 , 习岗
, 习岗Characteristics of Ultra-weak Photon Emission from Sorghum Seeds during Germination
ZHAOYan-Yan, XIGang通讯作者:
收稿日期:2017-09-8
接受日期:2018-01-8
网络出版日期:2018-01-29
版权声明:2018作物学报编辑部作物学报编辑部
基金资助:
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (368KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
高粱用途广泛, 经济价值高, 并且耐盐碱和对土壤适应能力强, 在农业生产中占有重要地位[1,2]。近年来, 能源危机和环境污染日益严重, 高粱作为重要的能源作物, 越来越引起重视[3,4], 高粱萌发期的抗逆机理及其评价成为关注的热点, 有关研究集中在形态指标和生理生化指标方面[2,5-9]。由于形态指标的获取费时费力, 不能做到早期诊断; 生理生化指标大多来源于破坏性的试管实验, 不适应于珍稀物种, 判断结果也不准确, 亟待研究能够反映细胞代谢及其功能状态变化, 并能够做到无损检测的新技术, 种子萌发过程中超弱光子辐射的研究有可能满足这一要求。
生物超微弱光子辐射是普遍存在于所有生命系统中的一种连续发光现象, 其辐射强度远远低于普通的生物发光和化学发光[10]。种子萌发过程中的超微弱光子辐射, 包括萌发种子在暗中的自发光子辐射和在外界光诱导下的延迟光子辐射。生物光子学的研究表明, 来自于活细胞的超微弱光子辐射包含着丰富的生命信息, 自发光子辐射与生物体内的细胞分裂、细胞死亡、光合作用、生物氧化、细胞内和细胞间的信息传递与功能调节等重要的生命过程密切相关[11,12,13,14], 延迟光子辐射与生物系统的功能状态有关, 是细胞生理状态的灵敏指标, 已被应用于评价植物系统的功能状态和对环境胁迫的检测[15,16]。因此, 超微弱光子辐射的研究有可能为活体细胞代谢与功能状态提供一种非侵入性的光学活检新技术[17]。目前, 尽管已有红豆[18]、大麦[19]、豇豆[20]、小麦[21]、大豆[22]、玉米[23]等萌发种子超微弱光子辐射的研究报道, 但是, 有关辐射机理和信息挖掘的研究不够, 且未见萌发高粱超弱光子辐射的研究报道。
本文研究了高粱种子萌发过程中自发光子辐射和延迟光子辐射的变化规律, 并通过施加蛋白质合成的转录抑制剂放线菌素D (AMD)、翻译抑制剂环己亚胺(CHM)和呼吸抑制剂NaN3, 探讨了辐射的来源与意义, 研究结果对于揭示高粱种子萌发过程中超弱光子辐射的机理、评价高粱抗逆性、鉴定与开发种质资源等方面的无损检测新技术提供了参考。
1 材料与方法
1.1 材料培养与处理
供试材料为万农两糯1号高粱种子。参照文献[24]的方法, 通过预备试验选择呼吸抑制剂为650 μg mL-1的NaN3溶液, 对于蛋白质合成抑制剂, 采用美国Sigma公司生产的转录抑制剂放线菌素D (AMD)和翻译抑制剂环己亚胺(CHM)。参照文献[25]选取AMD溶液浓度为50 μg mL-1, CHM溶液浓度为100 μg mL-1。挑选形状、大小相近的饱满种子2400粒, 经质量分数为0.2%的HgCl2消毒和清洗后, 平均分成12组, 每组200粒, 放入铺有滤纸的培养皿, 分别加入等量的蒸馏水(对照组)、650 μg mL-1的NaN3溶液、50 μg mL-1的AMD溶液和100 μg mL-1的CHM溶液, 置PRX-1000A型人工培养箱(杭州得聚仪器设备有限公司), 在温度、相对湿度、PPFD分别为35℃、50%、50 μmol m-2 s-1的条件下培养。对照组和各处理组均设置3个重复, 从开始培养起, 每天定时测量各组各重复中随机选取的20粒高粱种子的数据。
1.2 种子鲜质量的测量
从种子培养当天开始(设为0点), 使用JA103H型电子天平(常州市幸运电子设备有限公司, 精度1 mg)每天定时随机测量各组中20粒萌发高粱种子的鲜质量。测量时用滤纸吸干种子表面水分, 设3组重复, 取平均值。1.3 自发光子辐射的测量
自发光子辐射的测量仪器和测量方法见文献[21]。根据测量系统样品杯的大小, 每次分别随机选取各组中20粒高粱种子, 用滤纸吸干种子表面液体, 放入样品室暗处理5 min后测量自发光子辐射。单位时间的光子数为自发光子辐射的强度, 用ISL表示, 单位为counts s-1。对照组、NaN3、AMD和CHM处理组均分别设置3个重复, 取3个重复测量的平均值。1.4 延迟光子辐射的测量
采用自制的测量系统, 仪器组成和测量方法见文献[21]。从种子培养开始, 每天定时取种子20粒, 用滤纸吸干表面溶液, 用蓝色LED激发后测量延迟光子辐射。光照时间为30 s, 测量时间为60 s。每组测量均设置3个重复, 每个重复测量3次, 取平均值。1.5 延迟光子辐射的动力学分析
参照文献[23]的方法, 将延迟光子辐射动力学曲线按照下式拟合。$I\left( t \right)={{I}_{\text{SL}}}+{{\frac{{{I}_{0}}}{{{\left( 1+\frac{t}{\tau } \right)}^{\beta }}}}^{{}}}$ (1)
式中, t为测量时间(s), I0为初始光子数(counts s-1), τ为相干时间(s), β为衰减常量, 是一个无量纲的常数, ISL为单位时间的自发光子辐射(counts s-1)。
延迟光子辐射积分强度I (T)可由式(1)的积分得到。
$I\left( T \right)={{I}_{\text{SL}}}T+\frac{\tau {{I}_{0}}}{\beta -1}\left[ 1-\frac{1}{{{\left( 1+{T}/{\tau }\; \right)}^{\beta -1}}} \right] $ (2)
式中, T为测量周期(s), 在本文中T=60 s。
1.6 统计分析
对以上测量值应用SPSS软件进行差异显著性分析, P<0.05为显著水平, P<0.01为极显著水平。运用Origin 9.0软件进行数据的计算与作图。2 结果与分析
2.1 高粱萌发过程中鲜质量的变化
从图1可见, 在吸胀后前 3 d, 对照组高粱种子鲜质量随着萌发时间快速增长, 在 3 d后增幅变缓。NaN3、AMD和CHM处理对种子鲜质量的增长有抑制作用, 其中, NaN3的抑制作用最大。在第7和第8天时, 对照组高粱种子鲜质量相对于萌发开始时(第0天)的相对增长率分别为130.11%和138.85%, AMD处理组为69.46%和75.37%, CHM处理组则为42.55%和45.15%, 和对照组相比较差异均达到了极显著水平(P<0.01), 表明AMD和CHM处理不同程度地抑制了高粱种子的萌发; 在第3天后, NaN3溶液处理的高粱种子鲜质量开始下降, 直至低于萌发开始时的质量, 表明NaN3溶液完全抑制了高粱种子的萌发。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1高粱萌发过程中鲜质量的变化
-->Fig. 1Changes of fresh mass of sorghum seeds during germination
-->
2.2 高粱萌发过程中自发光子辐射的变化
从图2可以看出, 在吸胀后前3 d, 对照组高粱种子自发光子辐射随着萌发时间快速增长, 在3 d后不再增加, 呈现出小幅震荡的趋势。AMD和 CHM处理使高粱种子自发光子辐射强度明显下降, 其中, CHM处理对自发光子辐射的影响要大于CHM处理。在萌发第8天时, 对照组高粱种子的自发光子辐射强度相对于第0天的相对增长率为36.26%, AMD处理组为20.28%, CHM处理组为13.45%; 经过NaN3处理的高粱种子在第3天自发光子辐射强度开始呈逐渐下降的趋势, 并在萌发后期稳定在较低(低于初始)水平, 表明NaN3完全抑制了高粱种子萌发过程中自发光子辐射的增长。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2高粱萌发过程中自发光子辐射的变化
-->Fig. 2Changes of spontaneous photon emission of sorghum seeds during germination
-->
2.3 高粱萌发过程中延迟光子辐射的变化
2.3.1 延迟光子辐射曲线及动力学参数的变化为了定量分析高粱种子萌发时延迟光子辐射的特征及差异, 将各处理条件下的延迟光子辐射数据按照式(1)拟合, 得到高粱种子萌发时的延迟光子辐射动力学参数见表1~表4。由表1~表4可见, 各参数的拟合优度都在0.99以上, 说明表1~表4中的动力学参数很好的描述了式(1)所表述的延迟光子辐射的动力学行为。
Table 1
表1
表1对照组萌发高粱延迟光子辐射动力学参数
Table 1Dynamical parameters for delayed photon emission of sorghum seeds during germination in control group
| 萌发时间Germination time (d) | 自发光子辐射Spontaneous photon emission, ISL (counts s-1) | 初始光子数 Initial photon number, I0 (counts s-1) | 相干时间 Coherence time, τ (s) | 衰减常量Attenuation coefficient, β | 拟合优度Determination coefficient, R2 |
|---|---|---|---|---|---|
| 0 | 7.64 | 237.28 | 0.26 | 0.84 | 0.993 |
| 1 | 8.57 | 256.05 | 0.27 | 0.87 | 0.992 |
| 2 | 9.51 | 298.72 | 0.30 | 0.95 | 0.996 |
| 3 | 10.38 | 333.00 | 0.32 | 0.96 | 0.992 |
| 4 | 10.39 | 337.77 | 0.35 | 0.97 | 0.995 |
| 5 | 10.17 | 323.95 | 0.32 | 0.96 | 0.991 |
| 6 | 10.41 | 344.55 | 0.34 | 0.97 | 0.995 |
| 7 | 10.27 | 331.29 | 0.33 | 0.96 | 0.996 |
| 8 | 10.42 | 353.76 | 0.38 | 0.97 | 0.996 |
新窗口打开
Table 2
表2
表2NaN3处理组萌发高粱延迟光子辐射动力学参数
Table 2Dynamical parameters for delayed photon emission of sorghum seeds during germination in NaN3 treatment group
| 萌发时间 Germination time (d) | 自发光子辐射Spontaneous photon emission, ISL (counts s-1) | 初始光子数 Initial photon number, I0 (counts s-1) | 相干时间 Coherence time, τ (s) | 衰减常量 Attenuation coefficient, β | 拟合优度 Determination coefficient, R2 |
|---|---|---|---|---|---|
| 0 | 7.64 | 234.28 | 0.25 | 0.83 | 0.992 |
| 1 | 7.78 | 245.65 | 0.26 | 0.84 | 0.996 |
| 2 | 7.96 | 269.56 | 0.27 | 0.86 | 0.993 |
| 3 | 8.02 | 278.51 | 0.28 | 0.87 | 0.991 |
| 4 | 7.56 | 233.87 | 0.23 | 0.81 | 0.991 |
| 5 | 7.16 | 228.73 | 0.22 | 0.78 | 0.991 |
| 6 | 7.27 | 229.76 | 0.23 | 0.80 | 0.994 |
| 7 | 7.16 | 223.19 | 0.21 | 0.80 | 0.993 |
| 8 | 7.18 | 226.09 | 0.22 | 0.78 | 0.996 |
新窗口打开
Table 3
表3
表3AMD处理组萌发高粱延迟光子辐射动力学参数
Table 3Dynamical parameters for delayed photon emission of sorghum seeds during germination in AMD treatment group
| 萌发时间Germination time (d) | 自发光子辐射Spontaneous photon emission, ISL (counts s-1) | 初始光子数 Initial photon number, I0 (counts s-1) | 相干时间 Coherence time, τ (s) | 衰减常量Attenuation coefficient, β | 拟合优度Determination coefficient, R2 |
|---|---|---|---|---|---|
| 0 | 7.64 | 235.44 | 0.26 | 0.84 | 0.994 |
| 1 | 8.17 | 252.95 | 0.26 | 0.86 | 0.994 |
| 2 | 8.67 | 290.64 | 0.29 | 0.93 | 0.996 |
| 3 | 9.13 | 318.03 | 0.32 | 0.95 | 0.992 |
| 4 | 9.15 | 322.93 | 0.32 | 0.95 | 0.994 |
| 5 | 8.98 | 305.41 | 0.32 | 0.95 | 0.995 |
| 6 | 9.11 | 317.80 | 0.33 | 0.95 | 0.994 |
| 7 | 9.06 | 309.35 | 0.32 | 0.94 | 0.992 |
| 8 | 9.19 | 324.50 | 0.34 | 0.94 | 0.992 |
新窗口打开
Table 4
表4
表4CHM处理组萌发高粱延迟光子辐射动力学参数
Table 4Dynamical parameters for delayed photon emission of sorghum seeds during germination in CHM treatment group
| 萌发时间Germination time (d) | 自发光子辐射Spontaneous photon emission, ISL (counts s-1) | 初始光子数 Initial photon number, I0 (counts s-1) | 相干时间Coherence time, τ (s) | 衰减常量Attenuation coefficient, β | 拟合优度Determination coefficient, R2 |
|---|---|---|---|---|---|
| 0 | 7.64 | 235.07 | 0.26 | 0.83 | 0.993 |
| 1 | 7.93 | 249.44 | 0.26 | 0.85 | 0.990 |
| 2 | 8.31 | 281.77 | 0.29 | 0.92 | 0.994 |
| 3 | 8.64 | 311.75 | 0.30 | 0.92 | 0.993 |
| 4 | 8.73 | 314.14 | 0.31 | 0.93 | 0.991 |
| 5 | 8.56 | 307.29 | 0.31 | 0.93 | 0.996 |
| 6 | 8.70 | 312.99 | 0.31 | 0.94 | 0.992 |
| 7 | 8.61 | 299.47 | 0.31 | 0.93 | 0.993 |
| 8 | 8.67 | 318.98 | 0.32 | 0.93 | 0.995 |
新窗口打开
2.3.2 延迟光子辐射积分强度的变化 延迟光子辐射曲线下的面积称延迟光子辐射积分强度, 用I (T)表示, 其数值可由式(2)求出。将表1~表4中的各动力学参数带入式(2), 可以得到各延迟光子辐射的积分强度I (T)。由图3可见, 在高粱萌发过程中各组的延迟光子辐射积分强度I (T)在前 3 d也呈现出不同程度增长的趋势。在第3天后, 对照组出现小幅震荡, 在第8天时相对于第0天的相对增长率为32.99%; AMD和CHM处理组和对照组类似, 在第8天时相对于第0天的相对增长率分别为20.38%和8.89%; 而经过NaN3处理的高粱种子则从第3天快速下降至较低水平。
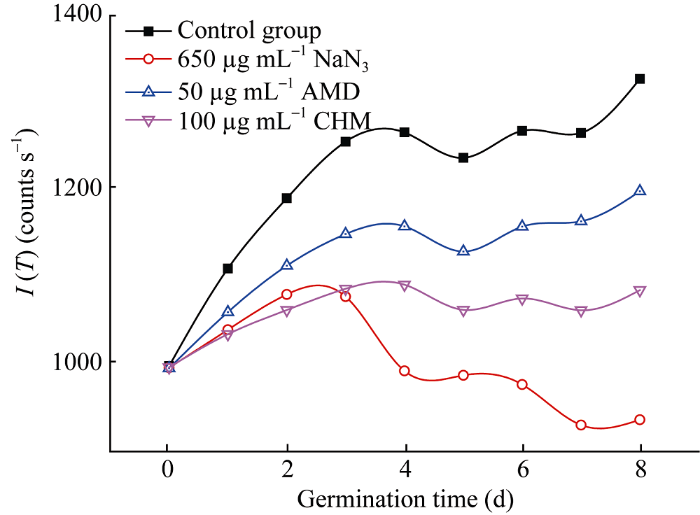 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3高粱萌发过程中延迟光子辐射积分强度的变化
-->Fig. 3Changes of the integrated intensity of delayed photon emission from sorghum seeds during germination
-->
3 讨论
种子在萌发过程中吸水膨胀, 启动DNA和RNA合成反应, 细胞开始分化, 各种水解酶和生物分子大量合成, 在宏观上表现为种子鲜质量的增加, 因此, 萌发种子的鲜质量可以作为表征种子萌发状况的宏观指标。为了探讨高粱萌发过程中自发光子辐射的来源, 本文进一步研究了蛋白质合成抑制剂AMD和CHM对萌发高粱自发光子辐射的影响。发现AMD和CHM对萌发高粱的自发光子辐射和鲜质量的增长都有同步的抑制作用, 但CHM的抑制程度比AMD大。由于AMD是蛋白质合成中转录过程的抑制剂, 50 μg mL-1的AMD足以完全抑制RNA的合成[26], 图1中AMD处理组萌发高粱种子鲜质量缓慢增加说明了萌发高粱种子中存在着长命mRNA, 由其产生的蛋白质合成的翻译过程不受AMD影响, 而图2揭示的AMD对自发光子辐射存在抑制作用说明高粱萌发过程中有一部分自发光子辐射来源于RNA合成反应。由于对DNA的抑制也会造成对RNA的抑制, 因此, 高粱萌发过程中的RNA/DNA合成反应可能是自发光子辐射的来源。
CHM是蛋白质合成中翻译过程的抑制剂, 作用于80S核糖体, 通过抑制蛋白质合成中的移位酶反应, 迅速而完全地阻断细胞中绝大部分蛋白质合成反应[26]。基于种子吸胀3 d以后的鲜质量源于新蛋白的大量合成[27], 图1显示的100 μg mL-1的CHM作用下种子鲜质量在吸胀3 d以后不再增长, 表明本研究采用CHM浓度完全抑制了高粱萌发过程中的蛋白质合成。由于在呼吸代谢中, 呼吸电子传递链中的电子漏会直接还原氧生成各种活性氧自由基, 这些自由基经过重组、裂解和歧化等一系列反应生成处于三重激发态的过氧化物, 或者单线态氧和激发态羰基, 后者经过退激后会产生光子辐射[11,19], 而种子吸胀萌发第3天后呼吸电子传递链中的主要复合蛋白体来自于蛋白质的从头合成[27], 由此推测, CHM通过阻断萌发高粱中呼吸电子传递链的主要组分复合蛋白体的合成, 形成对呼吸代谢的抑制, 造成了自发光子辐射的下降(图2)。CHM对萌发高粱自发光子辐射的抑制比AMD大, 说明呼吸代谢可能是高粱萌发过程中自发光子辐射的主要来源。由于NaN3是呼吸抑制剂, 其阻断线粒体呼吸电子传递链中的电子传递, 抑制线粒体吸收O2[28,29], 图2发现的NaN3作用下萌发高粱的自发光子辐射受到完全抑制证明了上述猜想。至于萌发3 d以后自发光子辐射呈现出小幅度震荡的原因, 可能是高粱种子萌发过程中, 细胞吸水膨胀后不断进行有丝分裂, 种子的呼吸代谢强度发生了周期性变化。
萌发高粱延迟光子辐射的动力学特征可以通过动力学参数和延迟发光积分强度定量表达。根据式(1), 初始光子数I0是激发光停止时的辐射强度, 在特定光照下, 与处于激发态的分子数有关, RNA/ DNA合成反应系统和呼吸代谢系统越完善, 能够处于高能激发态的分子数越多, I0就越大。由表1~表4可知, AMD、CHM和NaN3处理对萌发高粱种子初始光子数I0的抑制作用依次增强, 其中, NaN3完全抑制了I0的增长。产生这种现象的原因可能是AMD使RNA/DNA合成反应中能够处于激发态的分子数减少, CHM和NaN3分别通过阻碍呼吸电子传递链中各种复合蛋白体的合成和对呼吸电子传递链的影响抑制呼吸代谢中糖酵解/三羧酸循环(EMP/TCA), 导致呼吸代谢系统中能够处于激发态的分子数减少和呼吸代谢强度的降低, 后者又必然影响RNA/DNA合成反应系统[24], 引起初始光子数I0的进一步下降。
相干时间τ是延迟光子辐射动力学分析中的另一个重要参数。当处于激发态的分子关联性越好时, τ就越大。因此, τ值可以作为组织序性的量度[10]。对比表1和表4可知, AMD使萌发高粱种子τ值减小, 表明RNA/DNA合成反应系统的序性降低; CHM处理使萌发高粱种子的τ值下降, 表明CHM处理造成了呼吸代谢系统的紊乱; NaN3处理造成的萌发高粱种子τ值进一步减小, 则表明NaN3不但造成了呼吸代谢系统的紊乱, 也造成了RNA/DNA合成系统的破坏。
延迟光子辐射积分强度I (T)是综合反映延迟光子辐射特征的物理量。细胞代谢越强烈, 初始光子数I0越多, 相干时间τ越大, I (T)就越大, 因此, 延迟光子辐射可以反映细胞代谢的强度。本文的研究表明, AMD、CHM和NaN3都对萌发高粱延迟光子辐射积分强度I (T)有抑制作用, 这种抑制是通过对RNA/DNA合成系统和呼吸代谢系统的抑制所引起的, 由此看来, 通过对I (T)的测量和分析有可能对高粱萌发过程中细胞代谢系统发生的动态过程及其发生的变化进行实时、灵敏和无损伤的了解和判断, 进而在萌发种子抗逆性评价等方面发挥作用。
4 结论
萌发高粱自发光子辐射强度的增长与种子鲜质量的增长正相关(相关系数r为0.93121), 通过对高粱种子萌发过程中自发光子辐射强度的测量可以实现对种子萌发状况的无损检测。高粱萌发过程中RNA/DNA合成反应是自发光子辐射的来源之一, 呼吸代谢是其主要来源。延迟光子辐射积分强度是综合反映高粱种子萌发过程中延迟光子辐射特征的物理量, 由自发光子辐射、初始光子数和相干时间等动力学参数决定, 通过对高粱萌发过程中延迟光子辐射的测量和分析有可能对高粱种子萌发过程中细胞代谢系统发生的变化进行实时、灵敏和无损伤的了解和判断。The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . 以纤维高粱和普通高粱种子为材料,将NaCl、Na2 SO4和NaHCO3按不同比例混合,模拟15种盐度和碱度各不相同的复杂盐碱条件,采用培养皿纸上发芽法研究了混合盐碱胁迫对两种高粱种子萌发及幼苗生 长的影响,以探讨两种高粱种子和幼苗的耐盐碱能力及在盐碱胁迫下的变化规律.结果表明,低盐低碱环境促进了两种高粱种子的萌发;随着盐碱胁迫的增强,高粱 种子的萌发率、萌发指数与对照相比均显著下降,而纤维高粱的累积萌发率下降幅度较普通高粱小,表明纤维高粱对盐渍环境的适应性略高于普通高粱.在高粱幼苗 生长过程中,随着盐碱胁迫的增强,两种高粱的幼根和幼芽长均显著下降,且与对照均具有显著性差异.盐度对两种高粱的萌发率、萌发指数、幼苗长均有极显著影 响,盐度与pH相互作用对两种高粱的幼根长有极显著影响. . 以纤维高粱和普通高粱种子为材料,将NaCl、Na2 SO4和NaHCO3按不同比例混合,模拟15种盐度和碱度各不相同的复杂盐碱条件,采用培养皿纸上发芽法研究了混合盐碱胁迫对两种高粱种子萌发及幼苗生 长的影响,以探讨两种高粱种子和幼苗的耐盐碱能力及在盐碱胁迫下的变化规律.结果表明,低盐低碱环境促进了两种高粱种子的萌发;随着盐碱胁迫的增强,高粱 种子的萌发率、萌发指数与对照相比均显著下降,而纤维高粱的累积萌发率下降幅度较普通高粱小,表明纤维高粱对盐渍环境的适应性略高于普通高粱.在高粱幼苗 生长过程中,随着盐碱胁迫的增强,两种高粱的幼根和幼芽长均显著下降,且与对照均具有显著性差异.盐度对两种高粱的萌发率、萌发指数、幼苗长均有极显著影 响,盐度与pH相互作用对两种高粱的幼根长有极显著影响. |
| [2] | . 以8个品种(系)为材料,以两种碱性盐Na2CO3和NaHCO3摩尔比1∶1作为模拟碱胁迫,采用人工气候箱培养皿纸上发芽法培养高粱种子,测定不同品种(系)高粱发芽率、发芽势、根长、叶长、根重、叶重、计算出发芽指数与活力指数、分析不同胁迫处理下高粱各指标差异,计算各测定指标间相关性,用主成分分析法对高粱品种进行耐碱性强弱排序。结果表明:碱胁迫能抑制高粱种子萌发,降低幼苗生物量。多项萌发和生长参数的相对值之间存在极显著或显著的正相关。主成分分析结果表明,根干重、发芽势、叶鲜重、干重可作为高粱萌发期耐碱性鉴定的主要指标。根据主成分分析,8个高粱品种耐碱性的顺序为GZK4623A/千三623A/GZ623A千三623A/HN16HN16。 . 以8个品种(系)为材料,以两种碱性盐Na2CO3和NaHCO3摩尔比1∶1作为模拟碱胁迫,采用人工气候箱培养皿纸上发芽法培养高粱种子,测定不同品种(系)高粱发芽率、发芽势、根长、叶长、根重、叶重、计算出发芽指数与活力指数、分析不同胁迫处理下高粱各指标差异,计算各测定指标间相关性,用主成分分析法对高粱品种进行耐碱性强弱排序。结果表明:碱胁迫能抑制高粱种子萌发,降低幼苗生物量。多项萌发和生长参数的相对值之间存在极显著或显著的正相关。主成分分析结果表明,根干重、发芽势、叶鲜重、干重可作为高粱萌发期耐碱性鉴定的主要指标。根据主成分分析,8个高粱品种耐碱性的顺序为GZK4623A/千三623A/GZ623A千三623A/HN16HN16。 |
| [3] | . |
| [4] | . This review discusses the potential production for energy purposes of sweet sorghum ( Sorghum bicolor L.), a C 4 crop native to tropical areas with juicy stalks rich in sugars from which ethanol could be produced at lower costs than from starchy crops like maize. The growing interest in bioenergy and particularly in bioethanol is a great challenge for this relatively new crop that could be used for both thermo-electrical energy and biofuel. Nonetheless, the quantitative and qualitative production of sweet sorghum strongly depends on the use of appropriate and improved agronomic management techniques which is, in some aspects, still largely unknown. This review attempts to gather the sparse information on best agricultural practices for sweet sorghum, still very much a wild species in many aspects, while identifying the weak points that need to be deepened in further researches, especially under temperate climates. Sweet sorghum is a row crop and therefore the agronomic management and other well known cultural methods used for conventional row crops such as maize can be adapted. In general, low input requirements, low production costs, drought resistance, versatility, and high yields gives to sweet sorghum a better energy balance compared to other competing energy crops, especially if bagasse is also processed to energy. In non-traditional potential growing areas (such as in temperate climates of Europe) where productivity/adaptation improvements through genetically modified crops is not allowed, appropriate and sustainable agricultural practices constitute the most immediate option to improve yields. Based on scientific reports, research efforts seem particularly needed for harvesting techniques, and handling and storing. |
| [5] | . . |
| [6] | . Sweet is adapted to the hot and dry climatic conditions. It can be used for different products such as food, feed, fiber and fuel. This study was carried out to evaluate the effects of four irrigation intervals and three harvesting dates on stem height, stem diameter, biomass, juice volume, , and invert sugar of two sweet cultivars. The results showed that delay in irrigation intervals from 7 to 21 day significantly decreased all the above characteristics expect for invert sugar which was increased. The highest biomass (56.50 t ha(-1)) and content (11.35%) were obtained at 7 to 10 day irrigation interval. Both biomass and contents decreased significantly as harvesting date delayed from physiological maturity. Since both sweet cultivars, Keller and Sofra had similar biomass, content, and juice volume across irrigation intervals and harvesting dates, therefore it is suggested to plant either Keller or Sofra irrigated on 7 to 10 day interval and harvested at physiological maturity. |
| [7] | . <P>In semiarid regions of the Mediterranean basin, water and salinity stresses restrict crop establishment. The effects of salt and water stress on seed germination and early embryo growth (radicle and shoot growth) were investigated in laboratory in two cultivars of sweet sorghum [Sorghum bicolor (L.) Moench] – cv. ‘90-5-2′ and cv. ‘Keller’ – to verify how these stresses may limit crop growth during the very early stages of growing season. Six water potentials (ψ) of the imbibition solution (from 0 to 611.0 MPa) in NaCl or polyethylene glycol (PEG) for salt and water stress tests, respectively, were studied. Daily germination was recorded, and radicle and shoot lengths and dry weights (DWs) were measured 2 days after initial germination. Seed germination was reduced (8–30% lower than control) by water stress at ψ <610.6 MPa and was delayed (approx. +10 h) already at 610.4 MPa. Salt stress only prolonged germination time. Shoot and root growth was adversely affected by water stress (PEG), whereas the effect of salt stress was less relevant. Cv. ‘90-5-2’ was less sensitive than cv. ‘Keller’ to both stresses. These cultivars exhibited a greater sensitivity to abiotic stresses in terms of root and shoot growth; therefore, other parameters beside germination, such as early embryo growth rate, may help in evaluating sorghum lines for adaptation to semi-arid areas.</P> |
| [8] | . 【Objective】The objectives of the current study were to explore the response of different sorghum cultivars to salt stress, screen sorghum cultivars suitable for growing in saline soils, and provide a foundation for management of salt stress.【Method】 Thirty seeds of each of 42 sorghum cultivars were treated with 150 mmol61L-1NaCl or distilled water and incubated in Petri dishes for 10 days at humidity of 60%, light/dark of 12h/12h, illumination of 34 μmol61m-261s-1 and 28℃/25℃ of day/night. Germination and growth were assessed by counting the number of radicals and coleoptiles and leaves that had appeared on a daily basis from the fourth day and measuring their lengths and dry weights on the tenth day using destructive sampling. The cultivars were classified on the basis of salinity tolerance by means of principal component analysis and cluster analysis considering the range of data on germination and growth traits.【Result】The correlation analysis on the relative values of most germination and growth parameters showed positive relationships among several variables. The cultivars were ranked based on the principal components analysis and five groups on salinity tolerance were sorted for the 42 sorghum cultivars according to the cluster analysis.【Conclusion】Principal component analysis results showed that radical length, coleoptiles and leaf weight and germination rate were the most significant factors and are recommended as the main indexes to identify salinity tolerance of sorghum at germination. The analysis showed five of the 42 cultivars were highly salt tolerant, e.g. Liaoza 15, fourteen cultivars were salt tolerant, e.g. Shenshi 104, twelve cultivars were medium salt sensitive, e.g. Aoza 1, eight cultivars were salt sensitive, e.g Tieza 17, and three were highly salt sensitive, e.g. Longza 10. As this research only relates to ranking sensitivity during germination, further work is needed to determine the mechanism of tolerance and its persistence through the plant life cycle. . 【Objective】The objectives of the current study were to explore the response of different sorghum cultivars to salt stress, screen sorghum cultivars suitable for growing in saline soils, and provide a foundation for management of salt stress.【Method】 Thirty seeds of each of 42 sorghum cultivars were treated with 150 mmol61L-1NaCl or distilled water and incubated in Petri dishes for 10 days at humidity of 60%, light/dark of 12h/12h, illumination of 34 μmol61m-261s-1 and 28℃/25℃ of day/night. Germination and growth were assessed by counting the number of radicals and coleoptiles and leaves that had appeared on a daily basis from the fourth day and measuring their lengths and dry weights on the tenth day using destructive sampling. The cultivars were classified on the basis of salinity tolerance by means of principal component analysis and cluster analysis considering the range of data on germination and growth traits.【Result】The correlation analysis on the relative values of most germination and growth parameters showed positive relationships among several variables. The cultivars were ranked based on the principal components analysis and five groups on salinity tolerance were sorted for the 42 sorghum cultivars according to the cluster analysis.【Conclusion】Principal component analysis results showed that radical length, coleoptiles and leaf weight and germination rate were the most significant factors and are recommended as the main indexes to identify salinity tolerance of sorghum at germination. The analysis showed five of the 42 cultivars were highly salt tolerant, e.g. Liaoza 15, fourteen cultivars were salt tolerant, e.g. Shenshi 104, twelve cultivars were medium salt sensitive, e.g. Aoza 1, eight cultivars were salt sensitive, e.g Tieza 17, and three were highly salt sensitive, e.g. Longza 10. As this research only relates to ranking sensitivity during germination, further work is needed to determine the mechanism of tolerance and its persistence through the plant life cycle. |
| [9] | . 采用人工气候箱内培养皿培养,PEG-6000溶液模拟干旱胁迫环境,在萌发期以80、120、150和175 g L–1PEG-6000水溶液处理31个高粱品种,旨在根据高粱品种萌发期对不同干旱胁迫程度的响应,筛选出具有抗旱能力的高粱品种并探讨高粱萌发期抗旱性鉴定的方法。通过主成分分析法(PCA)和神经网络自组织映射(SOM)聚类分析法对各高粱品种进行抗旱性综合分析与评定。PCA结果表明,相对芽长、相对根长和相对萌发抗旱指数载荷量最大,将其作为萌发期高粱抗旱性筛选的主要评价指标,并对31个高粱品种抗旱性排序。通过SOM聚类分析将31个高粱品种按抗旱性强弱分为5类,吉杂305等4个品种为高度抗旱品种,HL5等4个品种为抗旱品种,辽杂10号等8个品种为中等抗旱品种,锦杂103等7个品种为干旱敏感品种,锦杂93等8个品种为高度干旱敏感品种。研究认为,相对芽长、相对根长和相对萌发抗旱指数等可以作为高粱品种抗旱性鉴定的重要指标;SOM聚类分析可作为品种抗旱性分类的重要方法。 . 采用人工气候箱内培养皿培养,PEG-6000溶液模拟干旱胁迫环境,在萌发期以80、120、150和175 g L–1PEG-6000水溶液处理31个高粱品种,旨在根据高粱品种萌发期对不同干旱胁迫程度的响应,筛选出具有抗旱能力的高粱品种并探讨高粱萌发期抗旱性鉴定的方法。通过主成分分析法(PCA)和神经网络自组织映射(SOM)聚类分析法对各高粱品种进行抗旱性综合分析与评定。PCA结果表明,相对芽长、相对根长和相对萌发抗旱指数载荷量最大,将其作为萌发期高粱抗旱性筛选的主要评价指标,并对31个高粱品种抗旱性排序。通过SOM聚类分析将31个高粱品种按抗旱性强弱分为5类,吉杂305等4个品种为高度抗旱品种,HL5等4个品种为抗旱品种,辽杂10号等8个品种为中等抗旱品种,锦杂103等7个品种为干旱敏感品种,锦杂93等8个品种为高度干旱敏感品种。研究认为,相对芽长、相对根长和相对萌发抗旱指数等可以作为高粱品种抗旱性鉴定的重要指标;SOM聚类分析可作为品种抗旱性分类的重要方法。 |
| [10] | |
| [11] | . Biophotons spontaneously emitted from radish root cells were detected using highly sensitive photomultiplier tube. Freshly isolated radish root cells exhibited spontaneous photon emission of about 402counts02s 611. Addition of hydrogen peroxide to the cells caused significant enhancement in biophoton emission to about 50002counts02s 611. Removal of molecular oxygen using glucose/glucose oxidase system and scavengering of reactive oxygen species by reducing agents such are sodium ascorbate and cysteine completely diminished biophoton emission. Spectral analysis of the hydrogen peroxide-induced biophoton emission indicates that biophotons are emitted mainly in green–red region of the spectra. The data provided by electron paramagnetic resonance spin-trapping technique showed that formation of singlet oxygen observed after addition of H 2O 2 correlates with enhancement in biophoton emission. These observations provide direct evidence that singlet oxygen is involved in biophoton emission from radish root cells. |
| [12] | . Abstract This review attempts to summarize molecular mechanisms, spectral and intensity properties, detection techniques and applications of ultra-weak photon emission. Ultra-weak photon emission is the chemiluminescence from biological systems where electronically excited species are formed during oxidative metabolic or oxidative stress processes. It is generally accepted that photons are emitted (1) at near UVA, visible, and near IR spectral ranges from 350 to 1300nm and (2) at the intensity of photon emission in the range of several units to several hundreds (oxidative metabolic process) and several hundreds to several thousands (oxidative stress process) photons s(-1)cm(-2). Current development in detection using low-noise photomultiplier tubes and imaging using highly sensitive charge coupled device cameras allows temporal and spatial visualization of oxidative metabolic or oxidative stress processes, respectively. As the phenomenon of ultra-weak photon emission reflects oxidative metabolic or oxidative stress processes, it can be widely used as a non-invasive tool for monitoring of the physiological state of biological systems. Copyright 脗漏 2014 Elsevier B.V. All rights reserved. |
| [13] | . Abstract Here we report our two applications of ultra-weak photon emission (UPE) measurements in agriculture. One is to find new types of agrochemicals that potentiate plants' defense, so-called "plant activator". We first analyzed the relation between plant defense and Elicitor-Responsive Photon Emission (ERPE) using a combination of rice cells and a chitin elicitor. Pharmacological analyses clarified that ERPE was generated as a part of the chitin elicitor-responsive defense in close relation with the generation of reactive oxygen species (ROS). Then we successfully detected the activity of plant activators as the potentiation of ERPE, and developed a new screening system for plant activators based on this principle. Another UPE application is to distinguish herbicide-resistant weeds from susceptible ones by measuring UPE in weeds. In our study, it was revealed that the weed biotypes resistant to sulfonylurea (SU) herbicides, one of the major herbicide groups, showed stronger UPE than susceptible ones after an SU herbicide treatment. By further analysis with a pharmacological and RNAi study, we found that the detoxifying enzyme P450s contributed to the UPE increase in SU herbicide resistant weeds. It is considered that weeds resistant to herbicides other than SU might also be able to be distinguished from susceptible ones by UPE measurement, as long as the herbicides are subject to detoxification by P450s. Copyright 2014 Elsevier B.V. All rights reserved. |
| [14] | . Ultra-weak photon emission originates from the relaxation of electronically excited species formed in the biological systems such as microorganisms, plants and animals including humans. Electronically excited species are formed during the oxidative metabolic processes and the oxidative stress reactions that are associated with the production of reactive oxygen species (ROS). The review attempts to overview experimental evidence on the involvement of superoxide anion radical, hydrogen peroxide, hydroxyl radical and singlet oxygen in both the spontaneous and the stress-induced ultra-weak photon emission. The oxidation of biomolecules comprising either the hydrogen abstraction by superoxide anion and hydroxyl radicals or the cycloaddition of singlet oxygen initiate a cascade of oxidative reactions that lead to the formation of electronically excited species such as triplet excited carbonyl, excited pigments and singlet oxygen. The photon emission of these electronically excited species is in the following regions of the spectrum (1) triplet excited carbonyl in the near UVA and blue–green areas (350–550nm), (2) singlet and triplet excited pigments in the green–red (550–750nm) and red-near IR (750–1000nm) areas, respectively and (3) singlet oxygen in the red (634 and 703nm) and near IR (1270nm) areas. The understanding of the role of ROS in photon emission allows us to use the spontaneous and stress-induced ultra-weak photon emission as a non-invasive tool for monitoring of the oxidative metabolic processes and the oxidative stress reactions in biological systems in vivo, respectively. |
| [15] | . Abstract Time resolved spectral components of delayed luminescence (DL) from single dry soybean seeds were measured using a device with single photon sensitivity. The seeds were aged by a thermal treatment to change their viability. A correlation was observed between the seeds viability and some DL parameters, i.e. the total number of photons emitted and the relative decay probability of excited states. This relevant result confirms the close connection between the state of biological systems and their DL, and it can allow the development of a quick selection technique for single dry seeds, a goal impossible up today. |
| [16] | . The delayed luminescence (DL) (i.e. the photo-induced photon emission long after the illumination is switched off) of unicellular green algae samples has been measured when different concentrations of heavy metals are added to the standard culture medium, with the aim of assessing the DL as a promising approach for assaying the toxicity of contaminants such as metals. In particular, samples of freshwater green micro-algae Selenastrum capricornutum have been used. Concentrations of cadmium, chromium, lead and copper, ranging from 10to 10M, have been tested. The analysis of the decay trends, in the time interval from tens of microseconds to seconds, of the DL spectral components demonstrates that the DL parameters are sensitive to the presence of such pollutants. More precisely, the performed analysis allowed us to determine phenomenological relationships between the DL parameters and the metal concentration that could be used in view of the possibility of realizing a biosensor for water pollution detection. Attempts to distinguish between different contaminants are also described. Results of this preliminary study show that the DL measure based technique is suitable as a general bioassay of metal contamination and it could also be used to test the efficiency in bioavailability studies |
| [17] | . All living organisms emit ultra-weak photon emission as a result of oxidative processes caused by the physical or chemical stress factors. In the present study, the effect of oxidative stress induced by the exposure of the plant model Arabidopsis thaliana to ultraviolet A (UVA) radiation was studied using ultra-weak photon emission. When Arabidopsis plants and leaves were exposed to UVA radiation, two kinetically distinguished phases in the ultra-weak photon emission decay were observed: the fast decay phase ( 1=0.805 0.024min) and slow decay phase ( 2=4.321 0.166min). The spectral analysis of the UVA radiation-induced ultra-weak photon emission showed that the photon emission during the fast decay phase is from both blue-green and red regions of the spectrum, whereas the photon emission during the slow decay phase is solely from the blue-green region of the spectrum. These observations reveal that triplet excited carbonyls contribute to ultra-weak photon emission during both fast and slow decay phases, whereas chlorophylls participate in the ultra-weak photon emission solely during the fast decay phase. It is illustrated here that the ultra-weak photon emission serves as a non-invasive method for the monitoring of oxidative stress in plants caused by UVA radiation. |
| [18] | . We study biophoton emission from red beans (Vigna angularis) during germination and seedling stages under drought stress. Strong photon emission is observed at the root apex when the beans are subjected to the dry condition. The spatial distribution of the emission is broader than that of emission due to the application of strong salt stress reported previously [T. Ohya et al.: Jpn. J. Appl. Phys. 39 (2000) 3696]. When they are rewatered, strong photon emission from them is again observed. As their drought damage is weaker, the intensity of the photon emission is weaker. Photon emission from damaged roots indicates their physiological response to external stress, that is, photon emission intensity measurement is useful for detecting physiological changes and evaluating the degrees of such changes before serious damage takes place without any invasion and destruction. |
| [19] | . The aim of this work was to develop a fast, optical and non-invasive method for determining the germination capacity of seeds. The relationship between delayed luminescence and germination capacity and water content and temperature, was studied in barley seeds (L.). Delayed luminescence (DL) is the photon emission from a sample directly after it has been illuminated in a dark space. DL displayed a negative correlation to the germination capacity of barley seeds, if water content and temperature were kept constant. The water content and temperature of barley seeds were also negatively correlated to DL. The mechanism of DL of barley seeds is discussed. |
| [20] | . Abstract The hypersensitive response (HR) is one mechanism of the resistance of plants to pathogen infection. It involves the generation of reactive oxygen species (ROS) which have crucial roles in signal transduction or as toxic agents leading to cell death. Often, ROS generation is accompanied by an ultraweak photon emission resulting from radical reactions that are initiated by ROS through the oxidation of living materials such as lipids, proteins, and DNA. This photon emission, referred to as 'biophotons', is extremely weak, but, based on the technique of photon counting imaging, a system has been developed to analyse the spatiotemporal properties of photon emission. Using this system, the dynamics of photon emission which might be associated with the oxidative burst, which promotes the HR, have been determined. Here, the transient generation of biophotons is demonstrated during the HR process in cowpea elicited by cucumber mosaic virus. The distinctive dynamics in spatiotemporal properties of biophoton emission during the HR expression on macroscopic and microscopic levels are also described. This study reveals the involvement of ROS generation in biophoton emission in the process of HR through the determination of the inhibitory effect of an antioxidant (Tiron) on biophoton emission. |
| [21] | . 为了将生物超弱光子技术应用于 萌发过程中小麦种子的抗旱性评价,采用聚乙二醇处理萌发的小麦种子,跟踪测量了正常和渗透胁迫下小麦种子萌发过程中的超弱光子辐射,得到了小麦种子萌发过 程中超弱光子辐射的自发发光和延迟发光信号.通过建立延迟发光动力学方程和数学拟合获得了种子萌发过程中延迟发光初始光强I(0)、衰减参量β、相干时间 τ和延迟发光积分强度I(T),根据延迟发光积分强度和自发发光的生物学意义构建了用于描述细胞状态及其有序性的状态参量和序参量.实验结果发现:小麦种 子萌发过程中,自发发光和延迟发光积分强度呈现阶跃式增长,细胞系统的状态参量和序参量也呈现阶跃式的增长,渗透胁迫抑制了这种增长,提示基于光子辐射的 细胞系统的状态参量和序参量有可能作为种子抗旱性评价的物理指标. , 为了将生物超弱光子技术应用于 萌发过程中小麦种子的抗旱性评价,采用聚乙二醇处理萌发的小麦种子,跟踪测量了正常和渗透胁迫下小麦种子萌发过程中的超弱光子辐射,得到了小麦种子萌发过 程中超弱光子辐射的自发发光和延迟发光信号.通过建立延迟发光动力学方程和数学拟合获得了种子萌发过程中延迟发光初始光强I(0)、衰减参量β、相干时间 τ和延迟发光积分强度I(T),根据延迟发光积分强度和自发发光的生物学意义构建了用于描述细胞状态及其有序性的状态参量和序参量.实验结果发现:小麦种 子萌发过程中,自发发光和延迟发光积分强度呈现阶跃式增长,细胞系统的状态参量和序参量也呈现阶跃式的增长,渗透胁迫抑制了这种增长,提示基于光子辐射的 细胞系统的状态参量和序参量有可能作为种子抗旱性评价的物理指标. |
| [22] | . 61Biophotons are related to various biological activities and processes of plant.61Biophotons can visualize the physiological states of plants in vivo.61Application of proteomics provides mechanism of plant response to biophoton emission. |
| [23] | . Evaluation of drought resistance during germination of maize is one of the basic problems that have not been solved in modern water-saving agriculture, while biological delayed luminescence technology may solve this problem. In order to explore the method of using delayed luminescence to evaluate drought resistance in maize germinating stage, the delayed luminescence of two maize varieties Wanrui No.168 and Yandan No.8 germinated in PEG-6000 solution with osmotic potential of-0.1 MPa and-0.3 MPa was measured. The results show that the integrated intensity of delayed luminescence of Wanrui No. 168 and Yandan No. 8 increase gradually with the germination process. Drought stress with osmotic potential of-0.1 MPa and-0.3 MPa inhibits the increase in integrated intensity of delayed luminescence of Wanrui No.168 and Yandan No.8, and the stronger the stress, the greater the difference of integrated intensity of delayed luminescence between two maize varieties. The relative change rate of delayed luminescence is defined as . The study found that the change of of Wanrui No. 168 and Yandan No. 8 was the same as the change of seed germination drought index and storage material transport rate under drought stress. It indicated that the size of during germination under drought stress could be used to evaluate drought resistance of maize during seed germination. . Evaluation of drought resistance during germination of maize is one of the basic problems that have not been solved in modern water-saving agriculture, while biological delayed luminescence technology may solve this problem. In order to explore the method of using delayed luminescence to evaluate drought resistance in maize germinating stage, the delayed luminescence of two maize varieties Wanrui No.168 and Yandan No.8 germinated in PEG-6000 solution with osmotic potential of-0.1 MPa and-0.3 MPa was measured. The results show that the integrated intensity of delayed luminescence of Wanrui No. 168 and Yandan No. 8 increase gradually with the germination process. Drought stress with osmotic potential of-0.1 MPa and-0.3 MPa inhibits the increase in integrated intensity of delayed luminescence of Wanrui No.168 and Yandan No.8, and the stronger the stress, the greater the difference of integrated intensity of delayed luminescence between two maize varieties. The relative change rate of delayed luminescence is defined as . The study found that the change of of Wanrui No. 168 and Yandan No. 8 was the same as the change of seed germination drought index and storage material transport rate under drought stress. It indicated that the size of during germination under drought stress could be used to evaluate drought resistance of maize during seed germination. |
| [24] | . . |
| [25] | . 为了揭示种子萌发过程中超弱光子辐射机理,分别采用蛋白质合成的转录抑制剂放线菌素D(actinomyin D,AMD)和翻译抑制剂环己亚胺酮(cycloheximide, CHM)处理萌发玉米种子,研究了玉米萌发过程中鲜质量的变化以及自发光子辐射和外界光诱导的延迟光子辐射的变化。结果表明,50 μg/mL的AMD部分抑制了萌发玉米鲜质量的增长,100 μg/mL的CHM完全抑制了萌发玉米鲜质量的增长,萌发玉米自发光子辐射强度的增长与种子鲜质量的增长呈现正相关(相关系数r分别为0.95492、0.93218和0.96235)。研究还发现,在玉米萌发过程中,AMD和CHM对延迟光子辐射的增长有不同的抑制作用,CHM部分抑制了延迟光子辐射的初始光子数、相干时间和积分强度的增大,AMD则使初始光子数、相干时间和积分强度不再增加。研究结果为揭示种子萌发过程中超弱光子辐射的机理及其技术开发提供参考。 . 为了揭示种子萌发过程中超弱光子辐射机理,分别采用蛋白质合成的转录抑制剂放线菌素D(actinomyin D,AMD)和翻译抑制剂环己亚胺酮(cycloheximide, CHM)处理萌发玉米种子,研究了玉米萌发过程中鲜质量的变化以及自发光子辐射和外界光诱导的延迟光子辐射的变化。结果表明,50 μg/mL的AMD部分抑制了萌发玉米鲜质量的增长,100 μg/mL的CHM完全抑制了萌发玉米鲜质量的增长,萌发玉米自发光子辐射强度的增长与种子鲜质量的增长呈现正相关(相关系数r分别为0.95492、0.93218和0.96235)。研究还发现,在玉米萌发过程中,AMD和CHM对延迟光子辐射的增长有不同的抑制作用,CHM部分抑制了延迟光子辐射的初始光子数、相干时间和积分强度的增大,AMD则使初始光子数、相干时间和积分强度不再增加。研究结果为揭示种子萌发过程中超弱光子辐射的机理及其技术开发提供参考。 |
| [26] | . 外磁场处理小麦种子可以提高萌发期过氧化物酶(POD)的活性;电泳图谱中,磁场透发出两条(POD)同工酶新带。在萌发初期(12h),亚胺环己酮(CHM)对外磁场提高POD活性的作用影响很小,外磁场可使POD酶蛋白的比活力增长,表明在萌发初期,磁场对种胚内的POD可能具有活化作用。萌发1d后,CHM逐渐削弱外磁场对POD活性的提高作用,而放线菌素D(AMD)则无影响。 . 外磁场处理小麦种子可以提高萌发期过氧化物酶(POD)的活性;电泳图谱中,磁场透发出两条(POD)同工酶新带。在萌发初期(12h),亚胺环己酮(CHM)对外磁场提高POD活性的作用影响很小,外磁场可使POD酶蛋白的比活力增长,表明在萌发初期,磁场对种胚内的POD可能具有活化作用。萌发1d后,CHM逐渐削弱外磁场对POD活性的提高作用,而放线菌素D(AMD)则无影响。 |
| [27] | . , |
| [28] | . We investigated the effect of benidipine, a calcium antagonist, against sodium azide (NaN(3))-induced cell death in cultured neonatal rat cardiac myocytes with increase of LDH release, depletion of cellular ATP contents, and collapse of mitochondrial membrane potential (DeltaPsi) as indicators. Cells were treated with 1 mmol/L NaN(3) for 18 h. Benidipine concentration-dependently inhibited NaN(3)-induced cell death. The protective effect of benidipine was compared with those of amlodipine, nifedipine, candesartan, and captopril. Calcium antagonists exhibited a protective effect and the IC(50) values of benidipine, amlodipine, and nifedipine were 0.65, 90, and 65 nmol/L, respectively. NaN(3)-induced cell death was inhibited completely with the calpain inhibitor. It was considered that the sustained elevation of [Ca(2+)](i) might be implicated in NaN(3)-induced cell death. Benidipine, moreover, concentration-dependently preserved cellular ATP contents and maintained DeltaPsi the extent of the control level. In conclusion, benidipine exhibited the protective effect at an approximately 100-fold lower concentration than those of amlodipine and nifedipine in the NaN(3)-induced cardiac cell death model. It was considered that both the inhibition of Ca(2+) influx and the preservation of cellular ATP contents might play an important role in the protective effect of benidipine. |
| [29] | . Sodium azide (NaN3), a well-known inhibitor of mitochondrial respiration, stimulated bud-dormancy release in grapevines similar to hydrogen cyanamide (HC), while HC, a well-known dormancy release... |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}