 ,, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑,*, 姜东,*南京农业大学农学院 / 农业部作物生理生态与生产管理重点实验室, 江苏南京 210095
,, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑,*, 姜东,*南京农业大学农学院 / 农业部作物生理生态与生产管理重点实验室, 江苏南京 210095Physiological mechanisms of pre-anthesis waterlogging priming on waterlogging stress tolerance under post-anthesis in wheat
MA Bo-Wen,, LI Qing, CAI Jian, ZHOU Qin, HUANG Mei, DAI Ting-Bo, WANG Xiao,*, JIANG Dong,*College Agronomy, Nanjing Agricultural University / Key Laboratory of Crop Physiology Ecology and Production Management of Ministry of Agriculture, Nanjing 210095, Jiangsu, China通讯作者: *王笑, E-mail:xiaowang@njau.edu.cn;姜东, E-mail:jiangd@njau.edu.cn, Tel: 025-84399627
收稿日期:2021-01-10接受日期:2021-04-14网络出版日期:2021-06-02
| 基金资助: |
Corresponding authors: E-mail:xiaowang@njau.edu.cn;E-mail:jiangd@njau.edu.cn, Tel: 025-84399627
Received:2021-01-10Accepted:2021-04-14Published online:2021-06-02
| Fund supported: |
作者简介 About authors
E-mail:2017101016@njau.edu.cn,Tel:025-84399623

摘要
采用对渍水胁迫和渍水锻炼响应差异的小麦品种为材料, 在四叶一心期和六叶一心期分别进行渍水锻炼2 d; 在开花后7 d进行渍水胁迫5 d, 分析不同小麦品种对渍水胁迫响应的差异及其生理机制。结果表明, 花后渍水胁迫显著降低旗叶叶绿素含量(SPAD)和实际光化学效率(ΦPSII), 抑制花后光合同化物积累(PAA), 导致籽粒千粒重和产量降低; 与不耐渍型品种相比, 耐渍型品种在渍水胁迫下可维持较高的SPAD、ΦPSII和PAA, 超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)以及谷胱甘肽还原酶(GR)酶活性提高, 过氧化氢(H2O2)、超氧阴离子自由基(O2?)和丙二醛(MDA)含量较低。与花前未进行渍水锻炼和花后渍水处理(NW)相比, 花前渍水锻炼和花后渍水处理(PW)下, 渍水锻炼敏感型品种较渍水锻炼不敏感品种显著提高了花后渍水胁迫下小麦旗叶SPAD (8.8%)和ΦPSII (17.6%)、降低非调节性能量耗散ΦNO (10.7%)和调节性能量耗散ΦNPQ (16.5%), 提升SOD (15.8%)、CAT (17.8%)、APX (8.9%)以及GR (30.7%)酶活性, 增加了叶片可溶性糖(17.5%)和蔗糖含量(21.6%), 促进花前贮藏物质向籽粒的转运率REP (20.0%), 同步提升PAA (10.8%)。与不耐渍型品种相比, 耐渍型小麦品种在花后渍水胁迫下旗叶的光合能力、抗氧化能力和干物质向籽粒的转运能力更强。花前渍水锻炼提高了各品种小麦花后渍水胁迫下旗叶的光合能力、抗氧化能力和干物质向籽粒的转运能力, 增强了小麦耐渍性; 与渍水锻炼不敏感型品种相比, 渍水锻炼敏感型品种的光合能力和抗氧化酶活性增幅较大。
关键词:
Abstract
In order to investigate the responses and mechanisms of different wheat varieties to waterlogging stress and waterlogging priming, waterlogging priming was conducted for two days at the four-leaf and six-leaf stages, respectively, and waterlogging stress was performed for five days at post-anthesis using wheat varieties with different responses to waterlogging stress and waterlogging priming as experimental materials. Results showed that waterlogging stress significantly reduced chlorophyll content (SPAD) and actual photochemical efficiency (ΦPSII), inhibited the accumulation of post-anthesis photosynthetic assimilation accumulation (PAA), decreased kernel weight and grain yield. Compared with the waterlogging-sensitive varieties, the waterlogging-tolerance varieties could maintain higher SPAD, ΦPSII and PAA, and higher activities of superoxide dismutase (SOD), Catalase (CAT), ascorbate peroxidase (APX) and glutathione reductase (GR), lower contents of H2O2, O2?production rate and malondialdehyde (MDA) under post-anthesis waterlogging stress. Compared with non-primed plants, primed plants could maintain higher chlorophyll fluorescence performance and higher activities of antioxidant enzymes. Compared with the waterlogging priming-insensitive varieties, the priming-sensitive varieties increased SPAD (8.8%) and ΦPSII (17.6%), decreased the non-regulated energy dissipation ΦNO (10.7%) and the regulation energy dissipation ΦNPQ (16.5%), increased the activities of SOD (15.8%), CAT (17.8%), APX (8.9%) and GR (30.7%), increased the contents of total soluble sugar (17.5%) and sucrose (21.6%), increased remobilization efficiency of pre-anthesis stored dry matter (REP, 20.0%) and PAA (10.8%). The waterlogging tolerant varieties could maintain higher photosynthesis rate, dry matter translocation capacity and activities of antioxidant enzymes. Compared with waterlogging sensitive varieties, the increase amplitude of photosynthetic ability and antioxidant enzyme activity of priming-sensitive cultivars was higher under waterlogging stress.
Keywords:
PDF (1344KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
马博闻, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑, 姜东. 花前渍水锻炼调控花后小麦耐渍性的生理机制研究. 作物学报, 2022, 48(1): 151-164 DOI:10.3724/SP.J.1006.2022.11005
MA Bo-Wen, LI Qing, CAI Jian, ZHOU Qin, HUANG Mei, DAI Ting-Bo, WANG Xiao, JIANG Dong.
降雨量偏多、灌溉过量或土壤排水不良等问题, 致使全球约10%的种植区域, 小麦种植总面积的25%受到土壤渍害的影响[1]。长江中下游地区是我国小麦主产区之一, 该区域降水时空分布不规律, 降雨量集中, 强度大, 水位经常超过小麦需求而导致田间涝渍, 涝渍灾害严重制约着该区域小麦的稳产、高产[2,3]。逆境锻炼[4]是指在生育前期对植物进行适度非生物逆境预处理, 可增强植株对生育后期再次发生的同一或不同逆境胁迫的抗性[5]。越来越多的研究表明, 逆境锻炼可实现植株主动抗逆, 是提高作物对非生物逆境耐性的重要途径, 已有研究表明, 不同小麦品种对干旱锻炼的响应存在着显著的形态差异[6], 然而不同小麦品种对渍水锻炼的响应评价还相对较少。涝渍胁迫下, 根系因供氧不足, 加速衰老, 进而影响养分的吸收导致叶片黄化, 叶片光合作用等生理生化过程受阻, 光合产物向籽粒的转运降低, 最终导致产量的降低[7,8]。前期研究表明, 与未锻炼过的植株相比, 花前渍水锻炼显著提高了小麦花后渍水胁迫下旗叶光合能力和抗氧化能力, 缓解重度渍水胁迫对植株造成的损伤[9]。通过蛋白质组学分析发现, 与能量代谢和应激防御相关的关键酶在渍水锻炼处理中的表达显著高于未锻炼处理, 如参与卡尔文循环的关键酶Rubisco酶和抗坏血酸过氧化物酶[10]。前人研究多集中于比较单个或几个耐渍程度不同的品种在形态和生理生化指标上的差异, 但不同品种对渍水锻炼的响应差异少有研究, 探究不同小麦品种对渍水锻炼的响应差异能更好的阐明小麦耐渍形成的生理机制, 可为小麦耐渍育种提供理论依据及抗渍栽培技术提供支持。本研究选用对渍水胁迫及渍水锻炼响应存在差异的16个小麦品种, 通过分析花后渍水胁迫下不同小麦品种产量及其构成因素、叶片叶绿素荧光参数、干物质积累与转运、渗透调节能力及抗氧化能力等, 以期阐明不同小麦品种对渍水锻炼的响应及小麦耐渍性形成的生理机制。
1 材料与方法
1.1 试验材料与设计
试验于2018—2019年(以下简称2019年)和2019—2020年(以下简称2020年)在南京农业大学白马基地进行盆栽种植。试验用土属黄棕壤, 养分状况见表1。土壤经自然风干后过筛与肥料混匀, 称取7.5 kg装入高25 cm, 直径24 cm的聚乙烯塑料桶, 浇水沉实后播种。每桶施用全N 1.2 g, P2O5 0.36 g和K2O 0.9 g, 两年氮素基追比为5∶5, 追肥于拔节期施用。精选饱满种子播种每盆14粒, 于三叶一心间苗, 留苗每盆7株。Table 1
表1
表1小麦播种前土壤基础地力
Table 1
| 年份 Year | 有机质 Organic matter (g kg-1) | 全氮 Total nitrogen (g kg-1) | 速效氮 Available nitrogen (mg kg-1) | 速效磷 Available phosphorus (mg kg-1) | 速效钾 Available potassium (mg kg-1) |
|---|---|---|---|---|---|
| 2018-2019 | 17.63 | 1.02 | 28.03 | 18.90 | 130.66 |
| 2019-2020 | 16.34 | 0.99 | 30.95 | 23.02 | 137.46 |
新窗口打开|下载CSV
渍水锻炼处理在四叶一心期和六叶一心期进行, 保持1~2 cm水层处理2 d; 渍水胁迫在花后7 d进行, 保持1~2 cm水层处理5 d。对照在整个生育期保持田间持水量的70%~80%。试验共计3个处理: 对照处理(CK)、花前渍水锻炼和花后渍水处理(PW)、花前未进行渍水锻炼和花后渍水处理(NW)。试验为随机区组设计, 每处理3个重复, 每重复种植2盆, 共计288盆。
通过2017—2018年和2018—2019年两年度以157个小麦品种为材料, 在盆栽条件下研究渍水胁迫时不同小麦品种对渍水胁迫和渍水锻炼响应的差异, 通过测定产量、千粒重、胁迫后生物量、收获指数、干物质转运与积累、叶片叶绿素荧光参数和叶绿素含量。采用渍水指数WI= NW/CK, 通过主成分分析法评价不同品种耐渍性的差异。同时采用锻炼指数WPI=经过锻炼处理(PW)/NW, 通过主成分分析法评价不同品种对渍水锻炼的响应差异。最后筛选出对渍水胁迫和渍水锻炼敏感性存在差异的品种组合共计16个, 作为本研究材料(表2)。
Table 2
表2
表2供试品种
Table 2
| 编号 No. | 品种名称 Variety | 类型 Type | 编号 No. | 品种名称 Variety | 类型 Type |
|---|---|---|---|---|---|
| 1 | 淮麦22 Huaimai 22 | 锻炼敏感且耐渍 ST | 9 | 师栾02-1 Shiluan 02-1 | 锻炼不敏感且耐渍 IT |
| 2 | 齐大195 Qida 195 | 锻炼敏感且耐渍 ST | 10 | 汶农17 Wennong 17 | 锻炼不敏感且耐渍 IT |
| 3 | 扬麦20 Yangmai 20 | 锻炼敏感且耐渍 ST | 11 | 周麦27 Zhoumai 27 | 锻炼不敏感且耐渍 IT |
| 4 | 镇麦10 Zhenmai 10 | 锻炼敏感且耐渍 ST | 12 | 扬麦9号 Yangmai 9 | 锻炼不敏感且耐渍 IT |
| 5 | 衡4399 Heng 4399 | 锻炼敏感且不耐渍 SI | 13 | 济麦22 Jimai 22 | 锻炼不敏感且不耐渍 II |
| 6 | 京冬22 Jingdong 22 | 锻炼敏感且不耐渍 SI | 14 | 济南矮6号 Jinanai 6 | 锻炼不敏感且不耐渍 II |
| 7 | 鲁垦麦9号 Lukenmai 9 | 锻炼敏感且不耐渍 SI | 15 | 石麦22 Shimai 22 | 锻炼不敏感且不耐渍 II |
| 8 | 郑麦004 Zhengmai 004 | 锻炼敏感且不耐渍 SI | 16 | 中麦175 Zhongmai 175 | 锻炼不敏感且不耐渍 II |
新窗口打开|下载CSV
1.2 测定指标与方法
1.2.1 土壤氧化还原电位(Eh) 使用FJA-4型氧化还原电位去极化法全自动测定仪(南京传滴仪器公司)进行土壤氧化还原电位的测定。在花前渍水锻炼处理以及花后渍水胁迫处理前、处理中和处理结束后测定, 每处理测定5个重复。1.2.2 叶绿素含量SPAD和叶绿素荧光参数 花后渍水胁迫处理最后1 d, 每品种每处理取3片生长一致且受光方向相近的旗叶测定。叶绿素含量使用叶绿素测定仪(日本, SPAD-502 Plus)测定, 叶绿素荧光参数使用便携式多功能植物测量仪MultispeQ (北京慧诺瑞德公司)进行测定, 手机端通过蓝牙连接仪器控制仪器运行并接收测量数据, 测定时用仪器测量端夹持叶片中间位置, 每处理测定3个重复。
1.2.3 干物质积累与转运 在开花期和成熟期取植株主茎, 按茎、叶、穗、籽粒等器官分样, 装入信封105℃下烘30 min, 80℃烘至恒重, 称干重, 每个处理3个重复, 每个重复2株。干物质积累与转运计算方法如下[9]:
花前贮藏物质运转量(remobilization amount of pre-anthesis stored dry matter, RAP) = 营养器官开花期干重-营养器官成熟期干重
花前贮藏物质运转量率(remobilization efficiency of pre-anthesis stored dry matter, REP) = RAP/开花期干重
花前贮藏物质对籽粒贡献率(contribution of pre-anthesis stored dry matter, CTA) = RAP/成熟期籽粒干重
花后光合同化量(post-anthesis photosynthetic assimilates accumulation, PAA) = 成熟期籽粒干重- RAP
1.2.4 可溶性糖和蔗糖含量 称取小麦叶片干样0.1 g, 加入8 mL浓度为80%的乙醇于10 mL离心管中混匀, 80℃水浴30 min, 3000×g离心10 min, 取出上清液, 重复提取3次, 定容至25 mL。提取液用于可溶性糖和蔗糖含量测定, 可溶性糖含量测定采用硫酸-蒽酮比色法[11], 蔗糖含量测定采用间苯二酚比色法[12]。
1.2.5 活性氧含量及抗氧化酶活性 称取小麦新鲜叶片0.5 g, 加入提取液5 mL pH 7.8 HEPES缓冲液(含20% (v/v)甘油, 1 mmol L-1的乙二胺四乙酸, 1 mmol L-1的抗坏血酸, 1 mmol L-1的还原型谷胱甘肽, 5 mmol L-1的氯化镁, 1 mmol L-1的二硫苏糖醇), 冰浴研磨, 10,000×g离心20 min, 取上清液。提取液用于活性氧和抗氧化酶活性测定。其中超氧阴离子(O2?)产生速率测定采用羟胺法[13], MDA含量测定采用硫代巴比妥酸法[14], 超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光还原法[15]。过氧化氢酶(CAT)活性测定采用过氧化氢法[16], 抗坏血酸氧化酶(APX)活性测定采用抗坏血酸法[17], 谷胱甘肽还原酶(GR)活性测定采用还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)还原法[18], 过氧化氢(H2O2)含量的测定采用H2O2测定试剂盒(南京建成生物工程研究所)。
1.3 统计分析
采用SPSS (Statistical Product and Service Solutions)统计分析软件对数据进行差异显著性检验, 使用Origin 2019软件作图。2 结果与分析
2.1 渍水胁迫及渍水锻炼对土壤氧化还原电位的影响
如图1所示,土壤处于正常水分状况时, 土壤氧化还原电位(Eh)在390~460 mV之间波动, 两次花前渍水锻炼处理(P)时, Eh迅速下降, 与花前未渍水锻炼处理(C)相比下降50%左右, 土壤通气状况变差, 排水恢复4 d后Eh与C无差异。在花后渍水胁迫时, Eh在胁迫前两天下降趋势与花前锻炼时基本一致, 胁迫5 d后Eh低于50 mV, 胁迫恢复6 d后, Eh与对照无差异。图1
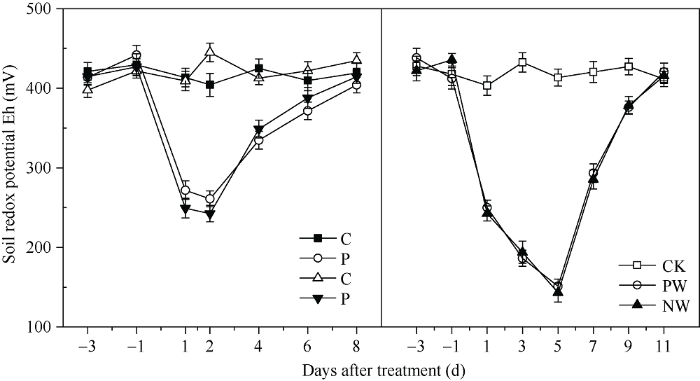 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1渍水锻炼和花后渍水胁迫下土壤氧化还原电位Eh (mV)的变化
C: 花前未渍水锻炼处理; P: 花前渍水锻炼处理; CK: 对照处理; PW: 花前渍水锻炼和花后渍水胁迫处理; NW: 花前未渍水锻炼和花后渍水胁迫处理。
Fig. 1Soil redox potential (Eh) under waterlogging priming and post-anthesis waterlogging stress in wheat
C: no pre-anthesis waterlogging priming treatment; P: pre-anthesis waterlogging priming treatment; CK: control treatment; PW: pre-anthesis waterlogging priming and post-anthesis waterlogging stress treatment; NW: no pre-anthesis waterlogging priming and post-anthesis waterlogging stress treatment.
2.2 渍水胁迫及渍水锻炼对小麦各品种产量及其构成因素的影响
花后渍水胁迫对穗数和穗粒数的影响较小, 产量降低主要是千粒重的降低所致(表3)。2019年研究结果表明, 与对照处理(CK)相比, 耐渍型品种(编号1~4和9~12)花前渍水锻炼和花后渍水处理(PW)与花前未渍水锻炼和花后渍水处理(NW)产量降幅分别为3.8%和6.0%; 不耐渍型品种(编号5~8和13~16)的PW和NW产量降幅分别为和21.9%和27.4%。锻炼敏感型品种(编号1~8)的产量锻炼指数(WPI)取值范围和均值分别为1.04~1.19和1.09, PW较NW增加9.3%; 锻炼不敏感型品种(编号9~16)的产量WPI取值范围和均值分别为0.97~1.04和1.01, PW较NW增加0.8%。耐渍型品种的PW和NW的千粒重降幅分别为7.1%和10.9%; 不耐渍型品种的PW和NW的千粒重降幅为21.5%和25.9%。锻炼敏感型品种的千粒重WPI取值范围和均值分别为1.04~1.15和1.09, PW较NW增加8.8%; 锻炼不敏感型品种的千粒重WPI取值范围和均值分别为0.97~1.06和1.02, PW较NW增加1.6%。2020年耐渍型品种和不耐渍型品种的PW和NW产量及其构成因素的趋势与2019年一致。Table 3
表3
表3渍水锻炼对花后渍水胁迫下小麦产量及其构成因素的影响
Table 3
| 品种 Variety | 处理 Treatment | 2019 | 2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 穗数 | 穗粒数 | 千粒重 | 产量 | 穗数 | 穗粒数 | 千粒重 | 产量 | ||
| Spikes | Kernels | 1000-kernel | Grain yield | Spikes | Kernels | 1000-kernel | Grain yield | ||
| pot-1 | spike-1 | weight (g) | (g plot-1) | pot-1 | spike-1 | weight (g) | (g plot-1) | ||
| 1 | CK | 22.5 a | 38.0 a | 52.1 a | 44.5 a | 24.7 a | 37.8 a | 49.9 a | 46.6 a |
| PW | 22.5 a | 40.6 a | 47.9 ab | 43.7 ab | 24.7 a | 37.9 a | 49.3 a | 46.1 a | |
| NW | 22.0 a | 41.1 a | 45.5 b | 41.1 b | 24.7 a | 37.5 a | 46.9 b | 43.3 b | |
| 2 | CK | 26.0 a | 40.3 a | 33.4 a | 35.0 a | 24.3 a | 39.5 a | 33.7 a | 32.3 a |
| PW | 26.5 a | 42.3 a | 30.1 a | 33.7 ab | 24.7 a | 40.6 a | 30.8 b | 30.8 ab | |
| NW | 26.0 a | 44.9 a | 28.1a | 32.5 b | 24.3 a | 40.1 a | 29.9 c | 29.2 b | |
| 3 | CK | 20.5 a | 42.2 a | 37.2 a | 32.2 a | 20.0 a | 42.1 b | 37.0 a | 31.1 a |
| PW | 21.0 a | 42.0 a | 35.5 a | 31.2 a | 20.0 a | 42.8 ab | 35.1 a | 30.0 a | |
| NW | 20.5 a | 43.7 a | 33.3 a | 29.8 b | 19.3 a | 43.5 a | 32.0 b | 26.9 b | |
| 4 | CK | 21.5 a | 34.8 a | 44.0 a | 32.8 a | 18.3 a | 35.5 a | 44.1 a | 28.7 a |
| PW | 20.5 a | 35.6 a | 43.4 a | 31.7 a | 19.3 a | 33.9 a | 43.6 a | 28.6 a | |
| NW | 20.5 a | 38.6 a | 37.8 b | 29.9 b | 19.0 a | 34.2 a | 40.0 b | 26.0 a | |
| 5 | CK | 22.0 a | 36.6 a | 51.8 a | 41.7 a | 23.0 a | 38.3 a | 51.4 a | 45.3 a |
| PW | 22.5 a | 36.3 a | 46.2 ab | 37.7 ab | 23.3 a | 38.7 a | 40.7 b | 36.7 b | |
| NW | 22.0 a | 37.3 a | 40.4 b | 33.1 b | 21.7 a | 39.9 a | 38.3 c | 33.1 b | |
| 6 | CK | 19.5 a | 27.8 a | 61.6 a | 33.4 a | 26.3 a | 28.6 a | 59.2 a | 44.7 a |
| PW | 20.5 a | 27.4 a | 47.6 b | 26.7 b | 26.7 a | 28.1 a | 55.3 b | 41.3 b | |
| NW | 20.5 a | 26.6 a | 45.7 b | 24.9 b | 25.7 a | 29.3 a | 47.7 c | 35.9 c | |
| 7 | CK | 24.0 a | 42.7 a | 45.6 a | 46.7 a | 29.3 a | 42.3 a | 44.1 a | 54.7 a |
| PW | 24.0 a | 43.5 a | 36.9 b | 38.5 b | 29.3 a | 43.8 a | 37.4 b | 48.1 b | |
| NW | 23.5 a | 41.9 a | 34.4 b | 33.9 b | 30.0 a | 42.7 a | 33.0 c | 42.3 c | |
| 8 | CK | 17.0 a | 40.1 a | 50.3 a | 34.3 a | 21.3 a | 38.2 a | 47.4 a | 38.6 a |
| PW | 17.0 a | 40.4 a | 38.6 b | 26.5 b | 22.0 a | 37.3 a | 39.7 b | 32.6 b | |
| NW | 16.5 a | 38.9 a | 34.8 b | 22.3 c | 21.3 a | 37.5 a | 37.5 c | 29.9 b | |
| 9 | CK | 21.5 a | 31.7 a | 49.0 a | 33.3 a | 32.7 a | 31.7 a | 44.7 a | 46.1 a |
| PW | 22.0 a | 33.9 a | 43.4 a | 32.3 a | 33.0 a | 31.6 a | 42.3 b | 44.1 a | |
| NW | 22.0 a | 32.2 a | 44.7 a | 31.7 a | 32.3 a | 31.6 a | 42.2 b | 43.1 a | |
| 10 | CK | 22.0 a | 37.6 a | 50.0 a | 41.1 a | 22.7 a | 38.7 a | 47.9 a | 42.0 a |
| PW | 21.5 a | 39.8 a | 45.2 b | 38.5 a | 23.0 a | 37.8 a | 47.1 a | 40.9 a | |
| NW | 21.5 a | 39.7 a | 45.9 b | 39.1 a | 23.0 a | 37.3 a | 46.5 a | 39.8 a | |
| 11 | CK | 23.0 a | 29.3 a | 43.1 a | 29.0 a | 24.7 a | 29.5 a | 41.4 a | 30.1 a |
| PW | 22.5 a | 31.4 a | 39.6 b | 27.9 a | 23.7 a | 30.1 a | 40.6 a | 28.9 a | |
| NW | 23.0 a | 32.3 a | 37.3 b | 27.7 a | 23.3 a | 31.2 a | 37.3 b | 27.2 a | |
| 12 | CK | 20.5 a | 48.2 a | 31.3 a | 32.4 a | 12.7 a | 49.6 a | 32.6 a | 20.4 a |
| PW | 19.5 a | 51.9 a | 30.2 a | 30.6 a | 12.0 a | 50.7 a | 33.2 a | 20.2 a | |
| NW | 19.5 a | 50.3 a | 30.1 a | 31.5 a | 12.0 a | 51.3 a | 31.2 b | 19.2 a | |
| 13 | CK | 23.0 a | 40.2 a | 53.2 a | 49.2 a | 26.3 a | 40.1 a | 48.8 a | 51.6 a |
| PW | 22.5 a | 41.0 a | 37.2 b | 34.2 b | 26.0 a | 39.6 a | 40.4 b | 41.5 b | |
| NW | 23.0 a | 40.1 a | 36.5 b | 33.6 b | 24.7 a | 41.0 a | 41.7 b | 42.2 b | |
| 14 | CK | 23.5 a | 27.3 a | 62.9 a | 40.4 a | 28.7 a | 26.2 a | 54.3 a | 40.7 a |
| PW | 23.5 a | 26.5 a | 43.2 b | 26.9 b | 28.3 a | 25.9 a | 45.3 b | 33.2 b | |
| 品种 Variety | 处理 Treatment | 2019 | 2020 | ||||||
| 穗数 | 穗粒数 | 千粒重 | 产量 | 穗数 | 穗粒数 | 千粒重 | 产量 | ||
| Spikes | Kernels | 1000-kernel | Grain yield | Spikes | Kernels | 1000-kernel | Grain yield | ||
| pot-1 | spike-1 | weight (g) | (g plot-1) | pot-1 | spike-1 | weight (g) | (g plot-1) | ||
| NW | 23.5 a | 26.6 a | 41.6 b | 26.0 b | 27.3 a | 27.1 a | 45.7 b | 33.9 b | |
| 15 | CK | 27.5 a | 24.9 a | 53.6 a | 36.8 a | 35.0 a | 24.1 a | 51.4 a | 43.3 a |
| PW | 26.5 a | 24.3 a | 43.8 b | 28.2 b | 35.0 a | 24.6 a | 36.8 b | 31.7 b | |
| NW | 25.0 a | 25.3 a | 43.0 b | 27.1 b | 36.0 a | 23.7 a | 35.1 c | 30.0 b | |
| 16 | CK | 19.0 a | 35.6 a | 51.8 a | 35.0 a | 16.7 a | 36.2 a | 50.1 a | 30.1 a |
| PW | 19.0 a | 35.0 a | 43.4 b | 28.8 b | 16.3 a | 35.7 a | 40.1 b | 23.4 b | |
| NW | 20.0 a | 34.6 a | 42.1 b | 29.1 b | 16.3 a | 36.7 a | 37.6 c | 22.6 b | |
新窗口打开|下载CSV
2.3 渍水胁迫及渍水锻炼对小麦各品种旗叶SPAD和叶绿素荧光参数的影响
渍水胁迫下旗叶叶绿素相对含量SPAD降低。耐渍型品种(编号1~4和9~12)的PW和NW的SPAD降幅分别为4.6%和7.7%; 不耐渍型品种(编号5~8和13~16)的PW和NW的SPAD降幅分别为19.3%和23.9%。锻炼敏感型品种的SPAD锻炼指数(WPI)取值范围和均值分别为1.02~1.15和1.09, PW较NW增加8.8%; 锻炼不敏感型品种SPAD的WPI取值范围和均值分别为0.97~1.04和1.01, PW较NW增加1.0% (图2)。图2
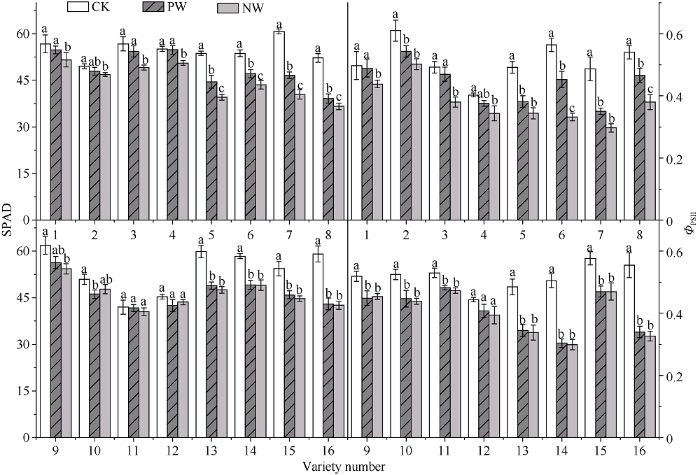 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2渍水锻炼对花后渍水胁迫下小麦叶片叶绿素含量(SPAD)和实际光化学效率(ΦPSII)的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 2Effects of waterlogging priming on SPAD and ΦPSII under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
花后渍水胁迫降低了旗叶实际光化学效率ΦPSII, 耐渍型品种的锻炼处理和未锻炼处理ΦPSII的降幅均值分别为8.7%和14.8%; 不耐渍型品种的锻炼处理和未锻炼处理的降幅为26.3%和33.8%。锻炼敏感型品种ΦPSII的WPI取值范围和均值分别为1.08~1.36和1.18, PW较NW增加17.6%; 锻炼不敏感型品种ΦPSII的WPI取值范围和均值分别为0.99~1.04和1.02, PW较NW增加1.7% (图2)。
花后渍水胁迫处理使旗叶非调节性能量耗散ΦNO和调节性能量耗散ΦNPQ升高, 耐渍型品种锻炼处理和未锻炼处理的ΦNO的增幅分别为3.7%和10.2%, ΦNPQ分别为21.2%和32.9%; 不耐渍型品种的PW和NW处理的增幅为27.6%和33.5%, ΦNPQ的增幅分别为34.5%和47.2%。由于各品种的ΦNPQ和ΦNO的数值整体上表现为NW高于PW, 故计算ΦNPQ和ΦNO的锻炼系数时需取计算值的倒数, O2?产生速率、H2O2含量和MDA含量的WPI的计算同上。如图3所示, 锻炼敏感型品种ΦNO的WPI取值范围和均值分别为1.02~1.20和1.11, NW较PW增加10.7%; ΦNPQ的WPI取值范围和均值分别为1.02~1.56和1.17, NW较PW增加16.5%。锻炼不敏感型品种的ΦNO的WPI取值范围和均值分别为0.96~1.09和1.01, NW较PW增加0.9%; ΦNPQ的WPI取值范围和均值分别为0.93~1.07和1.02, NW较PW增加2.2%。
图3
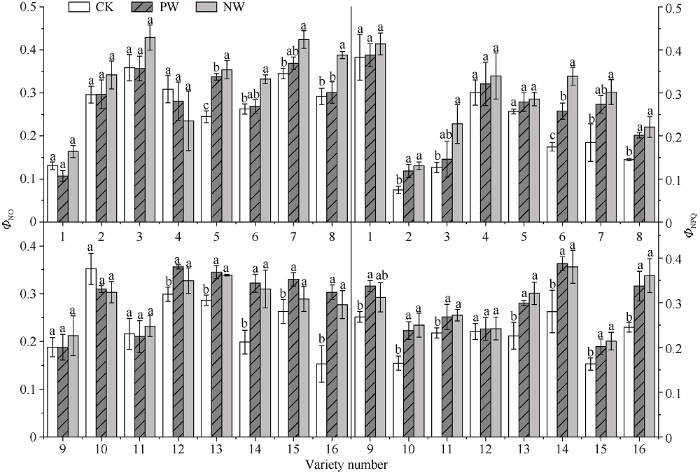 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3渍水锻炼对花后渍水胁迫下小麦叶片非调节性能量耗散(ΦNO)和调节性能量耗散(ΦNPQ)的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 3Effects of waterlogging priming on ΦNO and ΦNPQ under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
2.4 渍水胁迫及渍水锻炼对小麦各品种干物质积累与转运的影响
与对照相比, 渍水胁迫提高了小麦营养器官花前贮藏物质转运量(RAP)、花前贮藏物质转运率(REP)和花前贮藏物质对籽粒的贡献率(CTA), 降低了花后光合同化量(PAA)。渍水胁迫下耐渍型品种(编号1~4和9~12)的RAP、REP、CTA的增幅分别为7.2%、9.9%、17.2%; 不耐渍型品种(编号5~8和13~16)的增幅分别为37.5%、42.2%、75.9%。渍水胁迫下耐渍型品种的PAA的降幅分别为14.0%, 不耐渍型品种的降幅为38.7%。与锻炼不敏感型品种相比, 锻炼敏感型品种的干物质积累与转运能力较未锻炼处理的提升较大。锻炼敏感型品种RAP的WPI取值范围和均值分别为1.09~1.40和1.22, REP分别为1.12~1.31和1.20, CTA分别为0.98~1.12和1.05, PAA分别为1.07~1.19和1.11, 4项指标的PW较NW分别提高21.6%、20.0%、5.5%和10.8%; 锻炼不敏感型品种RAP的WPI取值范围和均值分别为0.96~1.06和1.01, REP分别为0.92~1.11和1.00, CTA分别为0.95~1.04和1.00, PAA分别为0.93~1.08和1.01, 4项指标的PW较NW分别提高0.6%、-0.4%、-0.4%和0.8% (图4)。图4
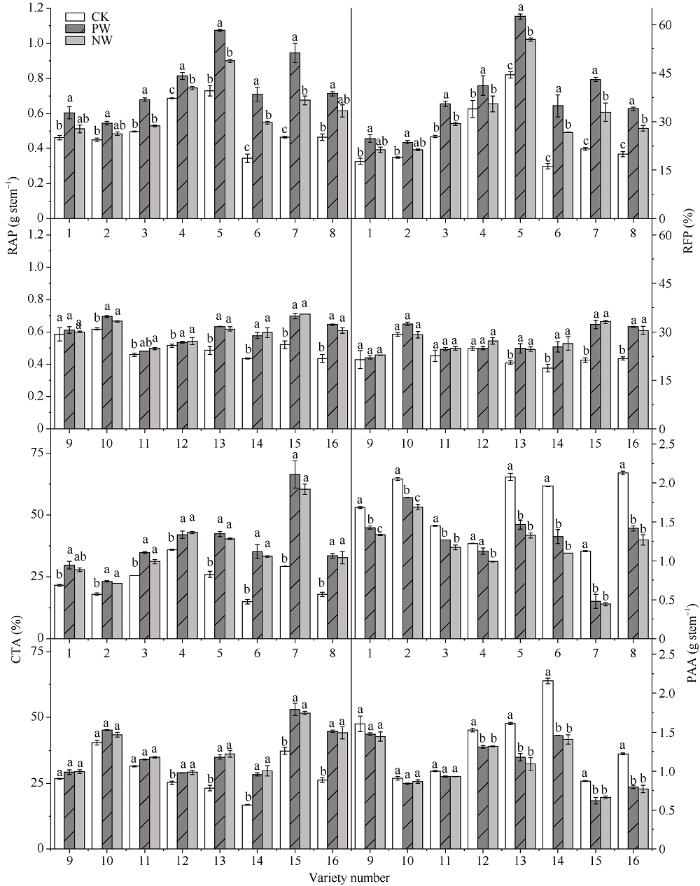 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4渍水锻炼对花后渍水胁迫下小麦干物质积累与转运的影响
RAP: 花前贮藏物质转运量; REP: 花前贮藏物质转运率; CTA: 花前贮藏物质对籽粒贡献率; PAA: 花后光合同化量。不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 4Effects of waterlogging priming on dry matter accumulation and translocation under post-anthesis waterlogging stress in wheat
RAP: remobilization amount of pre-anthesis stored dry matter; REP: remobilization efficiency of pre-anthesis stored dry matter; CTA: contribution of pre-anthesis stored dry matter; PAA: post-anthesis photosynthetic assimilates accumulation. Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
2.5 渍水胁迫及渍水锻炼对小麦各品种旗叶可溶性糖和蔗糖含量的影响
与对照(CK)相比, 花后渍水胁迫降低了可溶性糖含量和蔗糖含量; 耐渍型品种的PW和NW的可溶性糖含量的降幅分别为13.9%和20.3%; 不耐渍型品种的PW和NW的降幅为40.0%和45.1%。锻炼敏感型品种可溶性糖含量的WPI的取值范围和均值分别为1.08~1.33和1.17, PW较NW增加21.9%; 对应锻炼不敏感型品种分别为0.97~1.05和1.01, PW较NW增加1.7%。耐渍型品种的PW和NW蔗糖含量降幅分别为3.2%和19.9%, 不耐渍型品种的PW和NW的降幅为20.0%和29.8%。锻炼敏感型品种蔗糖含量的WPI的取值范围和均值分别为1.06~ 1.35和1.22, PW较NW增加21.8%; 对应锻炼不敏感型品种分别为0.98~1.08和1.04, PW较NW增加3.1% (图5)。图5
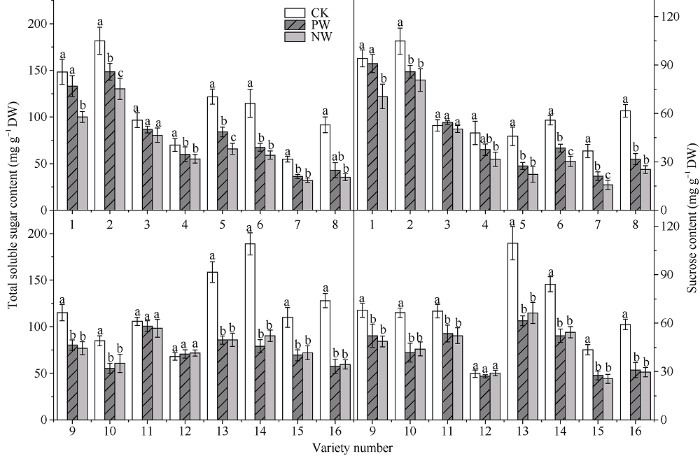 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5渍水锻炼对花后渍水胁迫下小麦叶片可溶性总糖含量和蔗糖含量的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 5Effects of waterlogging priming on the contents of soluble total sugar and sucrose under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
2.6 渍水胁迫及渍水锻炼对小麦各品种旗叶活性氧积累的影响
2.6.1 O2?产生速率和H2O2含量 渍水胁迫下各品种旗叶的O2?产生速率上升, 各品种在对照处理与胁迫处理间达显著差异。如图6所示, 耐渍型品种旗叶的O2?产生速率在PW和NW下分别增加了48.7%和68.5%, 不耐渍型品种增加了128.3%和167.9%。锻炼敏感型品种O2?产生速率的WPI的取值范围和均值分别为1.07~1.40和1.25, NW较PW提高27.7%; 对应锻炼不敏感型品种分别为0.92~1.33和1.08, NW较PW提高6.0%。图6
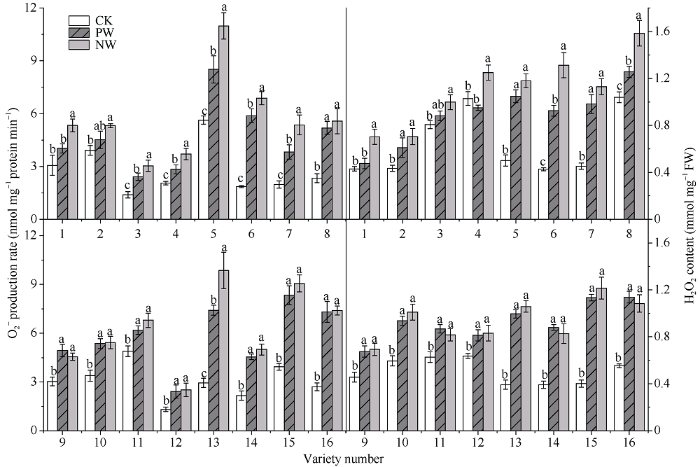 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6渍水锻炼对花后渍水胁迫下小麦叶片超氧阴离子自由基(O2?)产生速率和过氧化氢(H2O2)含量的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 6Effects of waterlogging priming on O2? production rate and H2O2 content under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
渍水胁迫下各品种旗叶的H2O2含量出现不同程度的增加, 耐渍型品种的H2O2含量整体上较不耐渍型品种更低, 耐渍型品种和不耐渍型品种旗叶的H2O2含量在PW和NW下分别整体增加了28.0%和44.1%; 不耐渍型品种旗叶的H2O2含量在PW和NW增加了116.3%和140.6%。锻炼敏感型品种H2O2含量的WPI的取值范围和均值分别为1.13~ 1.48和1.25, NW较PW提高25.8%; 对应锻炼不敏感型品种分别为0.94~1.08和1.01, NW较PW提高1.2% (图6)。
2.6.2 MDA含量 渍水胁迫下各品种旗叶的MDA含量均出现不同程度的上升; 与CK相比, 耐渍型品种的PW和NW的MDA含量的增幅均值分别为34.7%和38.3%; 不耐渍型品种的PW和NW增幅为93.5%和110.7%。锻炼敏感型品种MDA含量的WPI的取值范围和均值分别为1.07~1.24和1.14, NW较PW提高15.9%; 对应锻炼不敏感型品种分别为0.85~1.07和0.98, NW较PW提高-0.7% (图7)。
图7
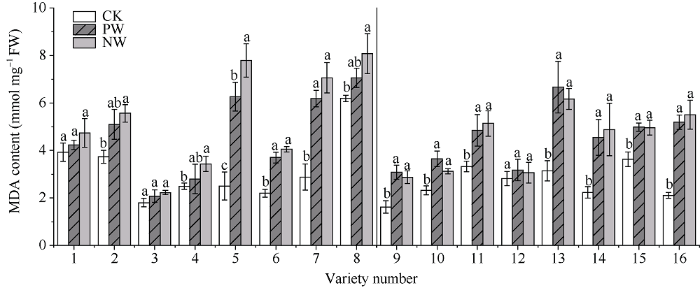 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7渍水锻炼对花后渍水胁迫下小麦叶片丙二醛(MDA)含量的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 7Effects of waterlogging priming on MDA content under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
2.7 渍水胁迫及渍水锻炼对小麦各品种旗叶抗氧化酶活性的影响
渍水胁迫下, 耐渍型小麦品种的超氧化物歧化酶(SOD)活性均出现不同程度的上升, 在PW和NW下增幅均值分别为29.3%和16.7%; 不耐渍型品种的SOD活性在PW和NW下整体下降, 降幅均值分别为0.9%和7.5%。渍水胁迫下, 锻炼敏感型品种的SOD活性的WPI的取值范围和均值分别为1.10~1.24和1.16, PW较NW提高16.9%; 对应锻炼不敏感型品种分别为0.97~1.10和1.03, PW较NW提高1.6%。过氧化氢酶(CAT)可催化H2O2分解为H2O和O2?, 减轻过氧化氢过度积累对植物细胞的损害。由图8可知, 渍水胁迫下, 耐渍型小麦品种的过氧化氢酶活性均出现不同程度的上升, 增幅均值为37.3%; 而不耐渍型品种的CAT活性较CK整体降低了4.1%。锻炼敏感型品种的CAT活性的WPI的取值范围和均值分别为1.09~1.32和1.19, PW较NW提高19.1%; 对应锻炼不敏感型品种分别为0.93~1.09和1.02, PW较NW提高1.5% (图8)。图8
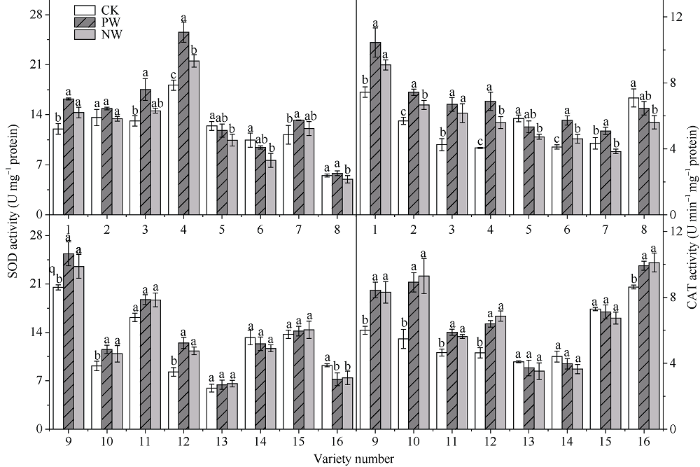 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8渍水锻炼对花后渍水胁迫下小麦叶片超氧化物歧化酶(SOD)活性和过氧化氢酶(CAT)活性的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 8Effects of waterlogging priming on the activities of SOD and CAT under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
除品种镇麦10 (编号5), 耐渍型小麦品种的谷胱甘肽还原酶(GR)活性均出现不同程度的上升,在PW和NW下增幅均值分别为36.1%和15.5%; 而不耐渍型品种的GR活性在渍水胁迫下均降低, 各品种的PW和NW的GR活性分别下降了28.1%和37.4%。锻炼敏感型品种的GR活性的WPI的取值范围和均值分别为1.05~1.57和1.31, PW较NW提高28.4%; 对应锻炼不敏感型品种分别为0.96~1.08和1.01, PW较NW提高0.4% (图9)。抗坏血酸过氧化物酶(APX)是植物叶绿体清除过氧化氢的关键酶。如图9所示, 耐渍型小麦品种的APX活性均出现不同程度的上升, 在PW和NW下增幅均值分别为32.6%和27.2%; 而不耐渍型品种的APX在渍水胁迫活性整体降低, PW和NW的下APX活性的降幅均值分别为3.3%和11.6%。锻炼敏感型品种的APX活性的WPI的取值范围和均值分别为1.03~1.23和1.09, PW较NW提高9.1%; 对应锻炼不敏感型品种分别为0.92~1.15和1.05, PW较NW提高5.0%。
图9
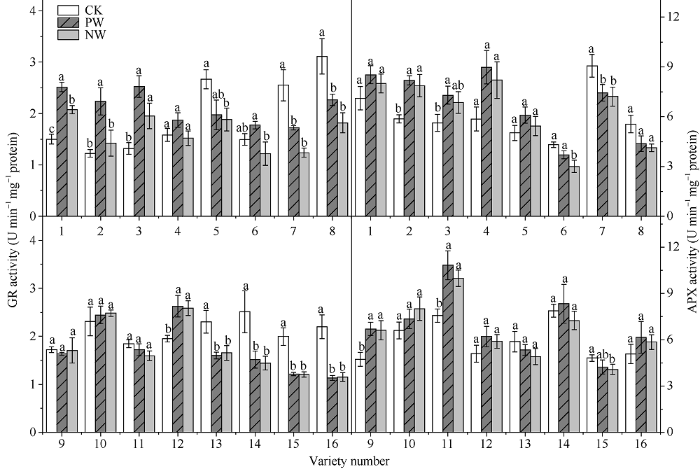 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9渍水锻炼对花后渍水胁迫下小麦叶片谷胱甘肽还原酶(GR)活性和抗坏血酸过氧化物酶(APX)活性的影响
不同字母表示品种内处理间在P < 0.05水平差异显著。品种信息同
Fig. 9Effects of waterlogging priming on the activities of GR and APX under post-anthesis waterlogging stress in wheat
Different lowercase letters indicate significant differences at P < 0.05 among different treatments of each variety. Varieties are as the same as in
3 讨论
渍水锻炼处理(短期渍水)下土壤氧化还原电位(Eh)降幅较低, 对小麦生育早期植株发育无明显影响; 而渍水胁迫处理(长期渍水)下Eh急剧下降, 对植株发育造成不可逆的伤害[10]。渍水胁迫条件下叶绿素的合成受到抑制, 过多的水分致使叶绿体膨胀使膜被破坏或消失, 从而引起叶绿素含量的降低, 不仅引发光合结构的异常[19], 同时也影响光合电子的传递[20]。本研究下, 小麦叶片叶绿素含量和实际光化学效率ΦPSII的降低表明光合能力受到损伤。与不耐渍型品种相比, 耐渍型品种旗叶实际光化学效率ΦPSII和SPAD的降幅较低, 且非调节性能量耗散ΦNO和可调性能量耗散ΦNPQ的升高比例较低, 与前人研究结果一致[21], 这说明耐渍性品种通过延缓叶片叶绿素含量降解, 维持较低的光能热耗散和较高的光能利用率,最终维持叶片光合作用。此外, 与锻炼不敏感型品种相比, 锻炼敏感型品种经过锻炼的植株(PW) ΦPSII和SPAD较未锻炼植株(NW)的增幅较高, 而ΦNO和ΦNPQ较NW的增幅较低, 说明经过渍水锻炼植株可以在渍水胁迫下维持较高的叶绿素含量及光合结构的稳定性, 从而维持较高的光合能力, 这与前人结果一致[9]。渍水胁迫会显著降低小麦产量, 作物产量一方面来自光合同化积累, 另一方面来自于营养器官同化物的转运, 前人研究发现, 在小麦各个生育时期进行渍水胁迫均会抑制花前贮藏物质向籽粒的转运, 缩短灌浆时间, 降低花后同化物的积累, 从而导致千粒重的降低[22,23]。与不耐渍型品种相比, 耐渍型品种可通过提高其花后光合同化量来减轻渍害对植株籽粒干物质积累的影响, 降低千粒重降幅, 降低减产幅度。同时, 与NW相比, PW下锻炼敏感型品种的花后光合同化量、花前贮藏物质转运量、花前贮藏物质转运率以及花前贮藏物质对籽粒贡献率整体增幅均高于锻炼不敏感型品种, 这说明在花后渍水胁迫下, 锻炼敏感型品种可通过维持较高的干物质向籽粒转运的比例以及较高的花后光合同化量, 减少渍水胁迫下产量的损失, 这与在高温锻炼上的研究结果类似[24]。渍水胁迫导致的光合速率的降低可使电子传递给O2形成活性氧O2?和H2O2等物质, 加剧膜脂过氧化伤害, 对植物细胞膜、DNA和蛋白质造成严重损伤[25,26]。而植株可通过提高体内SOD、POD、CAT等抗氧化酶活性来清除体内多余的活性氧[27,28]。研究发现, 花后渍水胁迫下小麦旗叶的抗氧化酶活性随处理时间的延长而逐渐降低, 耐渍型品种旗叶抗氧化酶的降低幅度低于不耐渍型品种[14]。本试验研究显示, PW和NW下各品种旗叶的O2?产生速率和H2O2含量均上升, 导致膜脂过氧化程度加剧, 使膜脂过氧化产物丙二醛(MDA)含量增加; 活性氧和MDA含量的增幅取决于品种耐渍性强弱, 多数耐渍型品种旗叶活性氧和MDA含量的增幅低于不耐渍型品种。渍水胁迫下不同类型品种旗叶抗氧化酶活性变化不一, 耐渍型品种PW和NW植株旗叶的CAT、SOD、GR和APX活性均高于CK; 而不耐渍型品种植株PW和NW的旗叶抗氧化酶活性较CK增幅较小, 部分品种的抗氧化酶活性与CK相比增幅不显著甚至略有下降, 这与前人结果一致[15]。渍水胁迫下, 不耐渍型品种衡4399、京冬22、鲁垦麦9号、郑麦004、济麦22、济南矮6号、石麦22以及中麦175的抗氧化酶活性无法升高可能是因其耐渍性太弱, 破坏了叶片中酶结构从而导致酶活性无法有效提升。小麦经渍水锻炼处理后, 生育后期再次遭受相同或不同胁迫时, 旗叶抗氧化酶活性增强, 可在一定程度上抑制叶片活性氧类物质的大量积累, 使膜脂过氧化程度降低, 降低对细胞膜的损伤, 使旗叶衰老速率减慢[9,10]。锻炼处理下各品种的旗叶抗氧化酶活性的整体增幅高于未锻炼处理, 表明花前渍水锻炼可提高花后渍水胁迫下旗叶的抗氧化酶活性, 进而增强对活性氧的清除能力, 减轻膜脂过氧化程度, 使旗叶能够维持较高的光合同化能力, 最终使产量损失降低。不同品种对渍水锻炼响应的敏感程度不同, 锻炼处理后锻炼敏感型品种旗叶抗氧化酶活性的增幅高于锻炼不敏感型品种。可溶性糖在植物抵御逆境胁迫的过程中发挥着重要的作用, 同时可作为渗透调节物质和其他有机物质合成的碳架和能量来源[29]。渍水胁迫下可溶性糖的积累既可提高细胞溶质浓度, 降低细胞水势, 增强植物细胞吸收功能, 又可稳定亚细胞结构和大分子结构[30]。有研究表明, 在渍水胁迫条件下, 与耐渍性弱的品种相比, 耐渍性强的植株体内可溶性糖和脯氨酸含量增幅较大[31], 渍水胁迫下, 各品种PW和NW旗叶的可溶性糖和蔗糖含量均下降; 耐渍型品种旗叶糖含量的整体降幅小于不耐渍型品种。前人研究结果表明, 经渍水锻炼后小麦叶片中可溶性糖和游离氨基酸含量均比未锻炼处理显著增加, 使叶片的渗透调节能力提高[9]。本文结果表明, 经锻炼处理的植株可溶性糖含量高于未锻炼植株, 表明渍水锻炼可提升渍水胁迫下小麦旗叶中可溶性糖和蔗糖含量。渍水锻炼敏感型品种旗叶的可溶性糖和蔗糖含量的增幅较高, 而锻炼不敏感型品种对渍水锻炼响应迟钝, 其旗叶糖含量提高幅度较小, 与未锻炼处理无显著差异, 说明锻炼敏感型小麦品种可以通过提高体内可溶性糖和蔗糖含量, 维持植株体内的能量供应, 增强小麦的抗渍性。
4 结论
花后渍水胁迫降低了小麦籽粒产量主要是降低了叶绿素含量和实际光化学效率, 增强了热耗散, 同时降低了花前积累的光合同化产物向籽粒中的转运, 同时花后光合同化量降低, 粒重减少, 最终导致籽粒产量下降; 花后渍水胁迫降低了叶片可溶性糖含量和蔗糖含量, 增加了H2O2含量, O2?的产生速率以及MDA含量, 表明膜脂过氧化伤害加剧。渍水胁迫下, 不耐渍型品种活性氧的积累及膜脂过氧化伤害加剧, 耐渍型品种的叶片光合能力和抗氧化能力受影响较小, 产量损失率较低。花前渍水锻炼提高了花后渍水条件下小麦旗叶光合能力, 促进了花前贮藏物质的转运和花后光合物的积累, 最终提高了籽粒产量; 花前渍水锻炼降低了活性氧类物质的含量, 提高了抗氧化酶的活性, 增强了小麦旗叶的抗氧化能力, 增强了小麦对花后渍水胁迫的抵御能力。与渍水锻炼不敏感型品种相比, 渍水锻炼敏感型品种其旗叶光合能力和抗氧化能力较未锻炼的植株提升幅度更高, 最终使籽粒产量的降幅较低。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 5]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
In: Kranner I C, Beckett R P, Varma A K, eds. Protocols in Lichenology. Springer Lab Manuals.
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}