 , 杨杰, 范方军
, 杨杰, 范方军Development and Application of the Functional Marker for Imidazolinone Herbicides Resistant ALS Gene in Rice
WANGFang-Quan, YANGJie, FANFang-Jun通讯作者:
收稿日期:2017-07-19
接受日期:2017-11-21
网络出版日期:2018-03-12
版权声明:2018作物学报编辑部作物学报编辑部
基金资助:
作者简介:
-->
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (7415KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
水稻是全世界近一半人口的主粮, 也是我国主要的粮食作物之一[1]。近年来, 随着我国新型城镇化和现代农业的发展, 水稻生产正在向集约化、规模化、机械化发展, 机插秧、直播等轻简栽培已成为发展趋势[2]。然而, 直播稻田容易滋生杂草及杂草稻, 严重影响水稻生长、产量及稻谷品质。人工除草的成本极高, 制约了水稻生产朝着高产、高效、低成本方向发展, 不利于发展现代农业。喷施除草剂是防治杂草危害的有效手段。
咪唑啉酮类除草剂是由美国氰胺公司开发的一类高效广谱低毒的除草剂, 目前已经有6种商品化产品, 包括咪唑烟酸、咪唑乙烟酸、咪草酸、咪唑喹啉酸、甲氧咪草烟和甲基咪草烟。咪唑乙烟酸, 又称咪草烟, 是一种常用于大豆田的高效除草剂, 能有效防除一年生禾本科杂草和阔叶杂草[3]。咪草烟的作用机理是通过抑制乙酰乳酸合酶(acetolactate synthase, ALS)的活性, 从而抑制侧链氨基酸的合成[4]。而作物对咪唑啉酮类除草剂的抗性则来自ALS基因一个或多个氨基酸变异而导致酶的结构变化[5,6,7]。已报道的ALS基因抗除草剂位点包括Gly95、Ala96、Ala122、Pro197、Asp376、Trp548、Ser627、Ser653和Gly654, 不同位点突变产生了抗除草剂水平的差异[7,8,9,10]。开展抗除草剂水稻育种与应用研究对于水稻轻简栽培、提升农业生产效益具有重要的理论和实践意义。
在先前的研究中, 我们从水稻品种资源材料中筛选到一个抗咪草烟的水稻材料金粳818。通过对金粳818 ALS基因的测序分析发现, 其ALS基因存在第1880位碱基G转换为A的突变, 即第627位氨基酸由丝氨酸(Ser, S)突变为天冬酰胺(Asn, N)。该突变与报道的抗除草剂红米稻strawhullR ALS基因功能突变位点相同[10]。
该基因可以通过回交转育到栽培品种中, 但传统上需要进行喷施除草剂逐代筛选鉴定, 其过程经历多代的杂交和回交, 育种周期长, 而且该基因为显性基因, 表型选择难以剔除杂合基因型, 更加大了育种周期和难度。因此, 利用分子标记辅助选择(marker-assisted selection, MAS)技术, 筛选鉴定基因型, 加快杂交后代抗性性状的早代稳定, 对于抗除草剂基因快速转育, 加快育种进程具有重要意义。本研究根据ALS基因第1880位点的变异, 设计了等位基因特异PCR标记, 能够有效选择抗除草剂基因型, 加快水稻材料的抗性稳定进程。
1 材料与方法
1.1 试验材料
抗除草剂水稻材料金粳818由本研究团队从7000多份水稻品种资源材料中筛选获得。金粳818由天津市水稻研究所利用津稻9618和津稻1007杂交经多年系谱法选育而成, 推测其抗性可能是在有残留除草剂的大豆田中产生。金粳818、日本晴、南粳40、南粳41、南粳44、南粳45、南粳46、南粳49、南粳9108、南粳5055、淮稻5号、苏秀867、武运粳21、武运粳24、武运粳27、徐稻3号、徐稻8号、镇稻88、镇稻99、连粳7号、常农粳7号为粳稻品种; 9311、IR36、南京16为籼稻品种。试验材料还包括以南粳9108和金粳818为亲本, 配制的杂交种及其衍生的F2群体; 以9311和金粳818为亲本, 通过多代回交和自交获得的抗除草剂高世代品系。将水稻材料种植于试验田隔离区, 防止花粉外漂, 以常规方法栽培。1.2 除草剂处理
所用除草剂为咪草烟(又称为咪唑乙烟酸), 由南京祥宇农药有限公司生产, 有效成分含量16%。将水稻种子催芽后, 按450 kg hm-2密度播种。在水稻长至二叶一心时, 排去田里的水, 将咪草烟(水剂)溶液稀释300倍, 每平方米喷洒70~80 mL, 喷洒24 h后复水, 14 d后调查抗性, 叶片全部枯萎或死亡为感, 植株存活为抗。1.3 抗性基因的克隆及序列分析
根据日本晴ALS基因编码区上下游序列(NC_008395.2)设计测序引物ALSs-F (5°-TCGCCC AAACCCAGAAAC-3°)和ALSs-R (5°-ATGCCAAGC ACATCAAACAA-3°)。以KOD-Plus (TOYOBO)高保真酶对金粳818基因组序列进行扩增, 扩增产物送往上海Invitrogen公司测序。用软件DNAMAN进行序列比对分析。1.4 分子标记设计
基于抗除草剂品种金粳818与感除草剂品种日本晴在ALS基因第1880位碱基的差异, 开发了11个分子标记(表1), 设置11个引物组合, 分别为S4/S2、S4/S3、S5/S2、S5/S3、S6/S2、S6/S3、S7/S2、S7/S3、S1/S8、S1/S9和S1/S10。部分引物在3°端引入了碱基错配。Table 1
表1
表1 ALS基因分子标记设计
Table 1Molecular markers design of ALS gene
| 编号 No. | 引物名称 Primer name | 引物序列 Primer sequence (5°-3°) |
|---|---|---|
| S1 | AS-ALS-F1 | GAGGCAATCATCGCTACTGG |
| S2 | AS-ALS-R1-1 | ACAAACCTAGACAGCAGGAAG |
| S3 | AS-ALS-R1-2 | CTCTTTATGGGTCATTCAGGTC |
| S4 | AS-ALSn-F2-1 | TGTGCTGCCTATGATCCtgAG |
| S5 | AS-ALSn-F2-2 | TGTGCTGCCTATGATCCCtAG |
| S6 | AS-ALSm-F2-1 | TGTGCTGCCTATGATCCCgAA |
| S7 | AS-ALSm-F2-2 | CATGTGCTGCCTATGATCtCgAA |
| S8 | AS-ALSn-R2-1 | TGTCCTTGAATGaGCCCCtAC |
| S9 | AS-ALSn-R2-2 | TGTCCTTGAATGCGCCCCtAC |
| S10 | AS-ALSm-R2-1 | TGTCCTTGAATGCGCCCCtAT |
| S11 | AS-ALSm-R2-2 | TGTCCTTGAATGCGCCCttAT |
新窗口打开
1.5 PCR扩增与电泳检测
用CTAB法提取待检测水稻叶片的基因组。以提取的基因组DNA为模板, 进行PCR扩增。PCR体系为DNA模板(20 ng µL-1) 2 µL, 10×PCR buffer 2 µL, MgCl2 (5 mmol L-1) 2 µL, dNTP (2 mmol L-1) 2 µL, 上游引物(2 μmol L-1) 2 µL, 下游引物(2 μmol L-1) 2 µL, Taq DNA聚合酶(5 U µL-1) 0.2 µL, ddH2O 7.8 µL。扩增条件为94°C 5 min; 94°C 30 s, 55°C 30 s, 72°C 30 s, 35个循环; 72°C延伸10 min, 结束反应。在Eppendorf Mastercycle热循环仪中进行PCR。扩增产物经琼脂糖凝胶电泳分离, 用凝胶成像仪拍照并记录结果。2 结果与分析
2.1 金粳818 ALS基因克隆及其功能标记的开发
金粳818抗除草剂的表型是由单基因控制的质量性状, 抗相对于感为显性。利用SSR标记将金粳818抗除草剂基因定位在第2染色体RM7413和RM7426之间, 其中RM7413与抗性基因紧密连锁。对这2个标记之间的基因分析发现, 前人报道的抗咪唑啉酮类除草剂靶标基因ALS位于这一区段[5,9,11-14]。因此, 我们对金粳818的ALS基因测序, 并与感除草剂粳稻品种日本晴和已报道感除草剂品种StrawhullS的ALS基因[10]比对。结果如图1所示, 金粳818与日本晴之间存在32处碱基变异, 与StrawhullS之间存在6处碱基变异, 金粳818与日本晴、StrawhullS在1593、1880和1927 bp处均存在变异。其中, 第1880 bp的变异导致第627位氨基酸由丝氨酸突变为天冬酰胺。根据Rajguru等[10]报道, 第1880 bp位点是水稻抗咪唑啉酮类除草剂的一个功能位点, 该位点的变异使得咪唑啉酮类除草剂不会抑制ALS蛋白活性, 从而对咪唑啉酮类除草剂产生抗性。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1ALS基因序列比对图
日本晴的ALS基因序列GenBank登录号为NC_008395.2, StrawhullS的ALS基因序列GenBank登录号为AY885673.1。
-->Fig. 1Sequencing alignment of the ALS gene
The ALS gene sequence of Nipponbare has a GenBank number NC_008395.2, the ALS gene sequence of StrawhullS has a GenBank number AY885673.1.
-->
基于金粳818在ALS基因1880 bp位点上由G到A的变异, 设计了11个引物标记(表1)。以日本晴(感除草剂品种, ALS-G)和金粳818 (ALS-A)的基因组为模板进行引物筛选, 以S4/S2、S4/S3、S5/S2、S5/S3、S6/S2、S6/S3、S7/S2、S7/S3、S1/S8、S1/S9和S1/S10引物组合进行PCR扩增。结果如图2所示, S4/S2及S4/S3仅能在日本晴品种中扩增, 但扩增效率较低。S6/S2、S7/S2和S7/S3仅能在金粳818中扩增。S1/S9能在日本晴中高效扩增, 在金粳818中有微弱扩增; 而S1/S10能在金粳818中高效扩增, 在日本晴中有微弱扩增。
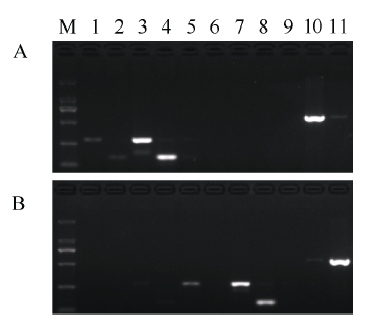 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2不同引物组合对日本晴和金粳818的PCR扩增结果
A为日本晴, B为金粳818; M为DL2000标记, 由大到小分别为2000、1000、750、500、250和100 bp; 泳道1~11分别为S4/S2、S4/S3、S5/S2、S5/S3、S6/S2、S6/S3、S7/S2、S7/S3、S1/S8、S1/S9和S1/S10。
-->Fig. 2PCR products of Nipponbare and Jinjing 818 using different primer pairs
A: Nipponbare; B: Jinjing 818; M: DL2000 marker (from up to down, 2000, 1000, 750, 500, 250, and 100 bp). Lanes 1 to 11 represent S4/S2, S4/S3, S5/S2, S5/S3, S6/S2, S6/S3, S7/S2, S7/S3, S1/S8, S1/S9, and S1/S10, respectively.
-->
基于上述扩增结果, 选择6对分子标记(S4/S2、S4/S3、S5/S2、S5/S3、S1/S9和S1/S10)进一步优化条件。PCR体系与之前相同, PCR程序中将退火温度提高到60°C。PCR产物经1%的琼脂糖凝胶电泳中分离, 于凝胶成像系统中拍照表明, S4/S2、S4/S3、S5/S2和S5/S3均不能有效扩增出目的条带(图3)。而S1/S9仅在日本晴品种中获得特异有效扩增, S1/S10只在金粳818中获得单一有效扩增。且这2个引物组合获得的条带明亮、清晰, 检测结果可靠。S1/S9能特异检测感咪草烟基因型(ALS-G), S1/S10能特异检测抗咪草烟基因型(ALS-A)。将S1/S9命名为F1N, S1/S10命名为F1M, 作为检测抗/感咪草烟的分子标记, 命名为AS-ALS。
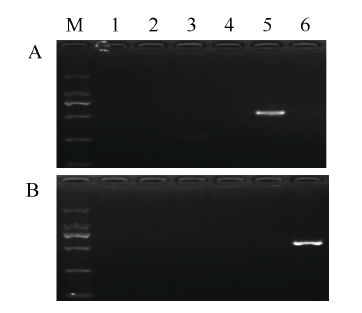 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3引物组合提高退火温度优化后PCR扩增结果
A为日本晴, B为金粳818; M为DL2000标记, 由大到小分别为2000、1000、750、500、250和100 bp; 泳道1~6分别为S4/S2、S4/S3、S5/S2、S5/S3、S1/S9和S1/S10。
-->Fig. 3PCR products after improving annealing temperature
A: Nipponbare; B: Jinjing 818; M: DL2000 marker (from up to down, 2000, 1000, 750, 500, 250, and 100 bp). Lanes 1 to 6 represent S4/S2, S4/S3, S5/S2, S5/S3, S1/S9, and S1/S10, respectively.
-->
2.2 F2群体的AS-ALS标记检测及表型分析
利用AS-ALS标记检测金粳818、南粳9108、南粳9108/金粳818杂交种及它们的93个F2单株, 所有携带ALS-A纯合和杂合基因型的F2单株都表现抗除草剂, 而所有携带ALS-G纯合基因型的F2群体单株均表现感除草剂(图4和图5)。分子标记基因型检测结果与表型鉴定结果完全对应。AS-ALS标记与抗/感除草剂表型完全共分离, 同时还能检测到抗性杂合基因型, 表明AS-ALS标记可用于抗咪草烟育种的精准选育, 在F2筛选抗性的纯合基因型, 可以实现早代选择。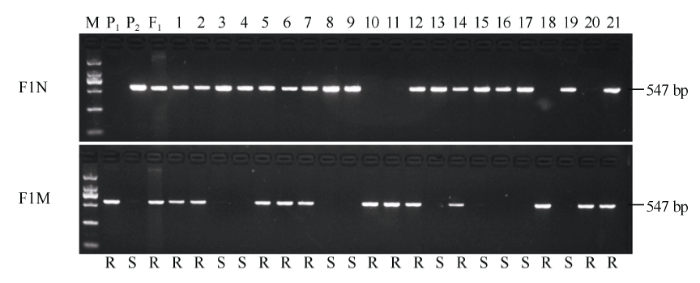 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4AS-ALS标记检测部分F2单株及表型对应情况
M为DL2000 DNA标记(由大到小分别为2000、1000、750、500、250和100 bp), P1为金粳818, P2为南粳9108, F1为南粳9108/粳9108/金粳818杂交种, 1~21为F2单株; R表示单株表现抗除草剂, S表示单株表现感除草剂。
-->Fig. 4Detection of parts of F2 populations using AS-ALS marker and the correspondence with phenotypes
M: DL2000 marker (from up to down, 2000, 1000, 750, 500, 250, and 100 bp); P1, Jinjing 818; P2: Nanjing 9108; F1: Jinjing 818/Nanjing 9108; 1-21: the plants of the F2 populations; R: resistant to herbicide; S: susceptible to herbicide.
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5喷施除草剂后F2植株表型
1为金粳818, 2为南粳9108, 3为南粳9108/金粳818杂交种, 4为ALS-A纯合基因型后代, 5为杂合基因型后代, 6为ALS-G纯合基因型后代。
-->Fig. 5Phenotypes of F2 populations after spraying herbicide
1: Jinjing 818; 2: Nanjing 9108; 3: Jinjing 818/Nanjing 9108; 4: the plant with ALS-A genotype; 5: the plant with ALS-G/A genotype; 6: the plant with ALS-G genotype.
-->
2.3 AS-ALS标记在回交育种中的应用
以金粳818为供体亲本, 以9311为轮回亲本连续回交3代, 每一代均对杂交种检测保留ALS-G/A基因型的单株, 与轮回亲本回交; 对BC3F2单株保留ALS-A纯合单株, 再自交一代, 结合农艺性状选择, 获得除草剂抗性稳定BC3F3株系。图6为利用AS-ALS标记对农艺性状优良的株系随机检测的结果, 表明这些株系都携带纯合ALS-A等位基因, 这些株系抗性稳定, 不再分离。因此, 利用AS-ALS标记辅助选择ALS-A基因可以快速获得除草剂抗性稳定的水稻材料。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6用AS-ALS标记检测高代品系及表型对应情况
M为DL2000 DNA标记(由大到小分别为2000、1000、750、500、250和100 bp), P1为金粳818, P2为南粳9108, 1~22为BC3F3群体单株; R表示单株表现抗除草剂, S表示单株表现感除草剂。
-->Fig. 6Detection of parts of the high generation lines using AS-ALS marker and the correspondence with phenotypes
M, DL2000 marker (from up to down, 2000, 1000, 750, 500, 250, and 100 bp); P1: Jinjing 818; P2: Nanjing 9108; 1-22: the plants of the BC3F3 populations; R: the plants resistant to herbicide; S: the plants susceptible to herbicide.
-->
2.4 用AS-ALS功能标记筛选水稻资源
如图7所示, 在检测的24份水稻品种资源中, 只有金粳818携带ALS-A等位基因, 属抗除草剂类型; 其他23份材料(20份粳稻: 日本晴、南粳40、南粳41、南粳44、南粳45、南粳46、南粳49、南粳9108、南粳5055、淮稻5号、苏秀867、武运粳21、武运粳24、武运粳27、徐稻3号、徐稻8号、镇稻88、镇稻99、连粳7号、常农粳7号; 3份籼稻: 9311、IR36、南京16)均为ALS-G等位基因, 表现感除草剂。标记检测的结果与基因的表型完全对应, 表明功能标记AS-ALS对籼稻和粳稻均具有多态性, 能同时用于粳稻和籼稻资源的筛选及抗除草剂遗传改良。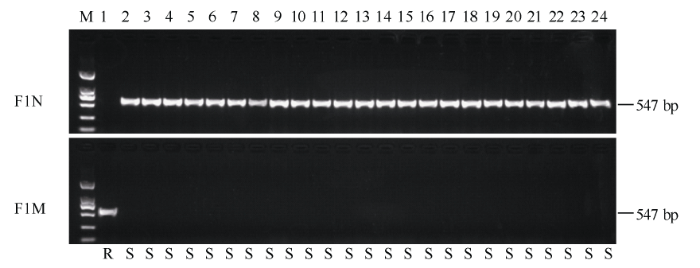 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7利用AS-ALS功能标记筛选水稻资源
M为DL2000标记, 由大到小分别为2000、1000、750、500、250和100 bp; 泳道1~24分别为金粳818、日本晴、南粳40、南粳41、南粳44、南粳45、南粳46、南粳49、南粳9108、南粳5055、淮稻5号、苏秀867、武运粳21、武运粳24、武运粳27、徐稻3号、徐稻8号、镇稻88、镇稻99、连粳7号、常农粳7号、9311、IR36和南京16。
-->Fig. 7Screening rice resources using AS-ALS marker
M: DL2000 marker (from up to down, 2000, 1000, 750, 500, 250, and 100 bp). Lanes 1-24 represent Jinjing 818, Nipponbare, Nanjing 40, Nanjing 41, Nanjing 44, Nanjing 45, Nanjing 46, Nanjing 49, Nanjing 9108, Nanjing 5055, Huaidao 5, Suxiu 867, Wuyunjing 21, Wuyunjing 24, Wuyunjing 27, Xudao 3, Xudao 8, Zhendao 88, Zhendao 99, Lianjing 7, Changnongjing 7, 9311, IR36, and Nanjing 16.
-->
3 讨论
随着国民经济的迅速发展和产业结构调整, 农村劳动力不断往城市转移, 水稻直播等轻简栽培生产发展迅猛[2]。但在直播稻田里容易发生草害, 尤其是与直播相伴的杂草稻危害, 已成为影响水稻品质和产量的重要因素。在江苏省镇江市扬中县一带的直播稻田中, 20世纪90年代后期就发现了红米杂草稻[15]。近年来, 在江苏以北地区随着直播稻生产方式的逐年扩大, 杂草稻危害面积呈逐年上升趋势。杂草稻分类上属于水稻, 利用稻田除草剂难以清除, 已成为水稻生产难以解决的世界性问题。美国有200多万公顷水稻, 主要生产方式是机器条播, 红米杂草稻也是影响美国水稻品质的主要问题之一[16,17]。咪唑啉酮类除草剂是一类广谱内吸性除草剂, 主要用于大豆田的高效除草, 能有效防除一年生禾本科杂草和阔叶杂草[3]。美国十多年前推出了抗咪唑啉酮类除草剂的水稻品种, 使得咪唑啉酮类除草剂在防除稻田杂草方面展现出了良好的应用前景。咪唑啉酮类除草剂是一类高效低毒性的除草剂, 但在偏碱性条件下降解较慢, 易对后茬油菜、番茄、马铃薯、高粱等作物造成药害。初步研究发现, 咪草烟当季残留对水稻生产影响比较大, 而多年使用咪草烟进行抗性鉴定的试验田对后茬小麦生产几乎没有影响, 为抗咪草烟水稻品种在江苏稻麦轮作区生产种植提供了可行性参考, 但对生产应用及环境影响还有待进一步研究。
咪唑啉酮类除草剂的靶标是乙酰乳酸合酶(acetolactate synthase, ALS), 该酶是细胞中合成侧链氨基酸的重要催化酶[18,19,20]。以ALS为靶点的除草剂能与植物体内的ALS蛋白结合, 阻碍侧链氨基酸如缬氨酸、亮氨酸、异亮氨酸的生物合成, 使细胞分裂被抑制, 杂草正常生长受到破坏而死亡[21,22,23]。ALS基因存在众多碱基变异位点, 部分变异引起编码氨基酸的改变而具有除草剂抗性。本研究发现金粳818携带的ALS基因第627位氨基酸由丝氨酸突变为天冬酰胺, 该位点的变异与Rajguru等[10]报道的突变点一致。抗除草剂水稻材料金粳818及调控位点的发现与利用, 对解决水稻直播中杂草稻的危害具有重要的理论和实践意义。
基因功能标记的开发和利用能够加快育种进程。功能标记可以用于基因型选择, 尤其是对显性基因, 可以将杂合基因型剔除, 加快目标基因的早代稳定, 可以省略表型鉴定过程, 节省成本, 加快育种进程[24,25]。基于ALS基因抗感等位基因第1880位由G到A的碱基变异, 设计了一对功能标记—— AS-ALS。该标记能有效区分感纯合基因型ALS-G、抗杂合基因型ALS-G/A、抗纯合基因型ALS-A, 是为针对单碱基突变开发的等位基因特异PCR分子标记, 只需要同一样本2次PCR就能区分3种基因型, 为开展ALS基因抗除草剂分子标记辅助育种奠定了基础。
4 结论
金粳818抗除草剂目标基因为ALS基因, 该基因第1880碱基为功能突变位点。基于该位点由G到A的碱基变异, 设计了一对功能标记AS-ALS。它是一个等位基因特异PCR标记, 由F1N和F1M组成。F1N能特异扩增ALS-G基因型, 属感除草剂类型; F1M能特异扩增ALS-A基因型, 属抗除草剂类型; F1N和F1M均能扩增出条带的样品为ALS-G/A杂合型, 属抗除草剂类型。AS-ALS标记与抗除草剂表型完全共分离。多代回交和自交并辅以AS-ALS标记选择, 能快速获得除草剂抗性稳定的水稻材料, 表明该标记能很好地用于分子标记辅助抗咪唑啉酮类除草剂水稻新品种的选育。The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . . |
| [3] | . . |
| [4] | . |
| [5] | . |
| [6] | |
| [7] | . |
| [8] | . |
| [9] | . |
| [10] | . |
| [11] | . |
| [12] | . |
| [13] | . |
| [14] | . |
| [15] | . . |
| [16] | . |
| [17] | . |
| [18] | . |
| [19] | . |
| [20] | . |
| [21] | . |
| [22] | . |
| [23] | . |
| [24] | . . |
| [25] | . . |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}