Sequence Characteristics and Expression Analysis of Potassium Transporter Gene GhHAK5 in Upland Cotton (Gossypium hirsutum L.)
CHAOMao-Ni1, WENQing-Yu2, ZHANGZhi-Yong1, HUGen-Hai1, ZHANGJin-Bao1, WANGGuo1, WANGQing-Lian1,*, 1 Henan Institute of Science and Technology / Henan Collaborative Innovation Center of Modern Biological Breeding / Henan Key Laboratory for Molecular Ecology and Germplasm Innovation of Cotton and Wheat, Xinxiang 453003, Henan, China2 Henan Academy of Agricultural Sciences, Zhengzhou 450002, Henan, China 通讯作者:王清连, E-mail: wangql985@163.com, Te1: 0373-3040337 收稿日期:2017-04-25 接受日期:2017-09-10 网络出版日期:2018-02-12 版权声明:2018作物学报编辑部作物学报编辑部 基金资助:本研究由国家自然科学基金项目(31601347, 31571600), 河南省科技攻关计划项目(172102110022), 河南省高等学校重点科研计划项目(15A210001)和河南科技学院高层次人才计划项目(2014004)资助
关键词:陆地棉;钾转运体基因;亚细胞定位;低钾;基因表达 Abstract KUP/HAK/KT potassium transporter gene family plays an important role in the absorption of K+ in plants. Identification and cloning of potassium transporter genes in cotton are helpful to improve potassium absorption, and facilitate the increase of yield and quality in cotton. Based on the genome sequence of upland cotton, we identified the potassium transporter gene GhHAK5 by homologous cloning, and amplified its CDS sequence from upland cotton variety Baimian 1. The CDS of GhHAK5 was 2451 bp, encoding a protein with 816 amino acids. The molecular weight and isoelectric point of this protein were 91.23 kD and 8.15, respectively. The GhHAK5 protein contained a conserved domain “K-trans” (Pfam02705) and a symbolic amino acid sequence GXXXGDXXXSPLY of KUP/HAK/KT family gene, and had 11 transmembrane domains. Phylogenetic tree analysis demonstrated that GhHAK5 had a close relationship with AtHAK5, followed by OsHAK5, and they all belong to Cluster I evolutionary cluster. The subcellular localization experiment indicated that GhHAK5 was located in plasma membrane, which is consistent with its function as a potassium transporter involved in K+ uptake. The expression of GhHAK5 was high in roots, but low in leaf, stem, petal, fiber and sepal, and induced by low potassium environment. These results provide a foundation for understanding the function of GhHAK5 and breeding varieties with high potassium efficiency in cotton.
通过同源克隆的方法鉴定到陆地棉基因组中[26]与拟南芥钾转运体基因AtHAK5高度同源的基因, 其ID号为Gh_D01G1760, 本研究将其命名为GhHAK5。基于该基因的CDS序列设计特异性引物, 利用PCR的方法从陆地棉品种百棉1号的根cDNA中扩增得到与GhHAK5基因大小一致的cDNA序列。测序结果表明, 该序列长2495 bp, 其中CDS全长为2451 bp, 与陆地棉基因组中GhHAK5基因的相似性达99.70%。进一步对GhHAK5基因编码的蛋白分析发现, GhHAK5蛋白编码816个氨基酸, 分子量和等电点分别为91.23 kD和8.15。运用NCBI的CDD在线工具进行GhHAK5蛋白功能保守域的分析表明, GhHAK5是钾转运体超级家族(K-trans superfamily)的一员, 并具有KUP/HAK/KT钾转运体家族的保守结构域“K-trans” (Pfam02705)(图1-A)。将GhHAK5蛋白与拟南芥KUP/HAK/KT家族成员进行氨基酸序列比对分析表明, GhHAK5与拟南芥KUP/ HAK/KT家族成员一样, 均具有该家族的标志性序列GXXXGDXXXSPLY (图1-B)。利用TMHMM工具进行跨膜区分析表明, GhHAK5含有11个跨膜区(图1-C)。 显示原图|下载原图ZIP|生成PPT 图1陆地棉GhHAK5蛋白的序列分析 A: GhHAK5的保守域分析; B: GhHAK5中含有的钾转运体标志性序列(用横线表出); C: GhHAK5的跨膜区预测。 -->Fig. 1Sequence analysis of GhHAK5 protein in upland cotton (Gossypium hirsutum L.) A: conserved domain of GhHAK5; B: symbolic amino acids sequence (underlined) of KUP/HAK/KT potassium transporter family harbored in GhHAK5; C: transmembrane domain prediction of GhHAK5. -->
2.2 陆地棉GhHAK5蛋白的进化关系分析
系统进化树表明, GhHAK5蛋白在进化上与拟南芥AtHAK5亲缘关系最近, 同源性达58.40%, 其次是水稻的OsHAK5, 同源性达55.90% (图2)。在植物中, KUP/HAK/KT家族基因大致可划分为4个进化簇Cluster I~IV [31], GhHAK5在进化上与AtHAK5和OsHAK5同属Cluster I进化簇(图2)。 显示原图|下载原图ZIP|生成PPT 图2陆地棉GhHAK5蛋白与拟南芥和水稻KUP/HAK/KT家族成员的进化关系分析 空心和实心分别代表水稻和拟南芥KUP/HAK/KT家族成员。 -->Fig. 2Phylogenetic relationships between GhHAK5 protein and KUP/HAK/KT family members in Arabidopsis and rice The hollow and solid small circles indicate the KUP/HAK/KT members of rice and Arabidopsis, respectively. -->
2.3 陆地棉GhHAK5蛋白的亚细胞定位
利用WoLF PSORT软件预测GhHAK5蛋白定位于质膜上。通过水稻原生质体瞬时表达系统进一步确定GhHAK5的亚细胞定位情况, 对于空载体在质膜、细胞质和细胞核中均可检测到绿色荧光蛋白, 而对于含有GhHAK5基因的表达载体只在质膜上检测到绿色荧光蛋白(图3), 表明GhHAK5是一个定位于质膜的蛋白, 该结果与预测结果一致。 显示原图|下载原图ZIP|生成PPT 图3陆地棉GhHAK5蛋白的亚细胞定位 a和d: 荧光下照片; b和e: 明场下照片; c和f: 荧光和明场叠加图。 -->Fig. 3Subcellular localization of GhHAK5 protein in upland cotton (Gossypium hirsutum L.) a, d: fluorescence images; b, e: bright-field images; c, f: overlap of fluorescence and bright-field images. -->
2.4 陆地棉GhHAK5基因的表达特性分析
qRT-PCR分析表明, GhHAK5在棉花的不同组织中表达量存在明显差异, 在根中表达量最高, 在茎、叶、花瓣、花萼和纤维中表达量很低(图4-A); 低钾处理后, GhHAK5在根中表达量显著上调(图4-B), 表明它是一个响应低钾胁迫诱导表达的基因。 显示原图|下载原图ZIP|生成PPT 图4陆地棉GhHAK5基因表达水平 A: GhHAK5在陆地棉不同组织中的表达分析(误差线代表3个独立重复的标准差); B: 低钾处理下GhHAK5在根中的表达分析(CK: normal potassium; LK: low potassium; **: P < 0.01; 误差线代表3个独立重复的标准差)。 -->Fig. 4Expression level of GhHAK5 gene in upland cotton (Gossypium hirsutum L.) A: organ-specific expression of GhHAK5 in upland cotton (error bars represent standard errors of three independent repetitions); B: expression of GhHAK5 in roots under low potassium (CK: normal potassium; LK: low potassium; **: P < 0.01; error bars represent standard errors of three independent repetitions). -->
ZhangY, LiN, HuW, GaoY, Yao YK, Tang MY.Bud & boll falling, yield and quality of cotton affected by rates of potassium .Xinjiang Agric Sci, 2012, 11: 2088-2095 (in Chinese with English abstract) [本文引用: 1]
[4]
MianA, Oomen RJ, IsayenkovS, SentenacH, Maathuis FJ, Véry AA.Over-expression of an Na+- and K+-permeable HKT transporter in barley improves salt tolerance .Plant J, 2011, 68: 468-479 [本文引用: 1]
Gao XZ, Ma WQ.Changes of soil nutrient contents and input of nutrientsin arable of China .Plant Nutr Fert Sci, 2000, 6: 363-369 (in Chinese with English abstract) [本文引用: 1]
Zhang ZY, Wang QL, Li ZH, Duan LS, Tian XL.Effect of potassium deficiency on root growth of cotton (Gossypium hirsutum L.) seedlings and its physiological mechanisms involved . Acta Agron Sin, 2009, 35: 718-723 (in Chinese with English abstract) [本文引用: 1]
Kong XQ, LuoZ, Li CD, Dong HZ.Molecular mechanisms of premature senescence in cotton .Cott Sci, 2015, 27: 71-79 (in Chinese with English abstract) [本文引用: 1]
Liu DQ, LiuR.Correlation analysis between soil fertility and premature senescence of transgenic cotton .Soils & Fert, 2002, (6): 41-42 (in Chinese with English abstract) [本文引用: 1]
[9]
PettigrewW, Meredith JrW.Dry matter production, nutrient uptake, and growth of cotton as affected by potassium fertilization .J Plant Nutr, 1997, 20: 531-548 [本文引用: 2]
Li ST, Xing SL, ZhangY, Cui RZ.Application rate and time of potash for high cotton yield, quality and balance of soil potassium .Plant Nutr Fert Sci, 2016, 22: 111-121 (in Chinese with English abstract) [本文引用: 2]
Song MZ, Mao SC, Xing JS, Yang HY.Effects of potassium on photosynthetic matter accumulation and yield .Acta Gossipii Sin, 1994, 6(suppl): 52-57 (in Chinese with English abstract) [本文引用: 2]
[12]
Ahn SJ, ShinR, Schachtman DP.Expression ofKT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake . Plant Physiol, 2004, 134: 1135-1145 [本文引用: 3]
[13]
Véry AA, Nieves-CordonesM, DalyM, KhanI, FizamesC, SentenacH.Molecular biology of K+ transport across the plant cell membrane: what do we learn from comparison between plant species? J Plant Physiol, 2014, 171: 748-769 [本文引用: 1]
[14]
MäserP, ThomineS, Schroeder JI, Ward JM, HirschiK, SzeH, Talke IN, AmtmannA, Maathuis FJ, SandersD, Harper JF, TchieuJ, GribskovM, Persans MW, Salt DE, Kim SA, Guerinot ML.Phylogenetic relationships within cation transporter families ofArabidopsis . Plant Physiol, 2001, 126: 1646-1667 [本文引用: 1]
[15]
YangZ, GaoQ, SunC, LiW, GuS, XuC.Molecular evolution and functional divergence of HAK potassium transporter gene family in rice (Oryza sativa L.) . J Genet Genomics, 2009, 36: 161-172 [本文引用: 1]
[16]
ZhangZ, ZhangJ, ChenY, LiR, WangH, WeiJ.Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.) . Mol Biol Rep, 2012, 39: 8465-8473 [本文引用: 1]
Chao MN, Wen QY, Zhang JY, Zhang ZY, DongJ, Yu YX.Identification and expression analysis of KUP/HAK/KT potassium transporter gene family in soybean [Glycine max (>L.) Merr .]. Acta Bot Boreal-Occident Sin, 2017, 37: 239-249 (in Chinese with Englishabstract) [本文引用: 1]
GierthM, MäserP, Schroeder JI.The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots . Plant Physiol, 2005, 137: 1105-1114 [本文引用: 2]
[20]
ZhaoS, Zhang ML, Ma TL, WangY.Phosphorylation ofARF2 relieves its repression of transcription of the K+ transporter gene HAK5 in response to low potassium stress . Plant Cell, 2016, 28: 3005-3019 [本文引用: 1]
[21]
Wang YH, Garvin DF, Kochian LV.Rapid induction of regulatory and transporter genes in response to phosphorus, potassium, and iron deficiencies in tomato roots. Evidence for cross talk and root/rhizosphere-mediated signals .Plant Physiol, 2002, 130: 1361-1370 [本文引用: 2]
[22]
Santa-María GE, RubioF, DubcovskyJ, Rodríguez-NavarroA. The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter . Plant Cell, 1997, 9: 2281-2289 [本文引用: 2]
XuJ.Cloning and Functional Characterization of Potassium Channel Gene GhAKT1 and Potassium Transporter Gene GhKT2 from Cotton (Gossypium hirsutum L.) . PhD Dissertation of China Agricultural University, Beijing, China, 2014 (in Chinese with English abstract) [本文引用: 3]
MuC, AnJ, Li TT, Wang YR, Zhang MC, Tian XL.Functional characterization ofGhAKT1 and GhKT2 in cotton(Gossypium hirsutum L.) using virus-induced gene silencing . Cott Sci, 2017, 29: 40-49 (in Chinese with English abstract) [本文引用: 2]
Guo ZK, YangQ, Yao QH, Wan XQ, Yan PQ.Transgenic tobacco withArabidopsis thaliana AtKup1 gene has high potassium content in leaves . J Chin Biotechnol, 2005, 25: 24-28 (in Chinese with English abstract) [本文引用: 1]
[26]
ZhangT, HuY, JiangW, FangL, GuanX, ChenJ, ZhangJ, Saski CA, Scheffler BE, Stelly DM, Hulse-Kemp A M, Wan Q, Liu B, Liu C, Wang S, Pan M, Wang Y, Wang D, Ye W, Chang L, Zhang W, Song Q, Kirkbride R C, Chen X, Dennis E, Llewellyn D J, Peterson D G, Thaxton P, Jones D C, Wang Q, Xu X, Zhang H, Wu H, Zhou L, Mei G, Chen S, Tian Y, Xiang D, Li X, Ding J, Zuo Q, Tao L, Liu Y, Li J, Lin Y, Hui Y, Cao Z, Cai C, Zhu X, Jiang Z, Zhou B, Guo W, Li R, Chen Z J. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement . Nat Biotechnol, 2015, 33: 531-537 [本文引用: 3]
[27]
ZhangZ, ChaoM, WangS, BuJ, TangJ, LiF, WangQ, ZhangB.Proteome quantification of cotton xylem sap suggests the mechanisms of potassium-deficiency-induced changes in plant resistance to environmental stresses .Sci Rep, 2016, 6: 21060 [本文引用: 1]
[28]
KumarS, NeiM, DudleyJ, TamuraK.MEGA: a biologist- centric software for evolutionary analysis of DNA and protein sequences .Brief Bioinform, 2008, 9: 299-306 [本文引用: 1]
[29]
EnglerC, KandziaR, MarillonnetS.A one pot, one step, precision cloning method with high throughput capability .PLoS One, 2008, 3: e3647 [本文引用: 1]
[30]
ChenS, TaoL, ZengL, Vega-Sanchez M E, Umemura K, Wang G L. A highly efficient transient protoplast system for analyzing defence gene expression and protein-protein interactions in rice .Mol Plant Pathol, 2006, 7: 417-427 [本文引用: 1]
Song YF, ZhangL, Dong LH, Jin YR, Shi SJ, BaiY, Liu CK, Feng GL, Feng XG, WangQ, Liu HB.Research progress on KUP/HAK/KT potassium transporter family in plant .J Agric Sci Tech China, 2013, 15: 92-98 (in Chinese with English abstract) [本文引用: 2]
[32]
Ashley MK, GrantM, GrabovA.Plant responses to potassium deficiencies: a role for potassium transport proteins .J Exp Bot, 2006, 57: 425-436 [本文引用: 1]
[33]
Rodrı?guez-NavarroA. Potassium transport in fungi and plants .Biochim Biophys Acta, 2000, 1469: 1-30 [本文引用: 2]
[34]
RubioF, Santamaría GE, RodrígueznavarroA.Cloning ofArabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells . Physiol Plant, 2000, 109: 34-43 [本文引用: 2]
[35]
Véry AA, SentenacH.Molecular mechanisms and regulation of K+ transport in higher plants .Annu Rev Plant Biol, 2003, 54: 575-603 [本文引用: 1]
[36]
SatoY, NanataniK, HamamotoS, ShimizuM, TakahashiM, Tabuchi-KobayashiM, MizutaniA, Schroeder JI, SoumaS, UozumiN.Defining membrane spanning domains and crucial membrane-localized acidic amino acid residues for K+ transport of a Kup/HAK/KT-typeEscherichia coli potassium transporter . J Biochem, 2014, 155: 315-323 [本文引用: 1]
[37]
Kim EJ, Kwak JM, UozumiN, Schroeder JI. AtKUP1: an Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell, 1998, 10:51-62 [本文引用: 2]
[38]
Maathuis FJ.The role of monovalent cation transporters in plant responses to salinity .J Exp Bot, 2006, 57: 1137-1147 [本文引用: 2]
[39]
TakahashiR, NishioT, IchizenN, TakanoT.High-affinity K+ transporterPhaHAK5 is expressed only in salt-sensitive reed plants and shows Na+ permeability under NaCl stress . Plant Cell Rep, 2007, 26: 1673-1679 [本文引用: 1]
[40]
GrossmanA, TakahashiH.Macronutrient utilization by photosynthetic eukaryotes and the fabric of interactions .Annu Rev Plant Physiol Plant Mol Biol, 2001, 52: 163-210 [本文引用: 1]
[41]
CoelloP, Polacco JC.ARR6, a response regulator from Arabidopsis, is differentially regulated by plant nutritional status . Plant Sci, 1999, 143: 211-220 [本文引用: 1]
Chao MN, Zhang ZY, Song HN, Li CQ, ZhangX, Hu GH, Zhang JB, Wang QL.Genome-wide identification and expression analysis of Pht1 family genes in cotton (Gossypium hirsutum L.) . Cott Sci, 2017, 29: 59-69 (in Chinese with English abstract) [本文引用: 1]
Han L. Nitrogen Efficiency of Genotype Selection and Related Gene Cloning in Cotton . MS Thesis of Shihezi University, 2011, Shihezi, China (in Chinese with English abstract) [本文引用: 1]

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT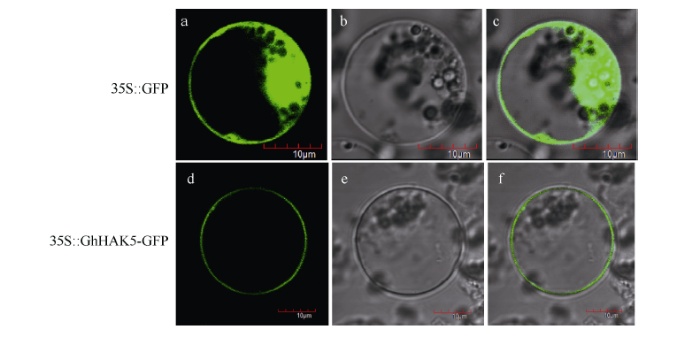 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}