 ,1, 宋宇航1, 张孟寒1, 李小艳1, 李华1, 汪月霞,1,*, 齐学礼,2,*
,1, 宋宇航1, 张孟寒1, 李小艳1, 李华1, 汪月霞,1,*, 齐学礼,2,*Effects of water deficit on physiology and biochemistry of seedlings of different wheat varieties and the alleviation effect of exogenous application of 5-aminolevulinic acid
CHEN Xin-Yi,1, SONG Yu-Hang1, ZHANG Meng-Han1, LI Xiao-Yan1, LI Hua1, WANG Yue-Xia,1,*, QI Xue-Li,2,*通讯作者: *汪月霞, E-mail:yxwang2100@126.com;齐学礼, E-mail:xueliqi888@163.com
收稿日期:2021-03-12接受日期:2021-07-12网络出版日期:2022-08-03
| 基金资助: |
Received:2021-03-12Accepted:2021-07-12Published online:2022-08-03
| Fund supported: |
作者简介 About authors
E-mail:xychen3988@163.com

摘要
黄淮主产区冬小麦生育期干旱灾害发生频繁, 造成小麦苗期严重光合生理伤害。为探讨不同小麦品种光合特性对干旱胁迫的响应, 以百农207, 周麦18以及小麦新品种郑麦1860为材料, 研究了干旱胁迫对不同品种小麦幼苗光合生理、抗氧化物酶、相关基因表达水平的影响以及外源ALA的干旱缓解作用。研究结果表明, 干旱胁迫下郑麦1860具有较高的根干重和根冠比, 与周麦18相比, 抗旱能力较强的郑麦1860和百农207叶绿素含量的下降幅度、MDA含量的增加幅度、叶绿素荧光参数和光合作用参数的下降幅度相对较低, 但SOD和CAT酶活性的增加幅度相对较大。同时, 干旱胁迫显著增加了CAT、SOD-Cu/Zn、MnSOD和FeSOD抗氧化酶相关基因的转录表达水平, 且增加程度与小麦的抗旱能力密切相关。外源ALA预处理能够通过对CAT、SOD-Cu/Zn和MnSOD的转录诱导, 进一步提高干旱胁迫下SOD和CAT酶的活性, 降低膜脂过氧化损伤程度, 同时提高ATP酶的活性, 缓解干旱对小麦光合生理的伤害。此外, 本研究首次发现, 小麦叶绿体光合机构相关psb28基因转录表达的维持也与不同品种的抗旱能力有一定联系, 且受外源ALA预处理的显著诱导。本研究结果显示, 干旱胁迫下小麦抗氧化酶及叶绿体光合作用相关基因的转录表达调控, 与不同品种的抗旱能力以及外源ALA的调节作用密切相关。
关键词:
Abstract
Drought stress happens frequently in Huang-Huai winter wheat planted area, causing severe injury on photosynthetic apparatus of wheat seedlings. To characterize the traits of photosynthetic physiology in different wheat cultivars in response to water deficit, the newly in-lab cultivated Zhengmai 1860, as well as Bainong 207 and Zhoumai 18 were used as materials in this study. We explored the effects of water deficit on photosynthetic traits, antioxidant enzyme activities, and related gene transcription of seedlings in different cultivars, as well as the drought alleviation effect of exogenous application of 5-aminolevulinic acid (ALA). The results were as follows: Zhengmai 1860 had comparatively higher root dry weight and root shoot ratio than the other two cultivars under drought stress. Compared to Zhoumai 18, the drought resistant cultivars (Zhengmai 1860 and Bainong 207) had lower reduction in chlorophyll content and increased MDA content, enhanced the activities of SOD and CAT, and lowered reductions in chlorophyll fluorescence parameters and the photosynthetic parameters. Meanwhile, water deficit obviously improved the transcriptions of antioxidant enzyme-encoded genes CAT, SOD-Cu/Zn, MnSO, and FeSOD, which showed a correlation between the increasing level with the drought resistant ability. The exogenous pretreatment of ALA further enhanced the activities of SOD and CAT to lower the damage to membrane lipid peroxidation by inducing the transcriptions of CAT, SOD-Cu/Zn, and MnSOD. ATPase activity was also increased to alleviate water deficit on the damage to photosynthetic physiology. Moreover, we found for the first time that the transcriptional maintenance of chloroplast photosynthetic apparatus related psb28 gene had a correlation with the drought resistance between different wheat cultivars, which was also significantly induced by the exogenously pretreated ALA. These results in this study proposed that the transcriptions of antioxidant enzymes and chloroplast photosynthesis related genes had a close relationship with the drought resistant ability of wheat and the alleviation effect of exogenous ALA in wheat.
Keywords:
PDF (4458KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用. 作物学报, 2021, 48(2): 478-487 DOI:10.3724/SP.J.1006.2022.11026
CHEN Xin-Yi, SONG Yu-Hang, ZHANG Meng-Han, LI Xiao-Yan, LI Hua, WANG Yue-Xia, QI Xue-Li.
小麦(Triticum aestivum)是我国第二大粮食作物, 也是我国北方最主要的粮食作物, 其产量的丰欠直接影响人民生活水平的提高和国民经济的发展。黄淮主产区冬小麦生育期干旱灾害发生频繁, 随着全球气候的变化, 干旱胁迫导致小麦植株生理伤害, 严重影响小麦产量[1], 培育抗旱节水小麦品种是保障我国粮食安全的重要途径之一[2]。
植物叶绿体光合系统对环境胁迫最为敏感, 其中干旱是抑制光合系统功能并且降低作物产量最为严重的逆境胁迫之一[3]。植物在干旱逆境下维持光合机构正常运转, 提供充足的光合产物, 叶绿体的光合功能必须能够得到足够的保护, 光合功能的修复能力也已经成为小麦抗旱品种选育的重要指标[4,5]。光合过程中的ATP酶, 通过类囊体膜传递质子, 催化光合磷酸化、在细胞生物代谢方面发挥着重要作用[6]。
植物PSII是承担光合作用中水分氧化和醌还原的多亚基色素-蛋白质复合物, 而Psb28蛋白(PSII蛋白28)是PSII复合物中一种大小约为13.5 kD的具有重要光合功能的可溶性蛋白[7]。集胞藻psb28基因的突变导致在高温下生长迟缓, PSII光损伤修复速率下降, 且在PSII修复中Psb28-1通过与CP43-less单体的结合来发挥重要作用[8]。在水稻中, 强光照射可诱导psb28基因的转录表达[9]。然而, 有关小麦psb28基因在干旱胁迫下PSII修复中的表达调节规律, 目前尚未有报道。
SOD和CAT等是植物体内负责清除活性氧的重要保护酶, 其活性高低与作物抗逆性密切相关[10]。植物SOD酶通常以多种形式存在, 如SOD-Cu/Zn、FeSOD、MnSOD等[11]。在小麦中这些基因过表达后, 植株的抗旱性明显增强, 说明这些基因的表达调控对于植物的抗旱性具有重要作用[12]。
生长调节物质在植物抵抗环境胁迫过程中具有重要的作用, 因而在作物抗逆栽培过程中具有广阔的应用前景[13]。5-氨基乙酰丙酸(5-aminolevulinic acid, ALA)是植物叶绿素中卟啉类化合物合成的前体, 在低浓度条件下能够对胁迫环境下生长的植物表现出生长调节特性。在干旱胁迫下, 外源ALA可以缓解小麦等多种植物的干旱损伤[14]。然而, 虽然已有研究表明, 外源ALA可以提高植物的光合能力, 从而缓解逆境损伤[15], 但从基因表达调控的角度, 探究外源ALA对作物光合作用的保护机制目前尚未有报道。
郑麦1860是河南省农业科学院小麦研究所分子育种团队最新育成的绿色超高产小麦新品种, 具有超高产, 对氮肥和磷肥具有高效利用的特性, 同时具有良好的综合抗病性。鉴于干旱是黄淮冬麦区主要的产量影响逆境因子, 本研究以郑麦1860为材料, 与耐旱性突出的百农207品种以及抗旱性较弱的周麦18作比较, 进一步分析其抗旱能力、分子机制以及外源ALA的干旱缓解机制, 以期为小麦的抗旱育种和栽培技术构建提供参考。
1 材料与方法
1.1 供试材料
供试材料为小麦品种郑麦1860、百农207以及周麦18, 均来自河南省农业科学院小麦研究所, 其中郑麦1860为该所自主选育。挑选大小一致, 无虫咬, 颗粒饱满的小麦种子, 用5%的H2O2消毒5 min, 流动的无菌水冲洗8~10次。于23℃暗中浸泡10 h。将吸胀的种子按照腹缝线朝下, 胚芽鞘置于同一方向, 均匀地平铺于放置了两层浸湿滤纸的培养皿中, 于25℃恒温培养箱中培养。1.2 干旱处理
待小麦幼苗培养3 d, 催芽至1 cm, 挑选长势一致的幼苗移植于河南省农业科学院现代农业科技试验示范基地, 每个品种设3个重复, 处理之前统一进行常规田间水肥管理。待各品种生长至三叶一心时取长势相近的随机分为3组: 对照组(CK)、干旱组(Drought)和干旱+ALA组(D+A), 每组处理为六小盆小麦(54株), 作为1次重复, 各处理重复3次以上。D+A组在干旱处理前进行100 mg L-1 ALA喷洒处理[15], CK进行蒸馏水喷洒, 连续喷施3 d后, Drought组、D+A组停止浇水来模拟干旱胁迫, CK组正常浇水处理, 当Drought组、D+A组小麦土壤含水量达到20%, 表型差异明显时(处理6 d), 于上午9:00取样并进行后续测定。1.3 测定项目与方法
1.3.1 植株干物质量 选取长势一致的小麦幼苗, 将植株分为地上部(茎叶)和地下部(根系)两部分, 105℃杀青15 min, 后进行85℃恒温烘干至质量不再变化, 进行称重, 再计算根冠比, 每组4次重复。1.3.2 叶绿素含量 称取100 mg左右叶片组织, 剪碎后浸泡于10 mL丙酮乙醇混合提取液中(按体积配比, 乙醇、丙酮、水的比例为4.5∶4.5∶1.0)。将样品放在黑暗下提取18 h, 分别在663 nm和645 nm波长下测量吸光值, 并计算叶绿素含量。
1.3.3 丙二醛含量 称取100 mg的叶片, 用液氮预冷的研钵冷冻研磨后, 采用硫代巴比妥酸法测定丙二醛的含量[16]。
1.3.4 SOD和CAT酶活性 参照Wang等[17]的方法, 1 g小麦叶片加入8 mL的酶提取液并进行酶蛋白的提取, 超氧化物歧化酶活性(SOD)用氮蓝四唑光化还原法进行, 过氧化氢酶(CAT)活性采用过氧化氢还原法进行。
1.3.5叶绿素荧光参数 使用叶绿素荧光仪 MINI- PAM-II (Zeaquest Scientific Technology Co., Ltd.), 参照说明书测定以旗叶叶片中心点处的叶绿素荧光参数的光反应曲线, 计算出Fv/Fm和qP。
1.3.6 光合气体交换参数 净光合速率(Pn)和气孔导度(gs)采用LI-6400光合仪进行, 参照Wang等[15]的方法, 光源设定在1000 µmol m-2 s-1的PPFD, 参照CO2浓度为400 µmol mol-1。
1.3.7 ATP酶活性 使用索莱宝公司Ca2+ Mg2+- ATP酶活性检测试剂盒, 按照试剂盒说明书方法进行测定。
1.3.8 总RNA的提取和基因表达水平的测定
采用OminiPlant RNA Kit (DNase I)试剂盒提取小麦总RNA, 并采用超微量核酸分析仪检测总RNA的浓度和纯度。采取HiScript II Q RT SuperMix for qPCR试剂盒进行逆转录第一条cDNA链的合成, 采用Primer5.0设计相关引物(http://frodo.wi.mit.edu/ cgi-bin/primer5/primer5_www.cgi), 由尚亚生物技术公司合成(表1)。实时荧光定量PCR采用ChamQ Universal SYBR qPCR Master Mix试剂盒和Bio-Rad iQ5实时荧光定量PCR仪, 并参照说明书进行。每个处理各做3次生物学重复, 用Actin作为小麦内参基因, 计算各个处理得到的Ct值, 采用2-ΔΔCt法来分别计算各基因的相对表达量[18]。
Table1
表1
表1实时荧光定量PCR引物序列
Table1
| 基因名称 Gene name | 基因功能 Gene function | 引物序列 Primer sequence (5′-3′) | 参考文献 Reference |
|---|---|---|---|
| SOD-Cu/Zn | 叶绿体Cu和Zn超氧化物歧化酶 Chloroplast Cu/Zn superoxide dismutase | F: CGCTCAGAGCCTCCTCTTT R: CTCCTGGGGTGGAGACAAT | [19] |
| FeSOD | 叶绿体Fe超氧化物歧化酶 Chloroplast Fe superoxide dismutase | F: GTCCTACTACGGCCTCACCA R: ACGTAGTCCTGCTGGTGCTT | [20] |
| MnSOD | 线粒体Mn超氧化物歧化酶 Mitochondrion Mn superoxide dismutase | F: CAGAGGGTGCTGCTTTACAA R: GGTCACAAGAGGGTCCTGAT | [20] |
| CAT | 过氧化氢酶 Catalase | F: CCATGAGATCAAGGCCATCT R: ATCTTACATGCTCGGCTTGG | [19] |
| psb28 | 叶绿体Psb28蛋白 Chloroplast Psb28 protein | F: AACTGTCGAGCTGGTAACGG R: ACCGGTTTTCCTCAGTTCGT | 本研究设计 Design in this study |
| Actin | β-actin | F: GGAATCCATGAGACCACCTAC R: GACCCAGACAACTCGCAAC | [21] |
新窗口打开|下载CSV
1.4 数据分析
所有的实验数据利用Microsoft Excel软件来进行作图和分析, 结果用平均值±标准差(mean ± SD)来表示, 并用单因素方差分析(ANOVA)方法比较小麦各处理之间的差异显著性(P < 0.05)。2 结果与分析
2.1 干旱胁迫对不同品种小麦幼苗生长的影响
干旱是黄淮栽培区小麦幼苗期经常遇到的逆境胁迫, 本研究采用的3个品种小麦, 在干旱胁迫6 d后, 小麦幼苗的生长均受到显著影响, 叶片表现出明显的下垂, 新叶停止生长。与其他品种相比, 周麦18幼苗干旱胁迫症状更为明显, 叶片下垂的程度更为严重。与干旱处理相比, 外源ALA预处理后, 小麦幼苗在干旱处理第6 d生长状况较好, 其中郑麦1860几乎无叶片因干旱而发生下垂的现象(图1)。本形态观察结果表明, 本供试3个小麦品种中, 百农207和郑麦1860小麦幼苗均具有较强的干旱胁迫耐受能力, 周麦18耐旱性相对较弱; 外源ALA预处理能够明显缓解干旱对小麦的损伤。如图1所示, 在小麦遭受干旱胁迫后, 百农207和周麦18的根干重和茎干重均显著下降(P < 0.05), 但根冠比变化不显著(P > 0.05)。外源ALA处理后根干重和茎干重的下降趋势得到缓解, 尤其是百农207的根冠比显著上升(P < 0.05)。然而, 郑麦1860品种根干重在干旱胁迫下显著上升(P < 0.05), 虽茎干重无显著变化(P > 0.05), 但根冠比显著提高67% (P < 0.05)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1干旱及外源ALA处理对不同抗旱性小麦外观形态和生物量的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 1Effects of drought stress and exogenous ALA on the appearance and biomass of different drought-resistant wheat cultivar
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: pretreatment with ALA prior to drought stress.
2.2 干旱胁迫对小麦幼苗叶绿素、MDA含量以及SOD和CAT活性的影响
如图2所示, 在干旱胁迫下, 百农207和郑麦1860小麦幼苗叶片叶绿素含量分别下降36%和32%, 而周麦18下降了56%。虽然外源ALA处理可以恢复干旱胁迫下叶绿素的含量, 但周麦18仍比对照低38%。干旱胁迫显著提高了叶片MDA的含量(P < 0.05), 在3个品种中, 外源ALA处理可降低小麦幼苗叶片在干旱胁迫下的MDA含量, 缓解叶片氧化损伤, 由图2可知周麦18 MDA含量上升最为明显, 且在外源ALA预处理后, 与其他2个小麦品种相比, 高于对照的程度仍最为显著。干旱胁迫下, 百农207和郑麦1860小麦叶片SOD和CAT酶活性也显著升高(P < 0.05), 且外源ALA处理进一步提高两种酶活性。虽然周麦18小麦叶片SOD酶活性在干旱胁迫和外源ALA处理下也显著升高, 但升高程度明显低于其他2个小麦品种, 其中CAT酶活性在干旱胁迫下虽有提高, 但并不显著(P > 0.05)。图2
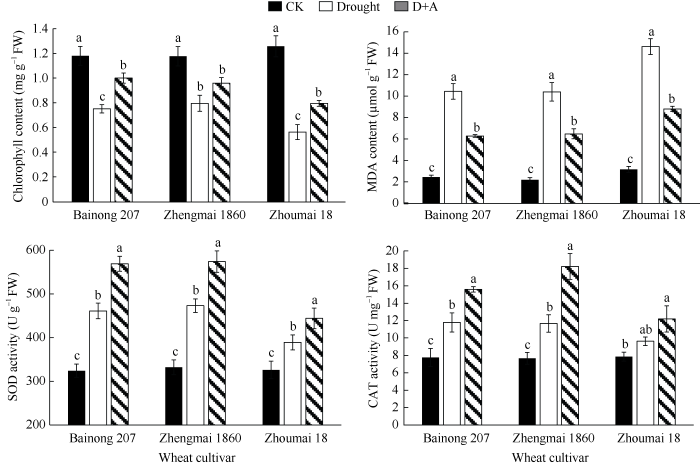 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2干旱及外源ALA处理对不同抗旱性小麦叶绿素、MDA含量以及抗氧化酶活性的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 2Effects of drought stress and exogenous ALA on the chlorophyll and MDA contents, and the activities of antioxidant enzymes for different drought-resistant wheat cultivars
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: exogenous ALA pretreatment prior to water deficit treatment.
2.3 干旱胁迫对不同品种小麦幼苗叶绿素荧光参数和气体交换的影响
如图3所示, 干旱胁迫下, 3个小麦品种的光合作用相关指标Fv/Fm、qP、Pn和gs值均显著降低(P < 0.05), 而外源ALA处理后, 所测干旱胁迫下的光合作用指标均显著高于未经ALA处理的小麦幼苗。在这3个小麦品种中, 周麦18幼苗叶片的光合作用指标下降最为显著, 且经外源ALA预处理后的改善程度也较其他2个品种低, 说明ALA能有效缓解干旱胁迫对小麦幼苗造成的光合损伤。图3
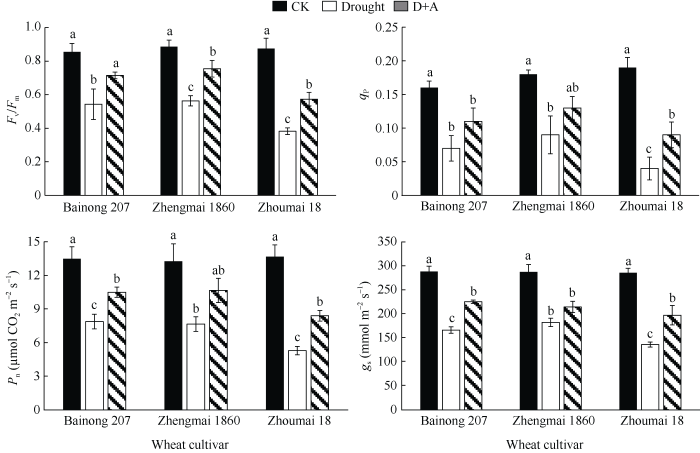 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3干旱及外源ALA处理对不同抗旱性小麦叶绿素荧光参数和光合参数的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 3Effects of drought stress and exogenous ALA on the chlorophyll fluorescence parameters and the photosynthetic parameters of different drought-resistant wheat cultivar
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: exogenous ALA pretreatment prior to water deficit treatment.
2.4 干旱胁迫对不同品种小麦幼苗ATP酶活性的影响
光合作用中线性电子传输产生ATP, ATP酶活性决定后续碳的固定[22]。由图4可知, 供试3个小麦品种的ATP酶活性在干旱胁迫下均显著下降(P < 0.05)并低于对照(正常水分处理), 其中郑麦1860的下降程度最小, 并明显高于干旱胁迫条件下的百农207和周麦18; 且在外源ALA预处理后, 在干旱胁迫下仅比对照低10%, 说明100 mg L-1的5-ALA预处理可明显缓解干旱胁迫下小麦幼苗叶片中ATP酶活的降低(图4)。图4
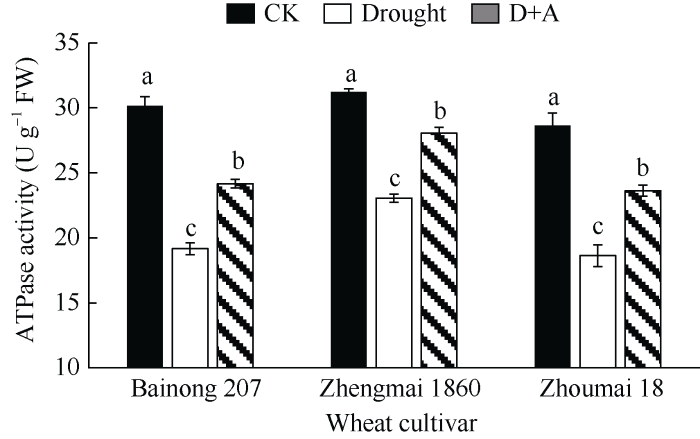 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4干旱及外源ALA处理对不同抗旱性小麦ATP酶活性的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 4Effects of drought stress and exogenous ALA on ATPase activity of different drought-resistant wheat cultivar
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: exogenous ALA pretreatment prior to water deficit treatment.
2.5 干旱胁迫对不同品种小麦幼苗抗氧化酶编码基因表达水平的影响
本研究测定了CAT、SOD-Cu/Zn、MnSOD和FeSOD等4种抗氧化酶相关基因表达水平的变化, 结果如图5所示, 除周麦18的MnSOD基因转录水平变化不显著之外, 其他小麦品种的抗氧化物酶基因在干旱胁迫下均显著上调(P < 0.05), 且百农207和郑麦1860品种的上调程度明显高于周麦18品种。小麦幼苗经外源ALA处理后, 在干旱胁迫下除FeSOD基因的转录水平无明显变化, CAT、SOD-Cu/Zn和MnSOD基因转录均显著上调(P < 0.05)。图5
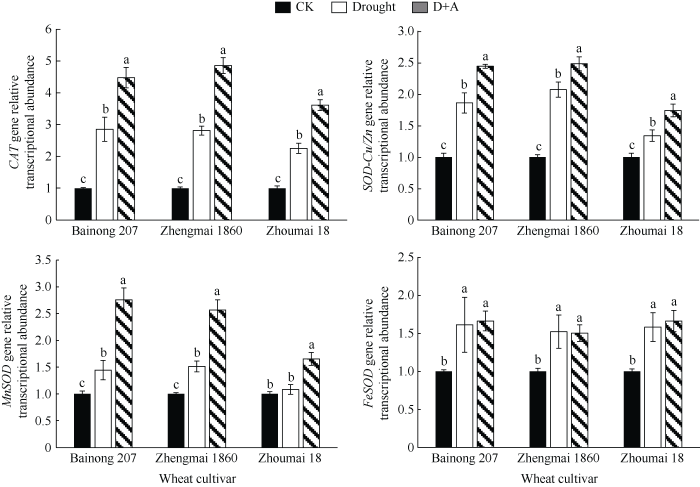 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5干旱及外源ALA处理对不同抗旱性小麦抗氧化物酶基因转录表达的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 5Effects of drought stress and exogenous ALA on the transcriptions of antioxidant enzyme-related genes of different drought-resistant wheat cultivar
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: exogenous ALA pretreatment prior to water deficit treatment.
2.6 干旱胁迫对不同品种小麦幼苗叶绿体基因psb28转录表达的影响
在干旱胁迫下, 百农207和郑麦1860小麦幼苗的psb28基因的转录表达水平虽有降低, 但并不显著(P > 0.05), 而周麦18小麦幼苗下降程度为52%。经外源ALA预处理后, 3个供试小麦品种幼苗的psb28基因转录水平均显著上升, 但周麦18的上升程度最低。表明干旱胁迫明显降低了叶绿体psb28基因的转录, 外源ALA能显著提高干旱胁迫下小麦幼苗中psb28基因的转录表达。图6
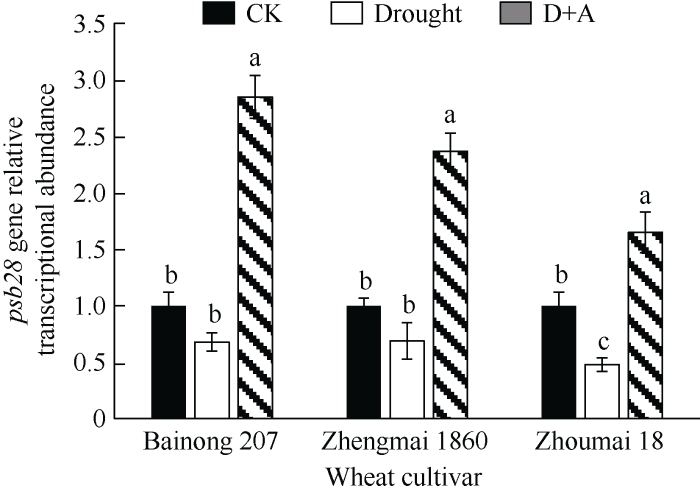 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6干旱及外源ALA处理对不同抗旱性小麦叶绿体psb28基因表达的影响
数据为3次重复的平均值±标准差, 图中不同字母表示品种内处理间有显著差异(P < 0.05)。CK: 正常供水; Drought: 干旱处理; D+A: 干旱处理之前用外源ALA预处理。
Fig. 6Effects of drought stress and exogenous ALA on the transcription of chloroplast psb28 gene in different drought- resistant wheat cultivar
Data are means ± standard deviations of three replicates. Different letters indicate significant difference among treatments within each cultivar at P < 0.05. CK: normal water supply; Drought: water deficit treatment; D+A: exogenous ALA pretreatment prior to water deficit treatment.
3 讨论
植物生物量生产与分配是适应干旱等环境胁迫的重要策略之一, 在不同逆境胁迫下可能采取不同的生产与分配方式[23]。植物的地上部分与地下部分, 是植物进行光合作用和养分吸收的重要组成部分, 而干物质的累积是植物进行光合作用的直接产物, 在小麦乃至其他农作物的产量形成有重要作用[24]。有研究表明, 抗旱性强的品种在干旱胁迫下的根系相对更为发达, 可以通过合成渗透调节物质, 使根内外水势差减小, 根细胞失水减少, 甚至抑制地上部分的伸长及生长, 为后期的养分调节及补充提供有利条件, 进一步抵御外来的干旱胁迫[25]。本研究结果表明, 干旱胁迫下郑麦1860根干重显著升高, 茎干重无明显变化, 因而根冠比显著增加, 可更多地吸收和利用土壤中的水分, 并提高气孔导度[26,27], 这可能是郑麦1860抗旱能力较强的重要原因。干旱胁迫对植物的生长生理造成显著影响, 降低叶绿素含量, 并造成脂质过氧化等组织损伤[28]。MDA是脂质过氧化的终产物, 其含量直接反映植物细胞膜脂过氧化的程度, 植物在干旱胁迫下需要增强抗氧化能力以维持活性氧(ROS)产生与清除之间的平衡, 这种能力也是植物抗旱性能力的重要体现[29,30]。较高的ATP酶活性可以促进光合产物的合成, 提高光合速率[22]。在本研究中, 干旱可显著促进SOD-Cu/Zn、FeSOD、MnSOD以及CAT基因的转录表达, 从而增强SOD和CAT酶的活性, 其中, 与周麦18品种相比, 抗旱能力较强的百农207和郑麦1860品种的增强幅度更大, 但这3个品种的MDA含量仍显著高于各自的对照水平, 说明干旱胁迫可通过促进抗氧化物酶相关基因的转录表达, 提高抗氧化物酶活性, 但这种促进作用并不足以维持活性氧代谢的平衡[31]。外源ALA处理可以进一步提高SOD-Cu/Zn、MnSOD和CAT基因的转录表达, 增加SOD和CAT酶的活性, MDA含量也进一步降低, 但本研究的结果发现, 外源ALA的这种保护促进作用与叶绿体FeSOD的转录诱导并无明显关系。MnSOD是线粒体中重要的抗氧化酶, 能够歧化超氧阴离子(O2?)形成过氧化氢(H2O2), 维持ROS的代谢平衡。Mn2+作为植物细胞中氧化还原、电子传递等过程密切相关的元素, 是叶绿体中光合放氧复合体的重要组成, 对小麦光合作用、呼吸作用和叶绿素含量都有影响, 可作为供体将电子传递给叶绿素反应中心, 使叶绿体膜中的脂粒免除过氧化的危害, 避免光合能力遭受干旱胁迫的过度破坏[32]。本研究结果说明外源ALA喷施处理可显著促进干旱对抗氧化物酶基因的转录诱导, 提高酶促防御水平, 从而降低膜脂过氧化损伤程度, 缓解干旱对小麦光合生理的伤害。
干旱胁迫能显著减少叶绿体电子传递活动, 降低光合速率, 这是因为水分的匮乏可以促进植物气孔的关闭, 减少CO2的进入, 造成胞内CO2浓度的降低和碳固定的减少, 从而导致PSII光催化和光合作用对电子需求的失衡[33]。在本研究中, 干旱胁迫降低小麦幼苗叶绿素含量和叶绿素荧光动力学参数Fv/Fm和qP, 该参数反映光合电子传递状态[34], 同时降低光合作用参数Pn和gs值。在空气与植物交界面, 叶片气孔为光合作用碳固定提供了CO2进入的通道, 同时阻止水分的过度散失, Pn值体现了叶绿体放氧复合体的功能状态[35], 而gs值的降低说明气孔关闭是Pn值降低和PSII化学活性在干旱胁迫下降低的重要因素。抗旱能力较弱的周麦18的叶绿素荧光动力学参数和光合作用参数, 在干旱胁迫下的降低程度更明显, 抗旱能力强的郑麦1860的ATP酶活性也能够在干旱胁迫下维持较高水平, 说明ATP酶活性的强弱也与小麦的抗旱能力密切相关, 这与干旱胁迫下的大麦中的研究结果相一致[36], 进一步证明光合作用的维持能力与不同小麦品种之间的抗旱能力差异具有明显关联。
外源ALA可以显著缓解干旱胁迫造成的小麦叶绿素荧光动力学参数和光合作用参数的降低, 这与干旱胁迫下的油菜中的研究结果类似[37]。ALA的这种保护作用可能是由于其是植物叶绿素生物合成的第一关键前体物, 本研究结果也证实, 外源ALA的补充可以促进干旱胁迫下叶绿素的快速合成[38]。充足的叶绿素对于干旱胁迫下PSII的功能修复具有重要作用, 这是因为PSII的核心复合物与一系列叶绿素-结合蛋白相链接, 这些蛋白作为光捕获天线, 利用光能驱动氧化还原反应催化水分氧化质体醌的还原[39]。本研究结果与在肯塔基蓝草中的研究类似, 说明外源ALA可削弱胁迫诱导产生的光抑制, 从而增强逆境下的光合作用[40]。
Psb28是与细胞色素b559结合并参与植物叶绿体CP47色素蛋白合成的重要蛋白[41], psb28基因的表达调控在逆境胁迫下PSII的功能修复中具有重要作用[42]。本研究首次揭示了干旱胁迫显著抑制了小麦psb28基因的转录表达, 而且这种抑制作用的强弱也与苗期小麦的抗旱性密切相关, 外源ALA对psb28基因转录的诱导可能也是干旱胁迫下光合功能的维持有关, 因为较高水平的psb28基因表达水平有助于CP47色素蛋白的生物合成, 以更好地吸收和转移光能至光合作用活性中心, 从而促进光合速率[42]。
4 结论
干旱胁迫对小麦生长和光合生理造成显著伤害, 与周麦18相比, 抗旱能力较强的郑麦1860和百农207具有较低叶绿素含量下降值和MDA含量增加值, 较高的SOD和CAT酶活性增加值, 叶绿素荧光参数和光合作用参数的下降值也相对较低。同时, 干旱胁迫显著增加了CAT、SOD-Cu/Zn、MnSOD和FeSOD抗氧化保护酶相关基因的转录表达水平, 且增加程度与小麦的抗旱能力密切相关。外源ALA预处理能够通过对抗氧化物酶基因的转录诱导, 进一步降低膜脂过氧化损伤程度, 缓解干旱对小麦光合生理的伤害。此外, 本研究首次发现, 小麦叶绿体光合机构相关psb28基因转录表达的维持也与不同品种的抗旱能力有一定联系, 且受外源ALA预处理的显著诱导。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}