϊ40, ��72°C����30 s�ռ�ӫ��, �������۽�����, ����ʽ2-ΔΔCt������������뷢ѿ������Vp15������������졣2 ��������
2.1 ͻ����vp-like4�ı��ͺ��Ŵ�����
һ�������, �����ڷۺ�30~40 d, ����������������֯��ʼӲ��, ��־��������ʼ����, �ڷۺ�60 d���������ﵽ��ȫ���졣����, �뷢ѿͻ����vp-like4���Ӻ������Խ��ڷۺ�30 d����, ���ڹ��������Թ۲쵽���뷢ѿ�����ķ���(ͼ1-A)�������뷢ѿ�������ڹ����ջ��, �ȷ�����ѿ�Ѹɿ�����(ͼ1-B, C), ��˽�ͬһ�����ϵ�������������ֲ���Խ�, �۲�vp-like4�뷢ѿ���Ŵ��ȶ��Ժͷ�����������������Խ�, ����vp-like4��ͻ����״���ȶ��Ŵ�������2��ʾ, ���ѡ���뷢ѿ����Ĺ���, ͳ���������뷢ѿ����, ���������������ȷ���3∶1, ����vp-like4ͻ������뷢ѿ��״�ܵ������Ժ˻�����ơ� ��ʾԭͼ|����ԭͼZIP|����PPT
��ʾԭͼ|����ԭͼZIP|����PPTͼ1vp-like4ͻ�����뷢ѿ����
A: vp-like4�Ӻ�ͻ�����ڷۺ�30 d�����������������뷢ѿ����; B:vp-like4�Ӻ�ͻ�����ڷۺ�60 d�����������������뷢ѿ����; C: �������������(WT)���뷢ѿ����(vp)�����=1 cm��
-->��
A: normal and viviparous kernels on a vp-like4 heterozygous ear at 30 days after self-pollination; B: normal and viviparous kernels on a vp-like4 heterozygous ear at 60 days after self-pollination; C: mature normal (WT) and viviparous kernels (vp). Bar = 1 cm.
-->
Table 2
��2
��2vp-like4�Ӻ�ͻ�����Խ��ڷۺ������������뷢ѿ�����ķ������
Table 2Segregation of normal and viviparous kernels on vp-like4 self-pollinated heterozygous ears
| ��� Year | �ص� Location | ֲ������� Plant genotype | ��������Kernel phenotype | |||
|---|---|---|---|---|---|---|
| �������� Normal | �뷢ѿ���� viviparous | ���� Total | χ2 (3:1) | |||
| 2014 | ����Hainan | vp-like4/+ | 355 | 127 | 482 | 0.398 |
| 325 | 111 | 436 | 0.027 | |||
| 2016 | ����Beijing | vp-like4/+ | 129 | 47 | 176 | 0.189 |
| 225 | 70 | 295 | 0.190 | |||
�´��ڴ�
2.2 vp-like4����λ
BSR-Seq����λ��������[21], �ڵ�5Ⱦɫ������һ�����Եķ�, ��P > 0.5Ϊ��, ��vp-like4��λ������5Ⱦɫ��173.8~175.6 Mb������(ͼ2)��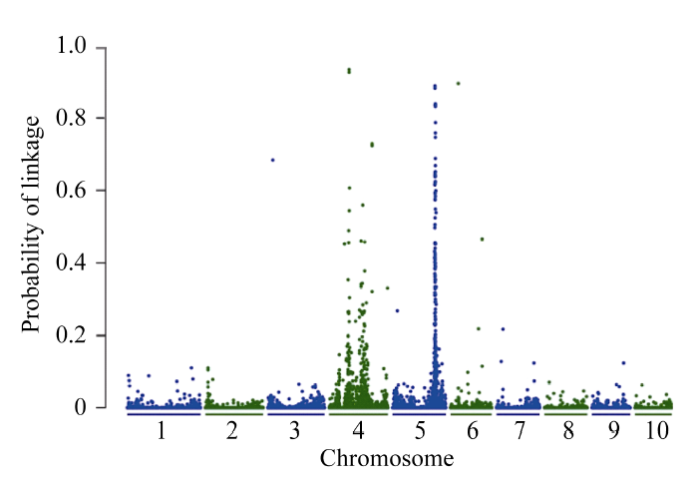 ��ʾԭͼ|����ԭͼZIP|����PPT
��ʾԭͼ|����ԭͼZIP|����PPTͼ2����BSR-Seq������vp-like4ͻ�������ij���λ
-->Fig. 2Gene mapping of the vp-like4 mutant by the BSR-Seq strategy
-->
2.3 vp-like4��vp15�����
��vp-like4����ӽ�vp15-umu1��vp15-DR1126��������, ���뷢ѿ����Ĺ���(ͼ3-A, B); ͬʱ, vp15-umu1��vp15-DR1126�ֱ����ӽ�vp-like4��������, Ҳ���뷢ѿ�Ĺ���(ͼ3-C, D)����һ�������˵��, vp-like4ͻ����ܿ��ܾ���Vp15�����λͻ���塣 ��ʾԭͼ|����ԭͼZIP|����PPT
��ʾԭͼ|����ԭͼZIP|����PPTͼ3�����Ӻ�ͻ�����vp-like4��vp15���е�λ����
A, B: ��vp-like4�Ӻ�ͻ�����۷ֱ��ӽ�vp15-umu1(A)��vp15-DR1126(B)�Ĺ��������뷢ѿ������C, D: ��vp15-umu1(C)��vp15-DR1126(D)�ֱ����ӽ�vp-like4�Ĺ��������뷢ѿ������
-->Fig. 3Allelism test of vp-like4 with vp15 by heterozygous mutants
A, B: viviparous kernels were respectively emerged on vp15-umu1 (A) and vp15-DR1126 (B) heterozygous ear crossed by the mixed pollen of vp-like4 heterozygous plants; C, D: viviparous kernels were emerged on vp-like4 heterozygous ear crossed by the mixed pollens of vp15-umu1 (C) and vp15-DR1126 (D) heterozygous plants.
-->
2.4 vp-like4ͻ��λ�����
Ϊ��һ����֤vp-like4��vp15�ǵ�λ����ȷ��vp-like4��ͻ��λ��, ��Vp15����������Ϊ�ο�, �����7 ��PCR����(��1), �ֱ�����vp-like4��vp15-umu1��vp15-DR1126������DNA, ���������-310 bp��4962 bp��Ƭʽ������Vp15���������С����������������, vp-like4ͻ������Vp15�����2������ĩ�˼�3°�Ƿ�������60 bp���ȱʧ; ��vp15-umu1��vp15-DR1126���ڵ�2�������Ӳ�ͬλ����Mutatorת���Ӳ���(ͼ4)���ɴ�˵��ͻ����vp-like4��vp15��Vp15����ͻ�䷽ʽ��ͬ, vp-like4��vp15һ���µĵ�λͻ�����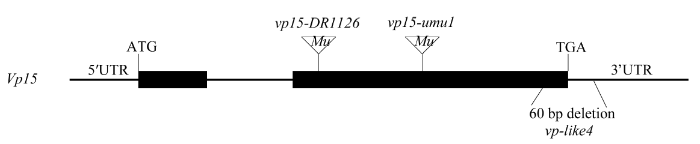 ��ʾԭͼ|����ԭͼZIP|����PPT
��ʾԭͼ|����ԭͼZIP|����PPTͼ4Vp15����ṹʾ��ͼ��3��ͻ�����ͻ��λ��
-->Fig. 4Gene structure of Vp15 and mutation site of three mutants
-->
���ݲ�����, ������������ȷ����vp-like4��vp15-umu1��vp15-DR1126�Ӻ�ͻ����, �����Ӻ�ͻ������һ�ν��е�λ����(��������������뷽������)���ڷۺ�30 d, ������Ҷ����뷢ѿ�������, ������vp-like4�����ӽ�vp15-umu1��vp15-DR1126, ����vp15-umu1��vp15-DR1126�����ӽ�vp-like4����, ���������뷢ѿ�����ķ���, ��ͳ�Ʒ������������������뷢ѿ��������3∶1�����(��3)����һ��֤��vp-like4ͻ������Vp15�����λͻ���塣
Table 3
��3
��3vp-like4��vp15ͻ�����λ����
Table 3Allelism test of vp-like4 with vp15
| ��ĸ�������� Parental genotype | �������� Kernel phenotype | |||
|---|---|---|---|---|
| ��������Normal | �뷢ѿ����viviparous | ����Total | χ2 (3:1) | |
| vp-like4/+ × vp15-umu1/+ | 217 | 68 | 285 | 0.141 |
| vp-like4/+ ×vp15-DR1126/+ | 123 | 40 | 163 | 0.002 |
| vp15-DR1126/+ ×vp-like4/+ | 262 | 83 | 345 | 0.116 |
| vp15-umu1/+ ×vp-like4/+ | 239 | 84 | 323 | 0.124 |
�´��ڴ�
2.5 Vp15������ͻ����vp-like4�еı���
�ֱ�ȡvp-like4��vp15-umu1��vp15-DR1126�Ӻ�ͻ��������ϵ������������뷢ѿ����, ����ʵʱ����ӫ��PCR, ��GAPDHΪ�ڲμ��Vp15����ı�����(��1)�������ͼ5��ʾ, ��������vp-like4������vp15-umu1��vp15-DR1126��, �뷢ѿ����Vp15����ı�����������������������, ����vp-like4�м����ȱʧ, ����vp15-umu1��vp15-DR1126��Mutatorת���ӵIJ���, ��Ӱ����Vp15�����������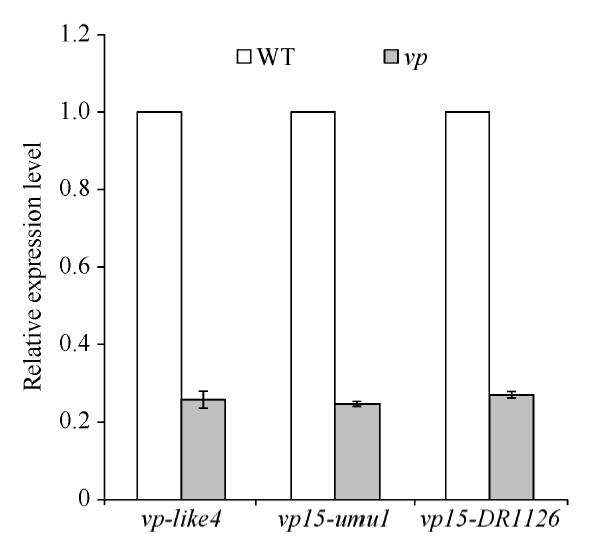 ��ʾԭͼ|����ԭͼZIP|����PPT
��ʾԭͼ|����ԭͼZIP|����PPTͼ5ʵʱ����PCR���Vp15����ֱ���3��ͻ����vp-like4��vp15-umu1��vp15-DR1126�е�����(WT)���뷢ѿ(vp)�����еı�����
-->Fig. 5Gene expression levels of Vp15 in the normal (WT) and viviparous (vp) kernels of three mutants, vp-like4, vp15-umu1, and vp15-DR1126, by quantitative real-time PCR analysis
-->
3 ����
������, ��ͨ����������Ѹ�ٷ�չҲ�ٽ��˻���λ�����ķ�չ������BSR-Seq (Bulked Segregant RNA-seq)�����ǻ�Ϸ������(BSA, Bulked Segregant Analysis)������RNA����(RNA-Seq)������Ϸ�չ�����ĸ�Ч��λ������·���������BSR-Seq����, ѡ�����Ⱥ���о��м�����״�ĸ���ֱ�2��������, ��ȡ2��������RNA����ת¼�������RNA-Seq������ȡ�����ķ��ӱ��, ����Щ���ӱ�ǽ��е�λ����������, ���ݵ�λ�����ڲ�ͬ��ϳصķֲ�����, ��λĿ�����ͬʱ, Ҳ��������RNA-Seq���ݽ��к�ѡ�����Ԥ��[21]���������������ֻ������Ϊ���ӵ�����, BSR-Seq������Ϊ��Ч, ��ΪBSR-Seqֻ��mRNA����, ���漰�ظ����к���������, �ȼ����˲���ɱ�, Ҳ�����Ч��[23]�����о�������һ���µ��뷢ѿͻ����vp-like4, �ܵ����Ժ˻�����ơ�����ʹ��BSR-Seq������ͻ���忪չ����λ, ��Ѹ�ٻ�ö�λ���䡣�ü�����ʹ�ô������˹�����, ������ʵ������, Ϊ����ͻ�������Ķ�λ��������ķ��㡣������֪, ��λ������ͨ���ӽ�ʵ����ӽ������״�ķ���������з���, ��ȷ������ĵ�λ�Լ����Ŵ����롣һ�������, ��λ�������������������Դ���ͻ��������ӽ�[24,25]���ڱ��о���, �����뷢ѿ����ͻ������ǰ��ѿ��������ѿ���ջ�ʱ����, ����, ������δ��¡vp-like4����֮ǰ, ��ȷ������Ϊ�Ӻ�ͻ���廹�Ǵ���Ұ����, ���, ���о����û�۵ķ������г����ĵ�λ�Լ�⡣ȡ���ۻ�۵�ͬʱҲ��ȡ�����еĵ�������Խ�, �����Խ������뷢ѿ�������ȷ֤��ȡ���۰���ͻ���廨�ۡ���֮, ���ܻ��۵�ֲ����˽��ܻ����ӽ��ڷ�, Ҳͬʱ������һ���Խ�, �������Խ������뷢ѿ�������ȷ֤���ܻ��۵������ͻ������롣���о���ȡ�����ķ���, ������֤��vp-like4��vp15�ĵ�λ�ԡ������Ļ���������֤��ȷ���Ӻ����λ�ӽ�, Ҳ֤����vp-like4ȷʵ��vp15�ĵ�λ����
�����о�������ʾVp15�������MPT�ϳ�øС�ǻ�, ��ø�����⸨����(MoCo)�ĺϳ�[12], MPT�ϳ�øС�ǻ��㷺��������˺�ԭ��������[26], ���������书��ȱʧ������ּ���[27,28,29]��MPT�ϳ�øС�ǻ���C���и߶ȱ��صĽṹ��, ��C�˵ĸʰ����ڴ���Ӧ����Ϊ�����ӹ�����������������Ҫ������[30]�����о���vp-like4�ڵ�2������ĩ�˼�3°�Ƿ�����ȱʧ60 bp���, �ƻ�VP15����C�˱��ؽṹ��, ����ʹ������������MoCo�ϳ�, ���²��������ϳ�ABA, ����Ϊͻ����������ĸ������ǰ�ȷ������, vp-like4ͻ������Vp15�����ͻ��, ����������vp15-umu1��vp15-DR1126ͻ����ֱ��ڵ�2����������Mutatorת���Ӳ����ͻ�䷽ʽ��ͬ, ��һ���µ�δ��������Vp15�����λͻ�䷽ʽ������, vp-like4ͻ������Vp15����60�������ȱʧ, �Ʋ��䵰�����з�������ͻ�䡣���Ǽ�vp-like4ͻ������Vp15����mRNA�ı�������������, �Ʋ�����������γ��˴����mRNA�Ӷ�������mRNA�ȶ��Բ��֮, ��ͻ����ļ����ͻ����¡, Ϊ��һ�������о�����Vp15�������ṩ���µ�ʵ����ϡ�
4 ����
�·���һ�������뷢ѿͻ����vp-like4, ��ͻ���������Ե�������ơ�ͨ��BSR-Seq����λ�ͻ����¡����, vp-like4��Vp15�����ڵ�2������ĩ�˼�3°�Ƿ�������60 bp���ȱʧ��ͻ��, ��һ���µ�Vp15�����λͻ���塣vp-like4����ͻ������ȷ��, Ϊ̽�������뷢ѿ�ķ��ӻ����ṩ���µ��Բġ�The authors have declared that no competing interests exist.
���������������������ϵ��
�ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
| [1] | . |
| [2] | . |
| [3] | . |
| [4] | . |
| [5] | . |
| [6] | . |
| [7] | . |
| [8] | . |
| [9] | . |
| [10] | . |
| [11] | . |
| [12] | . |
| [13] | . |
| [14] | . |
| [15] | . |
| [16] | . |
| [17] | . |
| [18] | . |
| [19] | . |
| [20] | . |
| [21] | . |
| [22] | |
| [23] | . |
| [24] | . . |
| [25] | . |
| [26] | . |
| [27] | . |
| [28] | . |
| [29] | . |
| [30] | . |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}