 ,1, 蒋成功1,2, 王红武1, 马庆2, 李坤1, 刘志芳1, 吴宇锦1, 李树强1, 胡小娇,1,*, 黄长玲,1,*
,1, 蒋成功1,2, 王红武1, 马庆2, 李坤1, 刘志芳1, 吴宇锦1, 李树强1, 胡小娇,1,*, 黄长玲,1,*Phenotype identification and gene mapping of defective kernel 48 mutant (dek48) in maize
SHI Hui-Min,1, JIANG Cheng-Gong1,2, WANG Hong-Wu1, MA Qing2, LI Kun1, LIU Zhi-Fang1, WU Yu-Jin1, LI Shu-Qiang1, HU Xiao-Jiao,1,*, HUANG Chang-Ling,1,*通讯作者:
收稿日期:2020-01-20接受日期:2020-04-15网络出版日期:2020-09-12
| 基金资助: |
Received:2020-01-20Accepted:2020-04-15Online:2020-09-12
| Fund supported: |
作者简介 About authors
E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (8650KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
石慧敏, 蒋成功, 王红武, 马庆, 李坤, 刘志芳, 吴宇锦, 李树强, 胡小娇, 黄长玲. 玉米籽粒突变体dek48的表型鉴定与基因定位[J]. 作物学报, 2020, 46(9): 1359-1367. doi:10.3724/SP.J.1006.2020.03005
SHI Hui-Min, JIANG Cheng-Gong, WANG Hong-Wu, MA Qing, LI Kun, LIU Zhi-Fang, WU Yu-Jin, LI Shu-Qiang, HU Xiao-Jiao, HUANG Chang-Ling.
玉米(Zea mays L.)是重要的粮食、饲料和工业加工原料, 也是我国第一大作物[1]。籽粒是玉米主要的营养储存器官, 籽粒的正常发育决定了其营养品质和经济价值。挖掘控制籽粒发育的关键基因并解析其参与的代谢网络, 对于提高玉米产量和营养价值具有重要的理论意义。
籽粒突变体是研究禾本科植物种子发育的重要材料。不同类型的玉米籽粒突变体, 如胚缺陷、胚乳缺陷、空果皮等被广泛应用于籽粒发育相关基因的克隆和功能解析研究[2]。籽粒发育缺陷突变体(defective kernel, dek)是一类胚和胚乳发育缺陷比较严重的突变体, 大多数由于胚致死导致成熟籽粒不能自然萌发[3]。dek突变体较野生型籽粒体积更小, 颜色更浅, 营养物质积累缓慢, 胚和胚乳滞缓发育, 一般在授粉后11~17 d可观察到明显的表型; 成熟的dek籽粒表面皱缩, 种皮颜色灰白, 硬度较小, 胚乳大部分呈粉质[4,5]。
国内外有许多关于玉米dek突变体的报道, 目前已有多个dek突变体基因被克隆。大部分已研究过的dek突变体均与三角状五肽重复蛋白(pentatricopeptide repeat protein, PPR)有关。PPR蛋白大多定位在线粒体或叶绿体, 靶向位点专一, 是一类参与线粒体和质体中RNA编辑和蛋白质翻译的RNA识别蛋白[6,7]。目前植物中已发现的PPR蛋白种类超过450种, 广泛地参与了植物生长发育的各个过程。在籽粒发育过程中, PPR蛋白发生突变往往会造成线粒体功能受损, 进而导致能量代谢受阻, 严重影响胚和胚乳的发育[8], 如dek2、dek10、dek35、dek36、dek37、dek39、dek40、dek41等[9,10,11,12,13,14,15,16]。Dek2编码P亚家族的PPR蛋白, 该蛋白功能的丧失导致线粒体复合体I的亚基nad1的第1个内含子剪接效率降低[9]; Dek10编码E亚家族的PPR蛋白, 基因突变后导致线粒体复合体I的亚基nad3和复合体IV的亚基cox2功能丧失[10]; Dek35编码P亚家族的PPR蛋白, 基因突变后线粒体复合体I的亚基nad4第1个内含子剪接效率降低[11]; Dek36编码E+亚家族的PPR蛋白, 基因突变后导致线粒体复合体I的atp4-59、复合体III的nad7-383、复合体IV的ccmFN-302共3个位点的编辑效率降低[12]; Dek37编码P亚家族的PPR蛋白, 该蛋白功能的丧失导致线粒体复合体I的亚基nad2的第1个内含子剪接效率降低[13]; Dek39编码一个参与多位点RNA编辑的E亚家族PPR蛋白, 基因突变后影响线粒体复合体I的nad3-247、nad3-275两个位点的编辑效率[14]; Dek40编码一个位于线粒体中的E+亚家族PPR蛋白, 基因突变后导致线粒体复合体I的亚基nad2-26和nad5-1916、复合体IV的亚基cox3-314这3个位点的编辑功能丧失[15]; Dek41编码P亚家族的PPR蛋白, 该蛋白功能的丧失导致线粒体复合体I的亚基nad4的第3个内含子剪接效率降低[16]。以上这些与RNA剪接和编辑相关的PPR蛋白, 其功能的丧失均会导致籽粒异常发育。也有一些dek突变基因的蛋白功能与PPR蛋白无关, 例如, Dek15编码一个姐妹染色单体粘连蛋白4 (SCC4), 突变后破坏有丝分裂细胞周期和内复制[17]; Dek33编码核黄素合成中的关键酶, 影响种子发育中的油体和ABA[18]; dek38基因编码TTI2 (tel2-interaction protein 2)分子伴侣蛋白, 突变后胚致死[19]; Dek42通过与其他剪接体的互作参与了前信使RNA剪接的调控[20]; Dek44功能的丧失影响了线粒体和核基因组中呼吸链相关蛋白编码基因的表达, 导致籽粒异常发育[21]。
本研究以EMS诱变的玉米籽粒突变体defective kernel 48 (dek48)为研究材料。通过开展表型鉴定、籽粒成分测定、遗传分析和基因定位, 为后续该基因的克隆和功能解析奠定了良好基础。
1 材料与方法
1.1 实验材料
玉米自交系郑58花粉经甲基磺酸乙酯(EMS)诱变处理后自交得到诱变一代(M1)突变体库。通过连续自交和筛选鉴定, 获得性状稳定遗传的籽粒缺陷突变体dek48。由于dek48突变体不能成苗, 因而将自交的M4和M5代杂合植株(+/dek48)与玉米自交系B73、Mo17、昌7-2 (C7-2)分别杂交得到F1群体, F1自交获得的F2群体用于后续的遗传分析和基因定位研究。1.2 突变体的表型鉴定和遗传分析
观察授粉后不同天数正常籽粒和突变籽粒的表型, 分析它们在籽粒大小、结构、胚和胚乳发育进程等方面的差异。选取M5代自交授粉后16 d的dek48突变体和野生型籽粒制作石蜡切片进行结构观察。通过卷纸发芽试验观察突变籽粒的发芽率。分别计算B73、C7-2、Mo17背景的F2分离群体中正常籽粒和突变籽粒的分离比, 利用卡方测验进行分离比检验。1.3 玉米籽粒主要成分测定
成熟籽粒粗淀粉使用Megazyme淀粉总量检测试剂盒(Cat#K-TSTA)提取, 用分光光度仪在510 nm下测定籽粒淀粉样品的吸光度值。利用布鲁克MPA近红外光谱仪(布鲁克科技有限公司, 德国)对成熟籽粒进行油分和蛋白测定。1.4 胚乳的扫描电镜观察
选取突变体和正常郑58的成熟籽粒, 置于65℃烘箱内干燥48 h, 在籽粒中部轻敲, 使其自然裂开。将样品固定于检测铜台上, 置于离子溅射仪中镀金膜, 利用日立SU8020扫描电镜(日立高新技术公司, 日本)进行胚乳结构观察。1.5 基因定位及候选基因分析
在F2代定位群体中随机选取表型分离比正确的果穗, 利用CTAB法提取48粒突变籽粒及其双亲的基因组DNA[22]。利用20 K GBTS (genotyping by target sequencing)靶向测序技术[23]分析样本基因型。计算每个多态性SNP标记的基因型频率(SNP-index), 即亲本型基因型占所有F2样本基因型的频率, SNP-index越接近1, 表明标记与目标基因越连锁[24]。将F2定位群体的突变籽粒数量增加至2300个, 并开发6对InDel标记In6.38、In7.29、In7.39、In8.29、In8.53、In9.06 (表1), 缩小定位区间。开发SNP标记, 利用KASP (kompetitive allele-specific PCR)基因分型技术分析重组单株基因型, 完成基因精细定位。利用Gramene (Table 1
表1
表1基因定位引物序列
Table 1
| 引物 Primer | 正向序列 Forward sequence (5′-3′) | 反向序列 Reverse sequence (5′-3′) |
|---|---|---|
| In6.38 | TTGGCATCTTGTGGTTTTGTG | CGAACGGAGTGAAAGTGATAGAAG |
| In7.29 | GCTCAAGAAAGCAATCGGGTAA | GTGATGAAGCCAGGATTCGCT |
| In7.39 | TTTCTGGCTCTTTATTTGTGCTG | CAAGCGTACATACACCTGTCAGA |
| In8.29 | ATGCAGTTAAGTTGACCTGAATTG | TATAGCTAGGAAACCAAGGCGT |
| In8.53 | GCTTTTTCCACCACTGCGAC | ATACTCGGGGGCTGCCTTAC |
| In9.06 | GGAAAACACTAAAACACATAACCCT | CTAAGGCGTCACTTTGCGG |
新窗口打开|下载CSV
选取正常籽粒与突变籽粒表型3︰1分离的F2果穗, 采用CTAB[22]和碱煮法[25]提取亲本和突变体的基因组DNA。扩增体系共20 μL: 2×PCR Mix 10 μL、正向和反向引物(10 μmol L-1)各1 μL、基因组DNA 2 μL、ddH2O 6 μL。PCR程序为: 95℃预变性3 min; 94℃变性30 s, 60℃退火30 s, 72℃延伸30 s, 共35个循环; 72℃终延伸5 min。利用3%琼脂糖凝胶电泳检测PCR产物。
2 结果与分析
2.1 成熟dek48突变籽粒的表型分析
对成熟的M4代分离果穗进行观察发现, 与野生型籽粒相比dek48籽粒皱缩扁小, 胚乳填充不饱满, 种皮灰白(图1-A)。对突变籽粒纵切发现胚和胚乳均表现出严重发育缺陷(图1-C)。将该突变体与B73、C7-2和Mo17分别杂交构建F2代分离群体, 发现不同遗传背景下dek48突变籽粒的胚乳缺陷程度有所不同, 其中具有Mo17背景的突变籽粒空瘪程度最为严重(图1-D~F)。百粒重比较结果表明, 不同遗传背景下dek48的突变籽粒的百粒重仅为正常籽粒的10%~17% (图2), 突变籽粒重量显著降低。进一步对突变籽粒和正常籽粒的组分含量进行测定, dek48突变籽粒中淀粉含量显著降低, 蛋白含量显著减少, 油分含量无显著差异(图3)。扫描电镜观察发现, 正常籽粒淀粉粒大而饱满, 排列有序, 基质蛋白位于颗粒缝隙间(图4-A); 突变籽粒淀粉粒显著变小且排列致密, 表皮疏松多孔, 基质蛋白增多(图4-B)。此外通过卷纸发芽试验发现, dek48突变籽粒不能发芽。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1玉米dek48突变体表型
A: 成熟M4代果穗; B: 野生型籽粒纵切面; C: dek48突变籽粒纵切面; D: 以B73为母本的F2果穗; E: 以Mo17为母本的F2果穗; F: C7-2为母本的F2果穗。
Fig. 1Phenotype of dek48 mutant in maize
A: mature M4 ear; B: longitudinal section of wild type kernel; C: longitudinal section of dek48 kernel; D: F2 ear with B73 as the maternal parent; E: F2 ear with Mo17 as the maternal parent; F: F2 ear with C7-2 as the maternal parent.、
图2
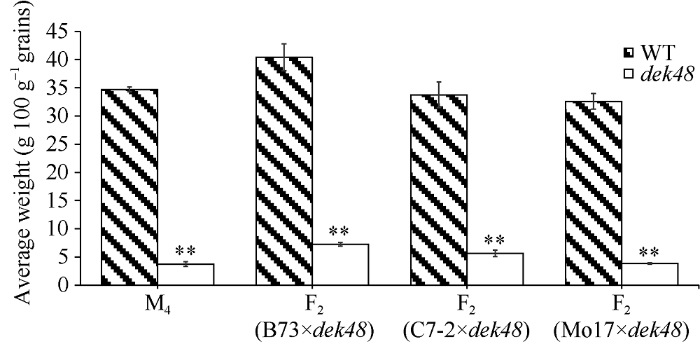 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2正常籽粒和突变籽粒的百粒重
**分别表示在0.01水平差异显著。
Fig. 2Hundred-kernel weight of wild type and mutant kernels
** Significant difference at P< 0.01.
图3
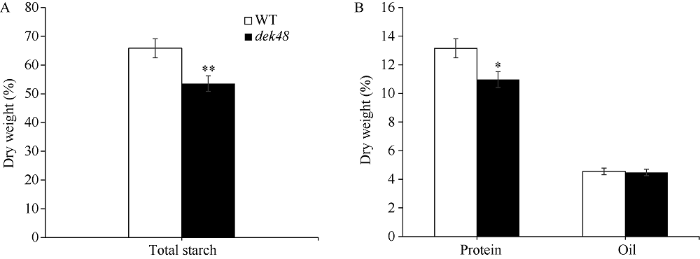 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3WT和dek48籽粒成分测定分析
A: 野生型和dek48的淀粉含量; B: 野生型和dek48的蛋白和油分含量。*, **分别表示野生型与突变体在0.05和0.01水平差异显著。
Fig. 3Determination and analysis of WT and dek48 kernel components
A: starch content of WT and dek48; B: protein and oil content of WT and dek48. *, ** represent significant difference between the dek48 mutant and WT at the 0.05 and 0.01 probability levels, respectively.
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4WT和dek48胚乳的扫描电镜观察
A: WT扫描电镜观察, bar = 10 μm; B: dek48扫描电镜观察, bar = 10 μm; SG: 淀粉粒; MP: 基质蛋白。
Fig. 4SEM observation of the endosperm of WT and dek48
A: SEM observation of WT, bar = 10 μm; B: SEM observation of dek48, bar = 10 μm; SG: starch granule; MP: matrix protein.
2.2 不同发育时期dek48胚和胚乳表型分析
在玉米授粉后12 DAP (day after pollination), dek48即可观察到明显的籽粒变小, 胚和胚乳发育缺陷的表型, 表明该突变发生在籽粒发育的早期阶段。20 DAP时, 正常籽粒胚乳呈浅黄色, 突变籽粒胚乳滞缓发育, 颜色发白; 25 DAP时, 正常籽粒饱满, 胚乳呈黄色, 而突变籽粒空瘪皱缩, 胚和胚乳发育不正常, 种皮内有空腔呈灰白色(图5)。进一步对16 DAP的玉米籽粒进行石蜡切片观察发现, 野生型胚已分化出5片叶原基和明显的胚根结构, 而dek48的胚体积略小, 发育滞后, 仅有2片叶原基, 胚根结构不明显。此外与野生型籽粒相比dek48胚乳细胞变小, 淀粉粒数量减少, 糊粉层细胞结构不规则且层数增加, 种皮与胚乳有空腔(图6)。以上结果表明DEK48基因功能丧失后, 严重影响了玉米籽粒胚和胚乳的生长发育进程及胚乳淀粉细胞结构。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5授粉后不同时期胚和胚乳观察分析
A: 突变型籽粒(dek48)在12、16、20、25、30 DAP胚和胚乳及完整种子的观察分析, bar = 2 mm; B: 野生型籽粒(WT)在12、16、20、25、30 DAP胚和胚乳及完整种子的观察分析, bar = 2 mm。
Fig. 5Observation of embryo and endosperm of WT and dek48 at different development stages
A: the observation and analysis of the embryo, endosperm and whole seed of dek48 at 12, 16, 20, 25, and 30 DAP, bar = 2 mm; B: the observation and analysis of the embryo, endosperm and whole seed of WT at 12, 16, 20, 25, and 30 DAP, bar = 2 mm.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6WT和dek48籽粒在16DAP时的石蜡切片观察
A: 授粉后16 d WT籽粒的胚, bar = 200 μm; B: 授粉后16 d dek48籽粒的胚, bar = 200 μm; C: 授粉后16 d WT籽粒的糊粉层, bar = 50 μm; D: 授粉后16 d dek48籽粒的糊粉层, bar = 50 μm; AL: 糊粉层。
Fig. 6Paraffin section observation of WT and dek48 kernels at 16DAP
A: embryos of WT seeds at 16 DAP, bar = 200 μm; B: embryo of dek48 seeds at 16 DAP, bar = 200 μm; C: aleurone layer of WT seeds at 16 DAP, bar = 50 μm; D: aleurone layer of dek48 seeds at 16 DAP, bar = 50 μm; AL: aleurone.
2.3 dek48突变性状由隐性单基因控制
随机挑选M4代和3种不同遗传背景的F2代分离果穗, 对正常籽粒和突变籽粒的分离比进行卡方(χ2)测验, 结果表明正常籽粒和突变籽粒符合3︰1孟德尔分离定律, 证明dek48的性状受隐性单基因控制(表2)。Table 2
表2
表2野生型与dek48突变籽粒分离比统计分析
Table 2
| 总粒数 Total kernel number | 突变体籽粒/野生型籽粒 dek48/WT | χ2 | |
|---|---|---|---|
| M4 | 515 | 108/407 | 1.20 |
| F2 (B73×dek48) | 1019 | 236/783 | 1.84 |
| F2 (C7-2×dek48) | 1067 | 259/808 | 0.30 |
| F2 (Mo17×dek48) | 1401 | 326/1075 | 2.24 |
新窗口打开|下载CSV
2.4 DEK48基因的定位
在具有B73遗传背景的F2代群体中挑选出48个突变体籽粒, 使用CTAB法[22]提取的DNA利用20 K GBTS技术分析样品基因型。数据分析结果表明F2亲本间存在多态性的SNP标记数量为6864个, 占总位点数的34.2%。平均每条染色体上SNP标记数目为686个, 其中1号染色体最多为1082个(图7)。分析所有多态性SNP位点对应的SNP-index在全基因组水平的变化, 发现在3号染色体上3.55 Mb~19.25 Mb区间可能与目标性状连锁(图8)。增加群体个数到232, 同时开发6个InDel标记(In6.38、In7.29、In7.39、In8.29、In8.53、In9.06)将基因DEK48定位到3号染色体7.39 Mb~8.29 Mb区间内。图7
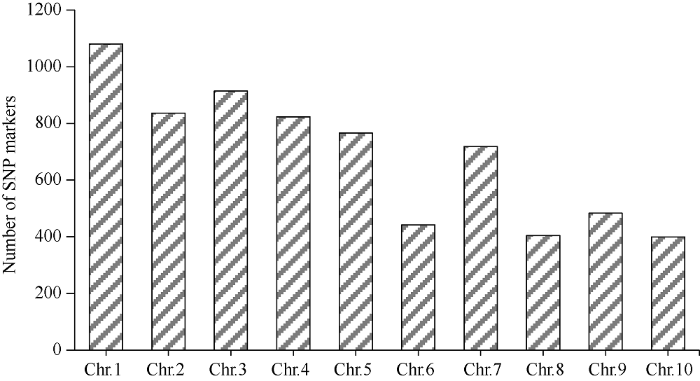 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7多态性SNP标记的数目
Fig. 7Number of SNP markers
图8
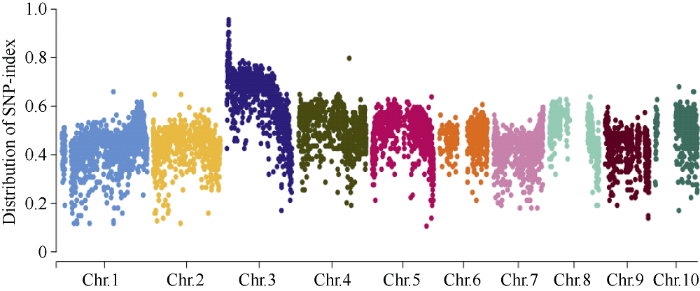 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8SNP-index全基因组频率分布图
Fig. 8Distribution of SNP-index on whole genome
扩大定位群体至2300, 用In7.39标记筛选出基因DEK48左侧交换单株42个, 用In8.29标记筛选出基因DEK48右侧交换单株50个。开发7个SNP标记, 利用KASP技术对这些交换单株进行基因分型, 发现在7.52 Mb处右侧交换单株减少到13个(图9)。因此进一步将定位区间缩短在7.39 Mb~7.52 Mb之间, 两侧相距130 kb。利用Gramene (
图9
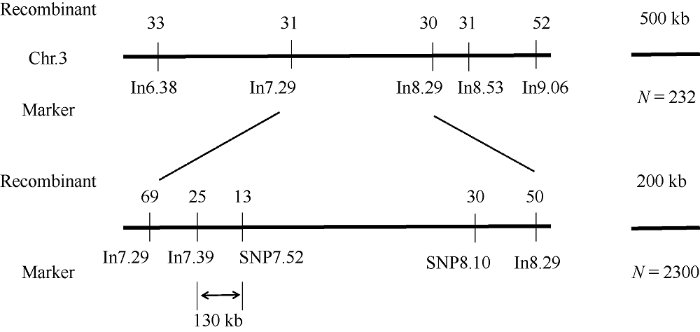 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9dek48突变体的精细定位
N: 群体大小; Recombinant: 重组单株数。
Fig. 9The fine mapping of dek48 mutant
N: number of population; Recombinant: number of recombinants.
Table 3
表3
表3dek48定位区间的候选基因注释
Table 3
| 基因位点 Gene locus | 基因位置 Gene location | 基因注释 Gene annotation |
|---|---|---|
| Zm00001d039532 | Chr. 3: 7,399,394-7,401,753 | WRKY-transcription factor 56 |
| Zm00001d039533 | Chr. 3: 7,446,385-7,450,083 | Plant cysteine oxidase 2 |
| Zm00001d039534 | Chr. 3: 7,451,813-7,457,371 | F-box family protein |
| Zm00001d039535 | Chr. 3: 7,479,692-7,482,979 | HXXXD-type acyl-transferase family protein |
| Zm00001d039536 | Chr. 3: 7,484,018-7,486,445 | Syntaxin22 |
| Zm00001d039537 | Chr. 3: 7,487,245-7,530,464 | Protein ALWAYS EARLY 3 |
新窗口打开|下载CSV
3 讨论
胚与胚乳的早期发育是一个复杂的生物学过程, 对于玉米籽粒形态建成具有重大影响。以籽粒突变体为材料, 挖掘相关的调控基因, 对揭示籽粒早期发育的分子机制以及玉米产量品质的分子改良具有重要意义。我们在郑58 EMS诱变突变体库中分离得到隐性遗传的籽粒缺陷突变体dek48。dek48在12 DAP时能被明确识别, 比野生型籽粒更小, 颜色更白, 胚乳质地更柔软; 成熟的dek48籽粒被观察到皱缩扁小, 种皮灰白, 胚和胚乳发育异常, 百粒重显著降低。在目前已报道的dek突变体中, 如dek10、dek35、dek36、dek37、dek39、dek42[10-14,20]尚有部分突变籽粒可以萌发, 但幼苗发育滞缓不能成活。而dek48突变籽粒则与dek15、dek36、dek41[12,16-17]等类似, 完全不能萌发, 为胚致死突变。与dek15[17]、dek37[13]、dek44[21]一样, dek48的总淀粉和总蛋白含量显著降低, 扫描电镜观察发现dek48淀粉粒显著变小, 结构疏松, 胚乳发育严重受阻。
对16 DAP的突变籽粒切片观察发现, 与野生型籽粒相比, dek48胚乳糊粉层细胞发育不规则, 细胞形状非立方体, 大小不均, 部分区域层数增加。表明DEK48基因很可能与糊粉层细胞发育有关。糊粉层位于玉米胚乳的最外层, 是一层含有大量糊粉粒的立方形厚壁细胞[26]。当种子萌发时, 糊粉层受到激素刺激产生许多代谢酶, 分解胚乳中营养物质提供用于萌发的能量[27]。
在已克隆的dek突变体中, DEK1是糊粉层发育的关键基因, 它参与了糊粉层细胞特化及其命运维持; 是一个具有21个跨膜区域, 一个细胞外多肽环和一个半胱氨酸蛋白酶结构域的质膜蛋白[28]。dek1突变体表现为糊粉层细胞缺失, 胚败育, 胚乳因缺乏醇溶蛋白而呈现粉质表型[28,29]。此外, CR4[30]、SAL1[31]、THK1[32]等基因也被证明与糊粉层细胞特化有关。CR4和DEK1参与了糊粉层细胞特化的信号接收与传导, SAL1负调控CR4和DEK1[30], THK1位于DEK1基因下游对糊粉层细胞特化进行负调控[32]。尽管这些基因已被克隆研究, 但糊粉层细胞发育分化的分子机制仍不清楚。我们将DEK48基因定位在了玉米3号染色体7.39 Mb~7.52 Mb区域内。现已研究报道的dek突变体多分布在4号 (dek10、dek15、dek42、dek44[10,17,20-21])和5号(dek33、dek36、dek38[12,18-19])染色体中, 且多数为PPR蛋白调控。在3号染色体候选区间内尚无已克隆的基因, 证明DEK48可能是一个参与糊粉层细胞发育的新基因。
在DEK48基因的预测区间范围内, 有6个候选基因。其中Zm00001d039532编码转录因子WRKY56。WRKY基因广泛参与植物的生长发育调控[33], 以及对生物胁迫[34]和对非生物逆境的响应[35]。此外部分拟南芥和水稻中的WRKY基因也被报道参与了籽粒的生长发育[36]。Zm00001d039533编码植物半胱氨酸氧化酶PCO2, PCO参与感知植物细胞内氧浓度变化及植物低氧应答响应[37]。Zm00001d039534的表达产物是一种F-box家族蛋白。F-box家族蛋白是SCF复合体中泛素连接酶E3的重要组分, 在植物生长发育、激素信号转导、应对逆境胁迫中均发挥重要的调节作用[38]。Zm00001d039535编码AX-阿魏酰转移酶, 该酶可以将阿魏酰基转移到阿拉伯木聚糖或UDP-阿拉伯木聚糖, 参与植物细胞壁木质素形成[39,40]。Zm00001d039536的表达产物是Syntaxin22, 在植物中, Syntaxin蛋白能通过膜泡运输参与植物细胞板形成[41]、与离子通道蛋白相互作用、植物的抗病性及植物的向重力性应答等众多生理过程[42]。Zm00001d 039537的表达产物是蛋白ALWAYS EARLY 3, 参与细胞循环过程。根据以上分析, Zm00001d039532、Zm00001d039534基因所编码蛋白可能参与籽粒的发育调控。后续将通过基因表达分析和基因组测序来进一步确定候选基因。
4 结论
本研究发现了一个新的隐性单基因控制的玉米籽粒缺陷突变体dek48。该突变体胚和胚乳发育严重受阻, 糊粉层细胞形态结构不规则。利用图位克隆策略将其定位在玉米3号染色体7.39 Mb~7.52 Mb之间。其中暂未发现已克隆的基因, 表明是DEK48可能是一个调控籽粒发育的新基因。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
URLPMID:17249053 [本文引用: 1]

A planting of 3,919 M(1) kernels from normal ears crossed by EMS-treated pollen produced 3,461 M(1) plants and 3,172 selfed ears. These plants yielded 2,477 (72%) total heritable changes; the selfed ears yielded 2,457 (78%) recessive mutants, including 855 (27%) recessive kernel mutants and 8 (0.23%) viable dominant mutants. The ratio of recessive to dominant mutants was 201:1. The average mutation frequency for four known loci was three per 3,172 genomes analyzed. The estimated total number of loci mutated was 535 and the estimated number of kernel mutant loci mutated was 285. Among the 855 kernel mutants, 432 had a nonviable embryo, and 59 germinated but had a lethal seedling. A sample of 194 of the latter two types was tested for heritability, lethality, chromosome arm location and endosperm-embryo interaction between mutant and nonmutant tissues in special hyper-hypoploid combinations produced by manipulation of B-A translocations. The selected 194 mutants were characterized and catalogued according to endosperm phenotype and investigated to determine their effects on the morphology and development of the associated embryo. The possibility of rescuing some of the lethal mutants by covering the mutant embryo with a normal endosperm was investigated. Ninety of these 194 mutants were located on 17 of the 18 chromosome arms tested. Nineteen of the located mutants were examined to determine the effect of having a normal embryo in the same kernel with a mutant endosperm, and vice versa, as compared to the expression observed in kernels with both embryo and endosperm in a mutant condition. In the first situation, for three of the 19 mutants, the mutant endosperm was less extreme (the embryo helped); for seven cases, the mutant endosperm was more extreme (the embryo hindered); and for nine cases, there was no change. In the reverse situation, for four cases the normal endosperm helped the mutant embryo; for 14 cases there was no change and one case was inconclusive.
URLPMID:17246478 [本文引用: 1]
We have examined the effects on embryonic and gametophytic development of two nonallelic defective-kernel mutants of maize. Earlier studies indicated that both mutants are abnormal in embryonic morphogenesis as well as in the formation of their endosperm. Mutant rgh*-1210 embryos depart from the normal embryogenic pathway at the proembryo and transition stage, by developing meristematic lobes and losing bilateral symmetry. They continue growth as irregular cell masses that enlarge and become necrotic. Somatic embryos arising in rgh*-1210 callus cultures display the rgh*-1210 mutant phenotype. Mutant fl*-1253B embryos are variably blocked from the coleoptilar stage through stage 2. Following formation of the shoot apex in the mutant embryos the leaf primordia and tissues surrounding the embryonic axis continue growth and cell division, while the scutellum ceases development and becomes hypertrophied. Mutant fl*-1253B embryos are unable to germinate, either in mutant kernels or as immature embryos in culture, and the mutant scutellar tissue does not produce regenerable callus. Expression of the fl*-1253B locus during male gametophytic development is revealed by a marked reduction in pollen transmission as a result of mutant expression during the interval between meiosis and the initiation of pollen tube growth. In both mutants, there is considerable proliferation of the aleurone cells of the endosperm. Mutant expression of rgh*-1210 in the female gametophyte is revealed by the abnormal antipodal cells of the embryo sac. These results show that these two gene loci play unique and crucial roles in normal morphogenesis of the embryo. In addition, it is evident that both mutants are pleiotropic in affecting the development of the endosperm and gametophyte as well as the embryo. These pleiotropisms suggest some commonality in the gene regulation of development in these three tissues.
URLPMID:17249054 [本文引用: 1]
This report presents the initial results of our study of the immature kernel stage of 150 defective kernel maize mutants. They are single gene, recessive mutants that map throughout the genome, defective in both endosperm and embryo development and, for the most part, lethal (Neuffer and Sheridan 1980). All can be distinguished on immature ears, and 85% of them reveal a mutant phenotype within 11 to 17 days post-pollination. Most have immature kernels that are smaller and lighter in color than their normal counterparts. Forty of the mutants suffer from their defects early in kernel development and are blocked in embryogenesis before their primordia differentiate, or, if primordia are formed, they are unable to germinate when cultured as immature embryos or tested at maturity; a few begin embryo degeneration prior to the time that mutant kernels became visually distinguishable. The others express the associated lesion later in kernel development and form at least one leaf primordium by the time kernels are distinguishable and will germinate when cultured or tested at maturity. In most cases, on a fresh weight basis, the mutants have embryos that are more severely defective than the endosperm; their embryos usually are no more than one-half to two-thirds the size, and lag behind by one or two developmental stages. in comparison with embryos in normal kernels from the same ear. One hundred and two mutants were examined by culturing embryos on basal and enriched media; 21 simply enlarged or completely failed to grow on any of the media tested; and 81 produced shoots and roots on at least one medium. Many grew equally well on basal and enriched media; 16 grew at a faster rate on basal medium and 23 displayed a superior growth on enriched medium. Among the latter group, 10 may be auxotrophs. One of these mutants and another mutant isolated by E. H. Coe are proline-requiring mutants, allelic to pro-1. Considering their diversity of expression as evidenced by their differences in morphological appearance, degree of defectiveness and response to embryo culturing, we believe that they represent many different gene loci.
[本文引用: 1]
URLPMID:15269332 [本文引用: 1]
[本文引用: 1]
URLPMID:22773745 [本文引用: 1]
DOI:10.1534/genetics.116.196105URLPMID:27815362 [本文引用: 2]
In flowering plants, many respiration-related proteins are encoded by the mitochondrial genome and the splicing of mitochondrion-encoded messenger RNA (mRNA) involves a complex collaboration with nuclear-encoded proteins. Pentatricopeptide repeat (PPR) proteins have been implicated in these RNA-protein interactions. Maize defective kernel 2 (dek2) is a classic mutant with small kernels and delayed development. Through positional cloning and allelic confirmation, we found Dek2 encodes a novel P-type PPR protein that targets mitochondria. Mitochondrial transcript analysis indicated that dek2 mutation causes reduced splicing efficiency of mitochondrial nad1 intron 1. Mitochondrial complex analysis in dek2 immature kernels showed severe deficiency of complex I assembly. Dramatically up-regulated expression of alternative oxidases (AOXs), transcriptome data, and TEM analysis results revealed that proper splicing of nad1 is critical for mitochondrial functions and inner cristaes morphology. This study indicated that Dek2 is a new PPR protein that affects the splicing of mitochondrial nad1 intron 1 and is required for mitochondrial function and kernel development.
DOI:10.1534/genetics.116.199331URLPMID:28213476 [本文引用: 4]
Respiration, the core of mitochondrial metabolism, depends on the function of five respiratory complexes. Many respiratory chain-related proteins are encoded by the mitochondrial genome and their RNAs undergo post-transcriptional modifications by nuclear genome-expressed factors, including pentatricopeptide repeat (PPR) proteins. Maize defective kernel 10 (dek10) is a classic mutant with small kernels and delayed development. Through positional cloning, we found that Dek10 encodes an E-subgroup PPR protein localized in mitochondria. Sequencing analysis indicated that Dek10 is responsible for the C-to-U editing at nad3-61, nad3-62, and cox2-550 sites, which are specific editing sites in monocots. The defects of these editing sites result in significant reduction of Nad3 and the loss of Cox2. Interestingly, the assembly of complex I was not reduced, but its NADH dehydrogenase activity was greatly decreased. The assembly of complex IV was significantly reduced. Transcriptome and transmission electron microscopy (TEM) analysis revealed that proper editing of nad3 and cox2 is critical for mitochondrial functions, biogenesis, and morphology. These results indicate that the E-subgroup PPR protein Dek10 is responsible for multiple editing sites in nad3 and cox2, which are essential for mitochondrial functions and plant development in maize.
[本文引用: 2]
DOI:10.1111/nph.14507URLPMID:28277611 [本文引用: 4]
Mitochondria are semi-autonomous organelles that are the powerhouse of the cells. Plant mitochondrial RNA editing guided by pentatricopeptide repeat (PPR) proteins is essential for energy production. We identify a maize defective kernel mutant dek36, which produces small and collapsed kernels, leading to embryos and/or seedlings lethality. Seed filling in dek36 is drastically impaired, in line with the defects observed in the organization of endosperm transfer tissue. Positional cloning reveals that DEK36, encoding a mitochondria-targeted E+ subgroup PPR protein, is required for mitochondrial RNA editing at atp4-59, nad7-383 and ccmFN -302, thus resulting in decreased activities of mitochondrial complex I, complex III and complex IV in dek36. Loss-of-function of its Arabidopsis ortholog At DEK36 causes arrested embryo and endosperm development, leading to embryo lethality. At_dek36 also has RNA editing defects in atp4, nad7, ccmFN1 and ccmFN2 , but at the nonconserved sites. Importantly, efficiency of all editing sites in ccmFN1 , ccmFN2 and rps12 is severely decreased in At_dek36, probably caused by the impairment of their RNA stabilization. These results suggest that the DEK36 orthologue pair are essential for embryo and endosperm development in both maize and Arabidopsis, but through divergent function in regulating RNA metabolism of their mitochondrial targets.
URLPMID:29301905 [本文引用: 3]
DOI:10.1111/jipb.12602URLPMID:28981206 [本文引用: 3]
RNA editing is a posttranscriptional process that is important in mitochondria and plastids of higher plants. All RNA editing-specific trans-factors reported so far belong to PLS-class of pentatricopeptide repeat (PPR) proteins. Here, we report the map-based cloning and molecular characterization of a defective kernel mutant dek39 in maize. Loss of Dek39 function leads to delayed embryogenesis and endosperm development, reduced kernel size, and seedling lethality. Dek39 encodes an E sub-class PPR protein that targets to both mitochondria and chloroplasts, and is involved in RNA editing in mitochondrial NADH dehydrogenase3 (nad3) at nad3-247 and nad3-275. C-to-U editing of nad3-275 is not conserved and even lost in Arabidopsis, consistent with the idea that no close DEK39 homologs are present in Arabidopsis. However, the amino acids generated by editing nad3-247 and nad3-275 are highly conserved in many other plant species, and the reductions of editing at these two sites decrease the activity of mitochondria NADH dehydrogenase complex I, indicating that the alteration of amino acid sequence is necessary for Nad3 function. Our results indicate that Dek39 encodes an E sub-class PPR protein that is involved in RNA editing of multiple sites and is necessary for seed development of maize.
[本文引用: 2]
[本文引用: 3]
DOI:10.1105/tpc.18.00921URLPMID:30705131 [本文引用: 4]
Cohesin complexes maintain sister chromatid cohesion to ensure proper chromosome segregation during mitosis and meiosis. In plants, the exact components and functions of the cohesin complex remain poorly understood. Here, we positionally cloned the classic maize (Zea mays) mutant defective kernel 15 (dek15), revealing that it encodes a homolog of SISTER CHROMATID COHESION PROTEIN 4 (SCC4), a loader subunit of the cohesin ring. Developing dek15 kernels contained fewer cells than the wild type, but had a highly variable cell size. The dek15 mutation was found to disrupt the mitotic cell cycle and endoreduplication, resulting in a reduced endosperm and embryo lethality. The cells in the dek15 endosperm and embryo exhibited precocious sister chromatid separation and other chromosome segregation errors, including misaligned chromosomes, lagging chromosomes, and micronuclei, resulting in a high percentage of aneuploid cells. The loss of Dek15/Scc4 function upregulated the expression of genes involved in cell cycle progression and stress responses, and downregulated key genes involved in organic synthesis during maize endosperm development. Our yeast two-hybrid screen identified the chromatin remodeling proteins chromatin remodeling factor 4, chromatin remodeling complex subunit B (CHB)102, CHB105, and CHB106 as SCC4-interacting proteins, suggesting a possible mechanism by which the cohesin ring is loaded onto chromatin in plant cells. This study revealed biological functions for DEK15/SCC4 in mitotic chromosome segregation and kernel development in maize.
[本文引用: 2]
DOI:10.1073/pnas.1703498114URLPMID:28461460 [本文引用: 2]
We have used the newly engineered transposable element Dsg to tag a gene that gives rise to a defective kernel (dek) phenotype. Dsg requires the autonomous element Ac for transposition. Upon excision, it leaves a short DNA footprint that can create in-frame and frameshift insertions in coding sequences. Therefore, we could create alleles of the tagged gene that confirmed causation of the dek phenotype by the Dsg insertion. The mutation, designated dek38-Dsg, is embryonic lethal, has a defective basal endosperm transfer (BETL) layer, and results in a smaller seed with highly underdeveloped endosperm. The maize dek38 gene encodes a TTI2 (Tel2-interacting protein 2) molecular cochaperone. In yeast and mammals, TTI2 associates with two other cochaperones, TEL2 (Telomere maintenance 2) and TTI1 (Tel2-interacting protein 1), to form the triple T complex that regulates DNA damage response. Therefore, we cloned the maize Tel2 and Tti1 homologs and showed that TEL2 can interact with both TTI1 and TTI2 in yeast two-hybrid assays. The three proteins regulate the cellular levels of phosphatidylinositol 3-kinase-related kinases (PIKKs) and localize to the cytoplasm and the nucleus, consistent with known subcellular locations of PIKKs. dek38-Dsg displays reduced pollen transmission, indicating TTI2's importance in male reproductive cell development.
DOI:10.1111/jipb.12798URLPMID:30839161 [本文引用: 3]
RNA-binding proteins (RBPs) play an important role in post-transcriptional gene regulation. However, the functions of RBPs in plants remain poorly understood. Maize kernel mutant dek42 has small defective kernels and lethal seedlings. Dek42 was cloned by Mutator tag isolation and further confirmed by an independent mutant allele and clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated protein 9 materials. Dek42 encodes an RRM_RBM48 type RNA-binding protein that localizes to the nucleus. Dek42 is constitutively expressed in various maize tissues. The dek42 mutation caused a significant reduction in the accumulation of DEK42 protein in mutant kernels. RNA-seq analysis showed that the dek42 mutation significantly disturbed the expression of thousands of genes during maize kernel development. Sequence analysis also showed that the dek42 mutation significantly changed alternative splicing in expressed genes, which were especially enriched for the U12-type intron-retained type. Yeast two-hybrid screening identified SF3a1 as a DEK42-interacting protein. DEK42 also interacts with the spliceosome component U1-70K. These results suggested that DEK42 participates in the regulation of pre-messenger RNA splicing through its interaction with other spliceosome components. This study showed the function of a newly identified RBP and provided insights into alternative splicing regulation during maize kernel development.
DOI:10.1104/pp.19.00546URLPMID:31182559 [本文引用: 3]
Mitochondrial respiration depends on proteins encoded by the nuclear and mitochondrial genomes. Many respiratory chain-related proteins are encoded by the mitochondrial genome and undergo translation by mitochondrial ribosomes. The newly identified maize (Zea mays) defective kernel44 (dek44) mutant produces small kernels showing embryo-lethal phenotypes. We cloned Dek44 by isolating the Mutator tag that produced the mutation and identified it as encoding a putative 50S ribosomal protein L9. Subcellular fractionation by ultracentrifugation confirmed that DEK44 is a mitochondrial ribosomal protein. DEK44 is highly conserved in monocots and only accumulates in kernels. Transcriptome and reverse transcription quantitative PCR analyses revealed that loss of DEK44 function affects the expression of genes encoding respiratory chain-related proteins from the mitochondrial and nuclear genomes. Blue native-PAGE revealed significantly reduced assembly of respiratory chain complexes in dek44 mutant kernels. Transmission electron microscopy indicated that the biogenesis and morphology of mitochondria were strongly affected in dek44 mutant kernels. Furthermore, DEK44 might regulate cell growth and kernel development via cyclin/cyclin-dependent kinase-mediated activities. This study provides insight into the regulation of kernel development based on mitochondrial ribosomal protein function.
[本文引用: 3]
[本文引用: 3]
[本文引用: 1]
URLPMID:22267009 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1146/annurev.ge.27.120193.001145URLPMID:8122901 [本文引用: 1]
DOI:10.1093/pcp/pcv126URLPMID:26374791 [本文引用: 1]
The cereal aleurone layer plays an important role in seed germination, and reactive oxygen species (ROS) in aleurone layers act as crucial signal molecules in this progression. Recent studies have revealed that epigenetic modification is involved in plant development and seed germination. However, little is known about a possible relationship between histone modification and the ROS signaling pathway in cereal aleurone layers during seed germination. Here, we found that the expression of both histone acetyltransferases (HATs) and histone deacetylases (HDACs) was increased gradually during seed germination, accompanied by an increase in global acetylation levels of histones H3 and H4 in maize aleurone layers. The acetylation was found to be promoted by GA(3) and suppressed by ABA. However, when the HDAC inhibitor trichostatin A (TSA) was used, the increased H3K9ac and H4K5ac level correlated with an inhibition of the germination. These results indicated that the overall histone acetylation in the aleurone layers is not required for germination. Similarly these two hormones, GA(3) and ABA, exerted opposed effects on the expression of the ROS-related gene sodCp. Furthermore, chromatin immunoprecipitation experiments showed that the promoter region of the sodCp gene was hyperacetylated during germination, and this acetylation was promoted by GA(3) and inhibited by both ABA and TSA. These results suggested that GA(3)-mediated expression of the sodCp gene in aleurone layers is associated with histone hyperacetylation on the promoter and coding region of this gene, consequently leading to an accumulation of H(2)O(2) which regulated production of alpha-amylase during seed germination.
DOI:10.1104/pp.111.177725URLPMID:21617032 [本文引用: 2]
The maize (Zea mays) aleurone layer occupies the single outermost layer of the endosperm. The defective kernel1 (dek1) gene is a central regulator required for aleurone cell fate specification. dek1 mutants have pleiotropic phenotypes including lack of aleurone cells, aborted embryos, carotenoid deficiency, and a soft, floury endosperm deficient in zeins. Here we describe the thick aleurone1 (thk1) mutant that defines a novel negative function in the regulation of aleurone differentiation. Mutants possess multiple layers of aleurone cells as well as aborted embryos. Clonal sectors of thk1 mutant tissue in otherwise normal endosperm showed localized expression of the phenotype with sharp boundaries, indicating a localized cellular function for the gene. Sectors in leaves showed expanded epidermal cell morphology but the mutant epidermis generally remained in a single cell layer. Double mutant analysis indicated that the thk1 mutant is epistatic to dek1 for several aspects of the pleiotropic dek1 phenotype. dek1 mutant endosperm that was mosaic for thk1 mutant sectors showed localized patches of multilayered aleurone. Localized sectors were surrounded by halos of carotenoid pigments and double mutant kernels had restored zein profiles. In sum, loss of thk1 function restored the ability of dek1 mutant endosperm to accumulate carotenoids and zeins and to differentiate aleurone. Therefore the thk1 mutation defines a negative regulator that functions downstream of dek1 in the signaling system that controls aleurone specification and other aspects of endosperm development. The thk1 mutation was found to be caused by a deletion of approximately 2 megabases.
DOI:10.1105/tpc.106.048868URLPMID:17933905 [本文引用: 1]
DEFECTIVE KERNEL1 (DEK1), which consists of a membrane-spanning region (DEK1-MEM) and a calpain-like Cys proteinase region (DEK1-CALP), is essential for aleurone cell formation at the surface of maize (Zea mays) endosperm. Immunolocalization and FM4-64 dye incubation experiments showed that DEK1 and CRINKLY4 (CR4), a receptor kinase implicated in aleurone cell fate specification, colocalized to plasma membrane and endosomes. SUPERNUMERARY ALEURONE LAYER1 (SAL1), a negative regulator of aleurone cell fate encoding a class E vacuolar sorting protein, colocalized with DEK1 and CR4 in endosomes. Immunogold localization, dual-axis electron tomography, and diffusion of fluorescent dye tracers showed that young aleurone cells established symplastic subdomains through plasmodesmata of larger dimensions than those connecting starchy endosperm cells and that CR4 preferentially associated with plasmodesmata between aleurone cells. Genetic complementation experiments showed that DEK1-CALP failed to restore wild-type phenotypes in maize and Arabidopsis thaliana dek1 mutants, and DEK1-MEM also failed to restore wild-type phenotypes in Arabidopsis dek1-1 mutants. Instead, ectopic expression of DEK1-MEM under the control of the cauliflower mosaic virus 35S promoter gave a dominant negative phenotype. These data suggest a model for aleurone cell fate specification in which DEK1 perceives and/or transmits a positional signal, CR4 promotes the lateral movement of aleurone signaling molecules between aleurone cells, and SAL1 maintains the proper plasma membrane concentration of DEK1 and CR4 proteins via endosome-mediated recycling/degradation.
DOI:10.1126/science.273.5280.1406URLPMID:8703079 [本文引用: 2]
The maize crinkly4 (cr4) mutation affects leaf epidermis differentiation such that cell size and morphology are altered, and surface functions are compromised, allowing graft-like fusions between organs. In the seed, loss of cr4 inhibits aleurone formation in a pattern that reflects the normal progression of differentiation over the developing endosperm surface. The cr4 gene was isolated by transposon tagging and found to encode a putative receptor kinase. The extracellular domain contains a cysteine-rich region similar to the ligand binding domain in mammalian tumor necrosis factor receptors (TNFRs) and seven copies of a previously unknown 39-amino acid repeat. The results suggest a role for cr4 in a differentiation signal.
DOI:10.1073/pnas.0732023100URLPMID:12750475 [本文引用: 1]
A microscopy-based screen of a large collection of maize Mutator (Mu) transposon lines identified the supernumerary aleurone layers 1-1 (sal1-1) mutant line carrying up to seven layers of aleurone cells in defective kernel endosperm compared with only a single layer in wild-type grains. Normal, well filled endosperm that is homozygous for the sal1-1 mutant allele contains two to three layers of aleurone cells. Cloning of the sal1 gene was accomplished by using Mu tagging, and the identity of the cloned gene was confirmed by isolating an independent sal1-2 allele by reverse genetics. Homozygous sal1-2 endosperm has two to three layers of aleurone cells in normal, well filled grains. In situ hybridization experiments reveal that the sal1 gene is ubiquitously expressed in vegetative as well as zygotic grain tissues, with no difference being detected between aleurone cells and starchy endosperm cells. Northern blot analysis failed to detect the sal1-2 transcript in leaves of homozygous plants, suggesting that the allele is a true sal1 knockout allele. The sal1 gene encodes a homologue of the human Chmp1 gene, a member of the conserved family of the class E vacuolar protein sorting genes implicated in membrane vesicle trafficking. In mammals, CHMP1 functions in the pathway targeting plasma membrane receptors and ligands to lysosomes for proteolytic degradation. Possible roles for the function of the sal1 gene in aleurone signaling, including a defect in endosome trafficking, are discussed.
DOI:10.1104/pp.111.177725URLPMID:21617032 [本文引用: 2]
The maize (Zea mays) aleurone layer occupies the single outermost layer of the endosperm. The defective kernel1 (dek1) gene is a central regulator required for aleurone cell fate specification. dek1 mutants have pleiotropic phenotypes including lack of aleurone cells, aborted embryos, carotenoid deficiency, and a soft, floury endosperm deficient in zeins. Here we describe the thick aleurone1 (thk1) mutant that defines a novel negative function in the regulation of aleurone differentiation. Mutants possess multiple layers of aleurone cells as well as aborted embryos. Clonal sectors of thk1 mutant tissue in otherwise normal endosperm showed localized expression of the phenotype with sharp boundaries, indicating a localized cellular function for the gene. Sectors in leaves showed expanded epidermal cell morphology but the mutant epidermis generally remained in a single cell layer. Double mutant analysis indicated that the thk1 mutant is epistatic to dek1 for several aspects of the pleiotropic dek1 phenotype. dek1 mutant endosperm that was mosaic for thk1 mutant sectors showed localized patches of multilayered aleurone. Localized sectors were surrounded by halos of carotenoid pigments and double mutant kernels had restored zein profiles. In sum, loss of thk1 function restored the ability of dek1 mutant endosperm to accumulate carotenoids and zeins and to differentiate aleurone. Therefore the thk1 mutation defines a negative regulator that functions downstream of dek1 in the signaling system that controls aleurone specification and other aspects of endosperm development. The thk1 mutation was found to be caused by a deletion of approximately 2 megabases.
DOI:10.1105/tpc.15.00829URLPMID:26578700 [本文引用: 1]
Plant shoot branching is pivotal for developmental plasticity and crop yield. The formation of branch meristems is regulated by several key transcription factors including REGULATOR OF AXILLARY MERISTEMS1 (RAX1), RAX2, and RAX3. However, the regulatory network of shoot branching is still largely unknown. Here, we report the identification of EXCESSIVE BRANCHES1 (EXB1), which affects axillary meristem (AM) initiation and bud activity. Overexpression of EXB1 in the gain-of-function mutant exb1-D leads to severe bushy and dwarf phenotypes, which result from excessive AM initiation and elevated bud activities. EXB1 encodes the WRKY transcription factor WRKY71, which has demonstrated transactivation activities. Disruption of WRKY71/EXB1 by chimeric repressor silencing technology leads to fewer branches, indicating that EXB1 plays important roles in the control of shoot branching. We demonstrate that EXB1 controls AM initiation by positively regulating the transcription of RAX1, RAX2, and RAX3. Disruption of the RAX genes partially rescues the branching phenotype caused by EXB1 overexpression. We further show that EXB1 also regulates auxin homeostasis in control of shoot branching. Our data demonstrate that EXB1 plays pivotal roles in shoot branching by regulating both transcription of RAX genes and auxin pathways.
DOI:10.1016/j.cell.2015.04.024URLPMID:26000484 [本文引用: 1]
Defense against pathogens in multicellular eukaryotes depends on intracellular immune receptors, yet surveillance by these receptors is poorly understood. Several plant nucleotide-binding, leucine-rich repeat (NB-LRR) immune receptors carry fusions with other protein domains. The Arabidopsis RRS1-R NB-LRR protein carries a C-terminal WRKY DNA binding domain and forms a receptor complex with RPS4, another NB-LRR protein. This complex detects the bacterial effectors AvrRps4 or PopP2 and then activates defense. Both bacterial proteins interact with the RRS1 WRKY domain, and PopP2 acetylates lysines to block DNA binding. PopP2 and AvrRps4 interact with other WRKY domain-containing proteins, suggesting these effectors interfere with WRKY transcription factor-dependent defense, and RPS4/RRS1 has integrated a
DOI:10.1016/j.bbrc.2015.07.085URLPMID:26196744 [本文引用: 1]
WRKY transcription factors play an important role in cold defense of plants. However, little information is available about the cold-responsive WRKYs in tomato (Solanum lycopersicum). In the present study, a complete characterization of this gene family was described. Eighty WRKY genes in the tomato genome were identified. Almost all WRKY genes contain putative stress-responsive cis-elements in their promoter regions. Segmental duplications contributed significantly to the expansion of the SlWRKY gene family. Transcriptional analysis revealed notable differential expression in tomato tissues and expression patterns under cold stress, which indicated wide functional divergence in this family. Ten WRKYs in tomato were strongly induced more than 2-fold during cold stress. These genes represented candidate genes for future functional analysis of WRKYs involved in the cold-related signal pathways. Our data provide valuable information about tomato WRKY proteins and form a foundation for future studies of these proteins, especially for those that play an important role in response to cold stress.
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/ncomms4425URLPMID:24599061 [本文引用: 1]
In plant and animal cells, amino-terminal cysteine oxidation controls selective proteolysis via an oxygen-dependent branch of the N-end rule pathway. It remains unknown how the N-terminal cysteine is specifically oxidized. Here we identify plant cysteine oxidase (PCO) enzymes that oxidize the penultimate cysteine of ERF-VII transcription factors by using oxygen as a co-substrate, thereby controlling the lifetime of these proteins. Consequently, ERF-VII proteins are stabilized under hypoxia and activate the molecular response to low oxygen while the expression of anaerobic genes is repressed in air. Members of the PCO family are themselves targets of ERF-VII transcription factors, generating a feedback loop that adapts the stress response according to the extent of the hypoxic condition. Our results reveal that PCOs act as sensor proteins for oxygen in plants and provide an example of how proactive regulation of the N-end rule pathway balances stress response to optimal growth and development in plants.
[本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.106.094995URLPMID:17351055 [本文引用: 1]
Arabinoxylans (AXs) are major components of graminaceous plant cell walls, including those in the grain and straw of economically important cereals. Despite some recent advances in identifying the genes encoding biosynthetic enzymes for a number of other plant cell wall polysaccharides, the genes encoding enzymes of the final stages of AX synthesis have not been identified. We have therefore adopted a novel bioinformatics approach based on estimation of differential expression of orthologous genes between taxonomic divisions of species. Over 3 million public domain cereal and dicot expressed sequence tags were mapped onto the complete sets of rice (Oryza sativa) and Arabidopsis (Arabidopsis thaliana) genes, respectively. It was assumed that genes in cereals involved in AX biosynthesis would be expressed at high levels and that their orthologs in dicotyledonous plants would be expressed at much lower levels. Considering all rice genes encoding putative glycosyl transferases (GTs) predicted to be integral membrane proteins, genes in the GT43, GT47, and GT61 families emerged as much the strongest candidates. When the search was widened to all other rice or Arabidopsis genes predicted to encode integral membrane proteins, cereal genes in Pfam family PF02458 emerged as candidates for the feruloylation of AX. Our analysis, known activities, and recent findings elsewhere are most consistent with genes in the GT43 families encoding beta-1,4-xylan synthases, genes in the GT47 family encoding xylan alpha-1,2- or alpha-1,3-arabinosyl transferases, and genes in the GT61 family encoding feruloyl-AX beta-1,2-xylosyl transferases.
DOI:10.3389/fpls.2013.00050URLPMID:23508643 [本文引用: 1]
The cell walls of grasses such as wheat, maize, rice, and sugar cane, contain large amounts of ferulate that is ester-linked to the cell wall polysaccharide glucuronoarabinoxylan (GAX). This ferulate is considered to limit the digestibility of polysaccharide in grass biomass as it forms covalent linkages between polysaccharide and lignin components. Candidate genes within a grass-specific clade of the BAHD acyl-coA transferase superfamily have been identified as being responsible for the ester linkage of ferulate to GAX. Manipulation of these BAHD genes may therefore be a biotechnological target for increasing efficiency of conversion of grass biomass into biofuel. Here, we describe the expression of these candidate genes and amounts of bound ferulate from various tissues and developmental stages of the model grass Brachypodium distachyon. BAHD candidate transcripts and significant amounts of bound ferulate were present in every tissue and developmental stage. We hypothesize that BAHD candidate genes similar to the recently described Oryza sativa p-coumarate monolignol transferase (OsPMT) gene (PMT sub-clade) are principally responsible for the bound para-coumaric acid (pCA), and that other BAHD candidates (non-PMT sub-clade) are responsible for bound ferulic acid (FA). There were some similarities with between the ratio of expression non-PMT/PMT genes and the ratio of bound FA/pCA between tissue types, compatible with this hypothesis. However, much further work to modify BAHD genes in grasses and to characterize the heterologously expressed proteins is required to demonstrate their function.
URLPMID:7961911 [本文引用: 1]
Syntaxins are a family of vesicular transport receptors that are involved in membrane traffic through both the constitutive and regulated secretory pathways. Syntaxins 1A/B,2,3, and 4 are principally associated with the plasma membrane. Two of the syntaxins, 1A and 1B, have been suggested to be the docking receptors for synaptic vesicles with the presynaptic membrane. The most distant member of the family, syntaxin 5, has been found in the Golgi region and has significant homology (35% identity) with Sed5p, an essential protein in yeast which is required for vesicular transport from the endoplasmic reticulum (ER) to the Golgi stack. Here we present evidence that syntaxin 5 performs an analogous function in ER to Golgi transport in mammalian cells. Transient expression of an hemagglutinin-tagged full-length clone of syntaxin 5 and a truncated mutant lacking the transmembrane domain inhibited the transport of vesicular stomatitis virus glycoprotein to the Golgi stack. Under these conditions, vesicular stomatitis virus glycoprotein accumulated in pre-Golgi intermediates, which were strongly enriched in syntaxin 5. Our results suggest that syntaxin 5 is the functional mammalian homologue of Sed5p and provides evidence for its role in regulating the potential targeting and/or fusion of carrier vesicles following export from the ER.
DOI:10.1016/j.pbi.2004.09.001URLPMID:15491921 [本文引用: 1]
Gravitropism has attracted much attention from plant biologists. Recent studies have provided molecular evidence supporting two long-surviving hypotheses about the mechanism of gravitropism: the starch-statolith hypothesis and the Cholodney-Went hypothesis. Amyloplast movement along the gravity vector within gravity-sensing cells in the root and shoot is the most likely trigger of subsequent intracellular signaling. Several possible events leading from this signaling to differential auxin distribution within the sensing cells have been suggested recently.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}