 ,1,*
,1,*Overexpression of OsMPK17 protein enhances drought tolerance of rice
MA Jin-Jiao1, LAN Jin-Ping1,2, ZHANG Tong1, CHEN Yue1, GUO Ya-Lu1,3, LIU Yu-Qing1, YAN Gao-Wei1, WEI Jian1, DOU Shi-Juan1, YANG Ming1, LI Li-Yun1, LIU Guo-Zhen,1,*通讯作者:
收稿日期:2019-02-26接受日期:2019-08-9网络出版日期:2019-09-04
| 基金资助: |
Received:2019-02-26Accepted:2019-08-9Online:2019-09-04
| Fund supported: |
作者简介 About authors
E-mail:majinjiao_mbb@126.com。

摘要
关键词:
Abstract
Keywords:
PDF (2806KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
马金姣, 兰金苹, 张彤, 陈悦, 郭亚璐, 刘玉晴, 燕高伟, 魏健, 窦世娟, 杨明, 李莉云, 刘国振. 过表达OsMPK17激酶蛋白质增强了水稻的耐旱性[J]. 作物学报, 2020, 46(1): 20-30. doi:10.3724/SP.J.1006.2020.92007
MA Jin-Jiao, LAN Jin-Ping, ZHANG Tong, CHEN Yue, GUO Ya-Lu, LIU Yu-Qing, YAN Gao-Wei, WEI Jian, DOU Shi-Juan, YANG Ming, LI Li-Yun, LIU Guo-Zhen.
蛋白质激酶可以使特定蛋白质侧链中的丝氨酸、苏氨酸或酪氨酸残基共价磷酸化[1], 它们在真核生物(酵母、哺乳动物、人及植物)中广泛存在[2,3]。促分裂原活化蛋白质激酶(mitogen-activated protein kinase, MAPK)属于丝/苏氨酸蛋白质激酶大家族, 通常由11个保守的亚结构域组成[4]。真核生物中的激酶级联反应一般通过3个磷酸激酶(MAP3K、MAP2K和MAPK)依次使底物蛋白磷酸化转导和放大信号, 调控特定基因的表达[5]。MAPK处于这个级联反应的末端, 它被上游MAP2K活化后再磷酸化下游的元件, 如转录因子WRKY、MYB等, 将信号传递下去。植物MAPK级联途径在多种信号传导过程中发挥着承上启下的作用, 与许多生物及非生物胁迫反应、激素反应、细胞分化和发育过程相互关联, 形成调控植物正常生长和逆境应答的网络。
OsBWMKI是第一个被鉴定的水稻MAPK基因, 受稻瘟病菌(Magnaporthe grisea)侵染和机械损伤诱导表达[6]。OsBWMK1和OsWJUMK1受脱落酸(Abscisic acid, ABA)、高盐、干旱、高温(37°C)或重金属等非生物胁迫的诱导表达, 而紫外线和低温(12°C)不影响OsBWMK1的表达水平, 但高温(37°C)会降低OsWJUMK1的水平[7]。Shi等[8]证明OsMPK1在脱落酸信号途径中能调高抗氧化物酶的活性。Xie等[9]证明OsMPK3能被OsMKK6磷酸化, 二者互作共同介导低温信号的传导[10]。OsMPK3和OsMPK6也参与盐胁迫反应。低温胁迫会启动OsMAPK3-OsbHLH002-OsTPP1信号途径, 提高水稻耐寒性[11]。OsMPK4与OsMKK1二者互作协调控制下游转录因子的表达并介导盐胁迫应答[12]。水稻OsMPK5在褐飞虱抗性反应中发挥作用, 其证据有OsMPK5与褐飞虱抗性基因Bphi008a互作, 且OsMPK5可磷酸化Bphi008a, 在褐飞虱感染后OsMPK5的表达水平发生变化[13]。OsMAPK6参与OsMKKK10-OsMKK4信号通路, 对水稻的籽粒大小和重量发挥调节作用[14]。OsMAP1、OsMPK14和OsMEKl等参与低温胁迫应答反应, 高温和脱落酸处理还能使OsMPK14的表达上调, 干旱和光照也对其有不同程度的影响[15]。OsMAPK15基因提高种子萌发期对干旱和盐胁迫的耐受性[16]。Lee等[17]发现OsMAPK33在水稻盐胁迫中起负调控作用。综上可见, 多个水稻MAPK都在逆境胁迫应答中发挥作用。所以进一步鉴定逆境应答相关的MAPK基因, 探讨水稻逆境胁迫应答机制具有重要的理论意义和应用价值。
目前对水稻MAPK家族基因功能研究的线索主要来自遗传分析及转录数据, 其中OsMPK17基因在逆境胁迫反应中的功能研究尚未见报道。本研究对不同逆境胁迫下的蛋白质样品进行免疫印迹分析, 筛查发现OsMPK17蛋白质在干旱胁迫下被诱导表达, 提示该蛋白质在干旱胁迫应答中有作用。为了调查OsMPK17的功能, 构建了过表达OsMPK17蛋白质的载体, 转化水稻后筛选获得了OsMPK17蛋白质过表达的纯合株系, 在田间表型鉴定的基础上, 通过PEG-6000拟旱处理、幼苗期失水试验和实际土培干旱胁迫试验, 证明过表达OsMPK17蛋白质的转基因水稻生长优于野生型, 该证据表明OsMPK17蛋白质在干旱胁迫应答过程中发挥作用。
1 材料与方法
1.1 供试材料
所用水稻品种为TP309 (粳稻)。克隆用的大肠杆菌菌株为DH5α, 融合蛋白质表达菌株为BL21(DE3) pLysS和Codon Plus。原核表达载体为pET30a和pGST (由pGEM载体改造[18])。pUC57-3HA质粒的插入片段由南京金斯瑞生物科技有限公司合成; pEASY-T1质粒购自生工生物工程(上海)股份有限公司; pUBI-C4300质粒由Pamela Ronald博士(UC Davis, USA)赠送。1.2 水稻OsMPK17基因的克隆
水稻OsMPK17基因全长cDNA质粒AK070644 (Os05g50120)购自日本农业生物资源研究所水稻基因组资源中心(Rice Genome Resource Center, National Institute of Agrobiological Sciences)。利用Primer CE软件[19]设计引物, 由北京华大基因研究中心有限公司合成。上游引物序列为5'-GCGGTACC ATGGGCGGCCGCGCCCGCTC-3', 其中下画线为Kpn I限制性内切酶位点, 下游引物序列为5'-GC GAGCTCGGTTTTCAGTTGAGCAAC-3', 其中下画线为Sac I限制性内切酶位点。以带有全长OsMPK17的cDNA的质粒为模板进行PCR扩增, 对PCR产物和pET30a载体进行双酶切并切胶回收, 将连接后的产物转化到大肠杆菌 DH5α中, 提取重组质粒 DNA, 双酶切验证后送北京华大基因研究中心有限公司测序验证。1.3 融合蛋白质表达纯化
将测序确认正确的pET30a-MPK17质粒转入表达菌Codon plus, 挑取单菌落过夜培养, 按1∶10000的比例转接至100 mL含50 μg mL-1卡那霉素的LB液体培养基中诱导表达, 37°C振荡培养至OD600 为0.6~0.8, 按1∶200比例加入100 mmol L-1的IPTG, 25°C过夜培养, 收菌后用20 mL 10 mmol L-1 Tris-HCl (pH 8.0)将菌液悬浮, 超声破碎(500 W, 60次, 每次10 s, 间隔15 s), 用His-tag beads进行蛋白质纯化, 10% SDS-PAGE分离, 考染检测重组蛋白质的浓度和纯度。1.4 抗体制备
用大肠杆菌表达并纯化后的OsMPK17蛋白质做免疫原, 免疫小鼠制备单克隆抗体, 抗体的制备由北京华大蛋白质研发中心有限公司完成。1.5 水稻总蛋白质提取及免疫印迹(Western blot, WB)检测
将水稻样品装入离心管, 用液氮速冻, 在研磨机(鼎昊源科技, 型号TL2010)上振荡, 1400 r min-1, 30 s, 低温研磨直至粉末状。按照样品和蛋白质提取缓冲液(62.5 mmol L-1 Tris-HCl pH 7.4、10%甘油、2% SDS、1 mmol L-1 PMSF、2 mmol L-1 EDTA、5% β-巯基乙醇) 3∶8比例, 充分混匀后置冰上, 放置10 min, 每隔2 min 剧烈振荡30 s, 12,000 × g, 4°C离心30 min后取上清液即为水稻组织总蛋白质。经SDS-PAGE进行分离, 上样体积一般为10 μL, 电泳完成后将蛋白质转移到PVDF膜上, 按文献[20]描述的过程进行WB检测, 并用抗HSP抗体的检测信号作为标定上样量的内参[21], 二抗为HRP标记的羊抗鼠二抗(北京华大蛋白质研发中心有限公司)。用化学发光成像仪(MiniChemi 610, 北京赛智创业科技有限公司)检测, Image J软件[22]采集信号。1.6 水稻非生物逆境胁迫处理
冷、热、淹和恒光、恒暗等胁迫处理方法及部分样品来自本实验室积累的蛋白质样品资源库RiceS-A300 [23]。1.7 水稻离体叶片激素处理
参照文献[24]进行水稻离体叶片的激素处理。取4~6周龄的水稻幼苗, 剪取长度为2 cm的叶片, 置培养皿中, 加15 mL 100 μmol L-1脱落酸(abscisic acid, ABA)或100 μmol L-1茉莉酸甲酯(methyl jasmonic acid, MeJA), 以灭菌水为对照, 在培养箱中培养, 温度为30°C, 光照周期为L12 h/D12 h, 取材时间点分别为0、6 h、12 h、1 d、2 d、3 d、4 d、5 d和6 d。1.8 过表达转基因载体构建
为了便于后续检测, 以pUC57为骨架, 通过基因合成改造成带3HA和终止子的质粒pUC57-3HA, 在3HA序列上游引入Xba I和Kpn I限制性内切酶位点, 3HA之前有Sac I酶切位点, 之后是终止密码子, 下游是Spe I和Hind III限制性内切酶位点。利用pEASY-T1质粒构建一个中间载体, 将pUC57-3HA中的Xba I与Hind III双酶切片段连接到同样双酶切的pEASY-T1质粒中, 称为pEASY-T1-3HA。将OsMPK17基因的扩增产物用Kpn I和Sac I双酶切, 插入pEASY-T1-3HA, 测序验证后将含有目的基因和3HA的片段用Xba I和Hind III连接到pUBI-C4300质粒中[25]。1.9 水稻转化
通过农杆菌介导法将质粒DNA转入水稻, 在培养基中加甘露糖进行筛选, 遗传转化由武汉伯远生物科技有限公司完成[26,27]。水稻遗传转化的主要步骤为, 从成熟胚诱导愈伤组织, 愈伤组织与根癌农杆菌的共培养, 抗性愈伤组织筛选, 预分化、分化、生根培养基培养出苗, 炼苗后温室培养[28]。1.10 水稻栽培及表型鉴定
将野生型和转基因水稻种植于河北农业大学西校区稻竹园, 在成熟期照相并测量农艺性状。测量5株以上株高、稻穗长度、结实率和分蘖数, 计算平均值和方差。1.11 水稻种子萌发期PEG-6000拟旱处理
参照文献[29]进行PEG-6000处理: 分别取对照和转基因材料30粒种子, 用70%无水乙醇浸泡(摇床上轻轻摇晃) 5 min, 用灭菌水洗2次, 每次1 min, 用25%次氯酸钠浸泡30 min, 用灭菌水清洗3次。在装有2层滤纸的玻璃培养皿中倒入15 mL 20% PEG-6000, 以灭菌水为对照, 将消毒后的种子用镊子均匀摆放在培养皿中, 培养箱温度为30°C, 光周期L12 h/D12 h, 7 d后测量株高和芽长等, 并照相记录。1.12 水稻幼苗失水率调查
取温室培养的四叶期水稻幼苗, 剪取长度为3 cm左右的叶片, 置室温(30°C)条件下, 每0.5 h称重一次。以0时间点重量为W0, 指定时间点为Wx, 失水率(%) = (W0-Wx) × 100/Wx。试验重复3次, 计算平均值和方差[30,31]。1.13 幼苗期土培干旱处理
盆栽参试水稻材料, 种子经30°C浸泡3 d露白, 播于蛭石土(土壤与蛭石1∶1), 30°C、光周期L12 h/D12 h培养5 d, 停止浇水, 让花盆自然干燥, 经一定时间的干旱胁迫后, 植株出现萎蔫伴有死亡的迹象时, 恢复浇水, 观察并照相记录水稻状况。2 结果与分析
2.1 OsMPK17融合蛋白质的表达及抗体制备
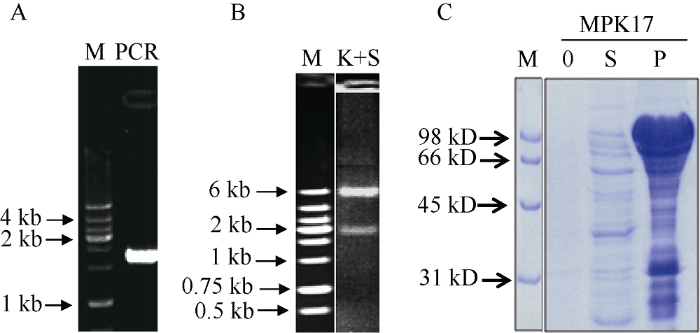
利用设计的PCR引物, 以带有全长OsMPK17 cDNA的质粒为模板进行扩增(图1-A), 扩增产物为1800 bp左右, 符合预期, 将PCR产物和pET30a表达载体进行双酶切, 连接转化细菌后提取质粒DNA进行双酶切验证(图1-B), 酶切产物为2条带, 分子量较高的是载体带, 约为6000 bp, 分子量较低的是插入片段, 约为1800 bp。将双酶切正确的质粒再进行测序验证, 挑取序列正确的重组质粒pET30a- MPK17转入表达菌Codon plus中, 挑取单菌落进行诱导表达。收集的菌体经超声破碎后离心分离, 用His-tag beads亲和层析柱纯化, 蛋白质样品经SDS-PAGE分离后, 考染观察(图1-C), 在大肠杆菌的沉淀中能清楚地看到诱导条带。利用纯化后的融合OsMPK17蛋白质作为免疫原, 免疫小鼠制备单克隆抗体。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1水稻OsMPK17基因的克隆与融合蛋白质表达
A: 水稻OsMPK17基因的PCR扩增: 以带有OsMPK17全长序列的质粒DNA为模板, 用上游引物5′-GCGGTACCATGGG CGGCCGCGCCCGCTC-3′, 下游引物5′-GCGAGCTCGGTTTTC AGTTGAGCAAC-3′, 扩增OsMPK17基因。B: pET30a-MPK17重组质粒的双酶切验证: 将PCR产物与pET30a用Kpn I+ Sac I双酶切, 连接后转化克隆菌DH5α, 提取质粒后再进行双酶切鉴定。C: 融合蛋白质OsMPK17的诱导表达及考染检测: 取pET30a-MPK17酶切验证的质粒进行测序再验证, 将测序正确的质粒转化表达菌Codon plus诱导表达。在含50 μg mL-1卡那霉素的LB液体培养基中诱导表达, 振荡培养至OD600为0.6~0.8, 加入IPTG, 25°C过夜培养, 收菌后超声破碎, 离心取上清液(S)和沉淀(P), 0: 0时间点培养物, 用10% SDS-PAGE分离、考染。M为分子量标记; PCR为扩增产物; K+S为Kpn I+ Sac I双酶切产物。
Fig. 1Cloning of rice OsMPK17 gene and fusion protein expression
A: PCR amplification of rice OsMPK17 gene. A plasmid containing full-length OsMPK17 gene was used as template for PCR amplification of OsMPK17 gene using primers 5′-GCGGTACCATGGG CGGCCGCGCCCGCTC-3′ and 5′-GCGAGCTCGGTTTTCAGTT GAGCAAC-3′. B: Verification of recombinant pET30a-MPK17 plasmid by double digestion using Kpn I and Sac I. The PCR products and pET30a plasmid DNA were digested by Kpn I and Sac I, the ligation product was used to transform DH5α. Recombinant plasmid was verified by double digestion. C: Induction of fusion protein OsMPK17 and Coomassie blue staining. Correct pET30a- MPK17 plasmid verified by double digestion was double checked by sequencing. Sequencing verified plasmid was transformed to Codon plus bacterial strain to express fusion protein. The bacteria was cultured in LB medium containing 50 μg mL-1 kanamycin and IPTG which was added when the OD600 reached 0.6-0.8. The bacteria was collected after over night culture at 25°C and disrupted by sonication. The supernatant (S) and pellet (P) were obtained after centrifugation and total protein was separated by 10% SDS-PAGE and stained with Coomassie blue. 0: Total protein isolated at 0 time point. M: Molecular weight marker; PCR: Amplification products; K+S: Double digestion product using Kpn I and Sac I.
2.2 苗期非生物逆境胁迫下OsMPK17蛋白质的表达特征
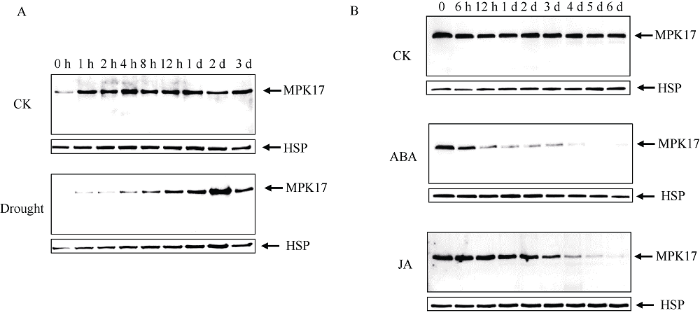
本实验室张剑硕等[23]报道了水稻RiceS-A300资源库构建的工作, 为了获得OsMPK17在逆境胁迫应答过程中可能的功能线索, 利用所制备的特异性抗体, 对这些样品进行了WB分析, 发现在干旱胁迫过程中, OsMPK17的表达量持续升高, 而对照样品中OsMPK17蛋白质的表达量维持稳定(图2-A), 提示该基因在干旱胁迫反应中可能发挥作用。对其他胁迫处理的材料进行WB分析, 没有检测到明显的表达变化(数据未附)。接下来, 以离体水稻叶片为材料, 进行ABA和MeJA处理, 提取蛋白质后进行WB分析, 发现ABA和MeJA处理均能降低OsMPK17蛋白质的丰度(图2-B), 比较而言, ABA处理的效果更为明显, 由此推测OsMPK17蛋白质的功能发挥与ABA、MeJA等激素有一定相关性。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2水稻OsMPK17蛋白质的表达特征分析
A: 干旱胁迫: 发芽后生长5 d的水稻幼苗在20% PEG-6000中水培, 分别于胁迫处理的0、1 h、2 h、4 h、8 h、12 h、1 d 、2 d和3 d取地上部样品, 提取总蛋白质后进行免疫印迹(Western blot, WB)分析。CK: 对照; Drought: PEG-6000胁迫处理。HSP: 以HSP82抗体检测的信号作为上样内参。B: 激素处理: 剪下水稻幼苗叶片在培养皿中培养, 分别添加100 μmol L-1 ABA或100 μmol L-1 MeJA进行激素处理, 取材时间点为0、6 h、12 h、1 d、2 d、3 d、4 d、5 d和6 d, 以水培为CK, 提取叶片样品总蛋白质进行WB分析。HSP: 以HSP82抗体检测的信号作为上样内参。
Fig. 2Expression profiling of OsMPK17 protein in rice by western blot analysis
A: Drought stress treatment: rice seedlings grown for 5 days were treated by 20% PEG-6000. Leaf samples were collected at 0, 1 h, 2 h, 4 h, 8 h, 12 h, 1 d, 2 d, and 3 d respectively; WB analysis were carried out for isolated total proteins. HSP: Loading control for WB analysis using HSP82 antibody. B: Treatment with hormones: leaves of rice were cultured in petri dish, 100 μmol L-1 ABA or 100 μmol L-1 MeJA was supplemented as hormone treatments. Samples were collected at 0, 6 h, 12 h, 1 d, 2 d, 3 d, 4 d, 5 d, and 6 d time points, respectively. Total proteins were isolated and analyzed by WB. HSP: Loading control for WB analysis using HSP82 antibody.
2.3 过表达OsMPK17转基因载体的构建
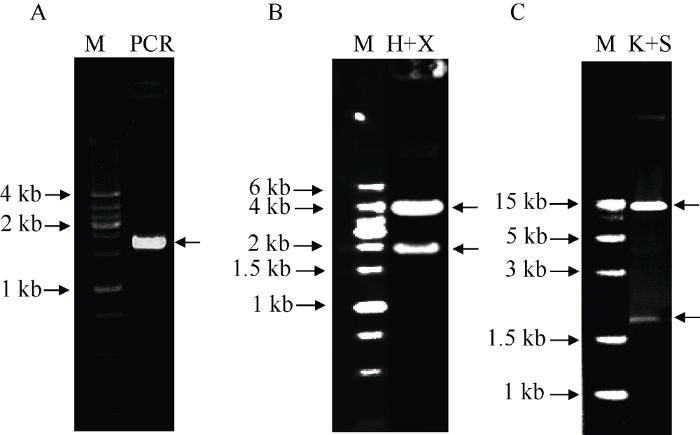
设计PCR引物, 以cDNA质粒为模板, 扩增获得了全长的OsMPK17基因(图3-A), 双酶切后装入pEASY-3HA中间载体, 测序验证后, 用内切酶Hind III和Xba I切下片段, 电泳检测到1848 bp的插入片段(图3-B), 装入水稻转化质粒pUBI-C4300中, 获得了pUBI-C4300-MPK17转化载体, 经Kpn I和Sac I酶切检测到符合预期的插入片段(图3-C), 证明获得了正确的过表达载体。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3水稻OsMPK17基因过表达载体的构建与鉴定
A: 水稻OsMPK17基因的PCR扩增; B: Hind III + Xba I双酶切鉴定pEASY-MPK17-3HA重组质粒; C: Kpn I+ Spel I双酶切鉴定pUBI-C4300-MPK17重组质粒。以带有目的基因cDNA的质粒为模板, 用上游引物5′-GCGGTACCATGGGCGGCCGCGCCC GCTC-3′ (下画线为Kpn I限制性内切酶位点)和下游引物5′-GCGAGCTCGGTTTTCAGTTGAGCAAC-3′ (下画线为Sac I限制性内切酶位点)进行PCR扩增。将扩增的OsMPK17片段插入中间载体pEASYT1-3HA, 双酶切验证。将测序正确的中间载体切胶回收目的片段, 再用Kpn I+Spel I双酶切插入转化载体pUBI-C4300, 获得重组质粒DNA双酶切验证。M: 分子量标记; PCR: 扩增产物; H+X: Hind III+ Xba I双酶切; K+S: Kpn I+ Spel I双酶切。
Fig. 3Construction and identification of rice OsMPK17 overexpression vector
A: PCR amplification of rice OsMPK17 gene; B: Hind III+ Xba I restriction enzyme digestion of recombinant pEASY-MPK17-3HA plasmid; C: Kpn I+ Spel I restriction enzyme digestion of recombinant pUBI-C4300-MPK17 plasmid. PCR amplification of OsMPK17 gene using plasmid containing full-length OsMPK17 cDNA as template, the primers used were 5′-GCGGTACCATGGGC GGCCGCGCCCGCTC-3′ (Kpn I restriction site was underlined) and 5′-GCGAGCTCGGTTTTCAGTTGAGCAAC-3′ (Sac I restriction site was underlined). The amplified fragment was inserted into pEASY-3HA vector and verified by double digestion. Sequence verified pEASY-MPK17-3HA was digested by Kpn I+ Spe I, the fragment was inserted into pUBI-C4300 and verified by double digestion. M: Molecular weight marker; PCR: PCR amplification product; H+X: Hind III+ Xba I restriction enzyme digestion; K+S: Kpn I+ Spel I restriction enzyme digestion.
2.4 水稻转化及阳性植株的鉴定
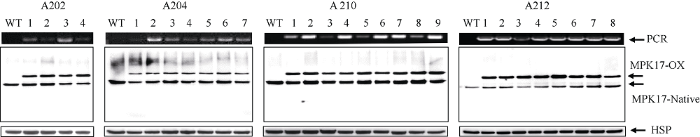
以水稻品种TP309为受体, 通过农杆菌介导的途径转化水稻, T0代获得了11个转基因株系, PCR鉴定其中10个为阳性, 收获阳性植株种子, T1代再次鉴定筛选阳性株系, 收获种子后T2代获得了纯合的过表达OsMPK17的水稻材料, 4个转基因株系(A202、A204、A210和A212)的PCR和WB检测结果如图4所示, 所检测的4个转基因株系的植株全部为PCR阳性和WB阳性, 而野生型受体水稻均表现阴性。由于过表达的OsMPK17蛋白质带有3×HA标签, 分子量比水稻中原有的OsMPK17蛋白质稍大, 所以在WB中可以检测到2个条带, 分子量稍大的是超表达的版本, 记做MPK17-OX, 分子量较小的是水稻中原来的版本, 记作MPK17-Native, 比较2个条带的信号强度可见, 二者的丰度比较接近。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4过表达OsMPK17蛋白质转基因水稻筛选与鉴定
上部: PCR结果; 中部: WB检测转基因水稻中OsMPK17蛋白质; 下部: HSP检测的信号为上样参照。WT: 野生型; A202、A204、A210和A 212为不同的转基因株系; 1、2、3、4、5、6、7、8和9为同一株系内不同的单株; PCR: PCR产物; MPK17-OX: 过表达的OsMPK17蛋白质; MPK17-Native: 水稻中原来的OsMPK17蛋白质。
Fig. 4Identification of transgenic rice plants with overexpression OsMPK17 protein
Upper panel: PCR product; Middle panel: WB detection of OsMPK17 protein in transgenic rice plants; Lower panel: HSP signal was used as loading control; WT: wildtype rice plants; A202, A204, A210, and A212 are independent transgenic lines; 1, 2, 3, 4, 5, 6, 7, 8, and 9 are independent plants among the same transgenic lines; PCR: PCR products; MPK17-OX: Over expressed OsMPK17 protein; MPK17-Native: the original form of OsMPK17 protein in rice.
2.5 过表达OsMPK17蛋白质转基因水稻植株的表型鉴定
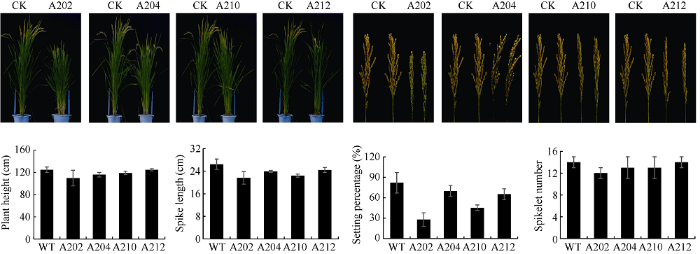
在大田栽培生长过程中调查了水稻植株的表型和主要农艺性状, 从图5可见, 过表达OsMPK17蛋白质转基因植株的株高、穗长和结实率等指标均低于野生型, 而分蘖数的差别不明显, 说明OsMPK17过表达对水稻的正常生长产生了不利的影响。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5过表达OsMPK17对水稻表型及农艺性状的影响
上部照片: 4个转基因株系(A202、A204、A210和A212)及对照水稻成熟期整株和穗部照片; 下部柱状图: 4个转基因株系及对照的株高、穗长、结实率和分蘖数的柱状图。每个指标测量5个以上单株, 计算平均值和方差。
Fig. 5Effects of over-expressed OsMPK17 protein on the phenotype and agronomic traits of rice
Photographs on the upper panel: rice whole plants and ears at mature stage of four transgenic lines and control. Bar graphs on the lower panel: plant height, spike length, seed setting rate, and tillers number of the four transgenic lines and control.
2.6 过表达OsMPK17水稻植株发芽期耐旱性鉴定
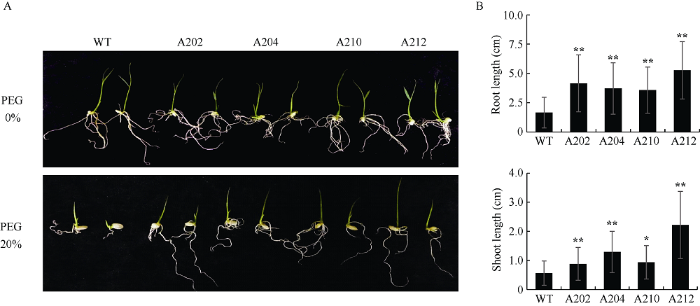
在干旱胁迫过程中, OsMPK17蛋白质的表达丰度提高, 提示该蛋白质在抵抗干旱胁迫过程中发挥正调控作用。图6-A表明4个过表达OsMPK17的株系均比野生型生长健壮, 其根长和芽长(图6-B)均明显高于野生型对照, 说明OsMPK17蛋白质的过表达提高了水稻发芽期的耐旱性。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6过表达OsMPK17蛋白质水稻发芽期耐旱性鉴定
A: 水稻种子萌发照片。上部: 对照(水); 下部: 干旱胁迫(20% PEG-6000)。B: 水稻种子萌发的根长和芽长柱状图。WT为野生型; A202、A204、A210和A212为4个转基因株系。试验重复3次, 计算平均值和方差。*表示在0.05水平差异显著; **表示在0.01水平差异显著。
Fig. 6Characterization of the germination trait of rice seeds with over-expressed OsMPK17 protein under drought stress
A: Photographs for seeds at germination. Upper panel: control (H2O); Lower panel: drought stress (20% PEG-6000) treatment. B: Bar graphs of root and shoot lengths for seeds at germination. WT: wild type; A202, A204, A210, and A212 are transgenic lines. Experiments were carried out with three replicates; average and standard derivations were calculated. * Significant at P < 0.05. ** Significant at P < 0.01.
2.7 过表达OsMPK17水稻植株幼苗期的失水率鉴定
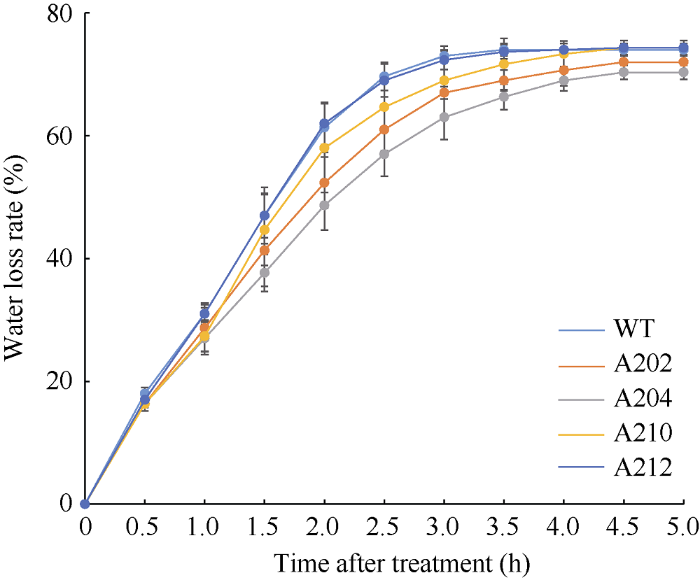
为了比较过表达OsMPK17蛋白质水稻的耐旱性, 进一步调查了水稻植株在苗期的失水率。图7表明4个过表达株系的失水率均低于或不高于野生型对照, 失水率试验支持过表达OsMPK17植株具有较好的保水性, 这可能是其耐旱性提高的原因之一。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7过表达OsMPK17蛋白质植株的失水率鉴定
WT为野生型; A202、A204、A210和A212为4个转基因株系。在水稻幼苗四叶期剪取3 cm左右叶片, 在室温下(30°C), 每30 min称重1次, 试验重复3次, 计算平均值和方差。
Fig. 7Characterization of water loss rate of OsMPK17 protein overexpressed transgenic lines
WT: wild type; A202, A204, A210, and A212 are transgenic lines. At four leaves stage, leaf blades were cut into pieces at about 3 cm, which were weighed every 30 min at room temperature (30°C). The experiment were repeated three times; the average and standard derivation were calculated.
2.8 过表达OsMPK17水稻植株苗期耐旱性鉴定
为了进一步鉴定转基因水稻的耐旱性, 将2个转基因株系(A202和A212)与野生型对照进行同盆栽培, 以便使培养条件尽量保持一致, 种子发芽后正常盆栽培养5 d, 停止浇水, 花盆土壤自然干燥, 使水稻幼苗承受实际的干旱胁迫, 经不同天数的干旱胁迫后, 待部分植株出现萎蔫并呈现叶片有50%干枯时, 恢复浇水并观察水稻苗的生长状况(图8)。转基因苗(右侧)在胁迫处理时和恢复浇水后都表现比野生型水稻苗(左侧)有更多的绿色部分, 说明转基因苗具有较强的耐旱性。图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8过表达OsMPK17蛋白质转基因水稻苗期耐旱性鉴定
水稻苗期干旱胁迫及恢复浇水试验, 照片分别为0时间点、干旱胁迫8 d和恢复浇水3 d时。WT为野生型, A202和A212为过表达OsMPK17的转基因株系。
Fig. 8Verification of drought tolerance at seedling stage of transgenic plants over-expressed OsMPK17 protein
WT: wild type; A202 and A212 were transgenic lines overexpressed OsMPK17 protein. The drought and restore experiments were carried out at seedling stage; the photographs were taken at 0 time point, eight days after drought treatment, and re-watering for 3 days.
3 讨论
蛋白质是生命活动的主要执行者, 蛋白质表达特征中蕴含着功能相关的线索。本研究制备了水稻OsMPK17蛋白质特异的抗体, 经WB分析发现该蛋白质在干旱胁迫过程中表达上调, 由此推测其在耐旱过程中发挥正调控作用, 为此构建了OsMPK17的过表达载体, 采用农杆菌介导法转化水稻, 获得了过表达OsMPK17蛋白质的转基因株系, 在萌发期对转基因材料进行PEG-6000的拟旱处理、叶片失水率调查和幼苗期的实际干旱试验, 结果均表明过表达OsMPK17转基因植株提高了水稻的耐旱性。在转基因材料的鉴定方面, 本研究除采用常规的PCR技术鉴定阳性植株外, 还通过WB对OsMPK17蛋白质的过表达进行了鉴定, 鉴定结果表明绝大部分PCR阳性植株均表现WB阳性, 但也发现有少数PCR阳性的植株表现WB阴性(数据未附), 说明PCR阳性并不能确保蛋白质的过表达, 通过WB进行鉴定是必要的。WB的检测直接证明了过表达OsMPK17蛋白质的存在。从技术上讲, PCR具有更高的灵敏度, 但也容易出现假阳性, 而WB检测的灵敏度较低, 一般不容易出现假阳性结果。有意思的是, 过表达的OsMPK17与野生型OsMPK17蛋白质的分子量有所不同, 能够被电泳区分, 所以WB可清晰地检测到野生型和过表达的OsMPK17蛋白质。根据条带信号的强度, 可大致判断出二者的丰度基本相近。用WB对转基因材料进行鉴定, 获得了转基因材料中目标蛋白质的有无、丰度及分子量等信息。
利用前期建立的RiceS-A300水稻蛋白质样品资源库[23], 比较方便地实现了OsMPK17蛋白质在各种非生物逆境胁迫下表达特征的调查, 由此发现OsMPK17蛋白质在干旱胁迫条件下表达丰度被明显诱导上调, 而在其他非生物逆境胁迫下没有发现明显的变化, 蛋白质表达特征数据提示该蛋白质可能在干旱胁迫下发挥作用。另外, 我们也检测了转基因水稻在干旱胁迫下OsMPK17的丰度变化, 未检测到原有版本OsMPK17-Native条带信号的明显增强, 这可能是由于过表达的版本OsMPK17-OX发挥作用, 抑制了OsMPK17-Native的诱导表达。通过对激素处理的离体叶片中OsMPK17蛋白质分析, 发现ABA和MeJA处理会降低OsMPK17的表达丰度。
对不同发育时期、不同部位组织中OsMPK17蛋白质的丰度分析发现, OsMPK17主要在叶片中表达, 在其他检测的部位中, 包括根、茎、叶鞘、幼穗、花药、颖壳等, 均没有检测到OsMPK17蛋白质(数据未附), 由此推测其功能应该主要通过叶片来发挥。对转录数据进行挖掘分析(http://rice.plantbiology. msu.edu/)表明, OsMPK17基因的转录主要在花药中, 在其他部位的转录信号很低(附表1)。比较蛋白质和转录信息可以看出, 二者相关性不大。此种情况下, 基于WB分析获得的蛋白质丰度信息更为直观, 也应该更具参考价值。蛋白质的表达特征既是功能表现的一种外在形式, 也是其功能的组成部分。开展重要蛋白质的表达特征调查具有重要的意义, 这也是基于抗体的靶向蛋白质组学策略试图发挥作用的方向[32]。
Supplementary table 1
附表1
附表1OsMPK17基因在不同组织中的转录丰度比较
Supplementary table 1
| 组织 Libraries | FPKM |
|---|---|
| 四叶期幼苗 Seedling four-leaf stage | 0.640 |
| 幼苗地上部 Shoots | 0.904 |
| 20 d的叶片 20-day leaves | 0.637 |
| 抽穗前花序 Pre-emergence inflorescence | 3.036 |
| 抽穗后花序 Post-emergence inflorescence | 6.281 |
| 花药 Anther | 169.643 |
| 雌蕊 Pistil | 2.371 |
| 开花后5 d种子 5 DAP seed | 5.971 |
| 开花后10 d种子 10 DAP seed | 10.372 |
| 开花后25 d幼胚 25 DAP embryo | 7.719 |
| 开花后25 d胚乳 25 DAP endosperm | 7.230 |
新窗口打开|下载CSV
水稻基因组中有17个MAPK基因, 目前, 已经鉴定到多个MAPK在生物胁迫或非生物胁迫反应中发挥功能。如通过RNAi技术降低OsMPK5表达后, 也降低了水稻对ABA的敏感性以及对干旱、盐和冷胁迫的敏感性, 但提高了对稻瘟病菌和白叶枯病菌的抗性[10,33-34]。可以认为, OsMPK5在水稻的胁迫反应中起着两方面的作用, 既可正调控ABA途径中的非生物胁迫反应, 又能负调控对稻瘟病和白叶枯病的抗病反应。此外, OsMPK7能被多种生物胁迫和非生物胁迫诱导表达[10,15,35], OsMPK17-1 (OsBWMK1)和OsMPK4 (OsMPK2)也参与植物的防御反应[6,36]。本研究中对OsMPK17的功能阐释增进了对水稻MAPK蛋白质激酶的了解。本研究通过WB检测到OsMPK17蛋白质在激素ABA和MeJA处理后, 表达丰度下降, 由此可推测OsMPK17蛋白质介导的水稻耐旱性可能与激素相关。据报道, 高水平的ABA含量会提高种子萌发期对旱的耐受性、抑制种子发芽、侧根形成、幼苗生长等, 通过促进气孔的闭合减少水分蒸腾来调控旱胁迫应答反应[37,38,39,40,41,42,43]。此外, 外源ABA的处理会影响植物中许多MAPK基因的表达, MAPK参与了ABA介导的多种信号通路, 包括氧化防御、保卫细胞信号转导和种子的萌发等[44,45,46,47]。在烟草和拟南芥中, MeJA和SA可以诱导MPK6和MPK3的表达及SIPK的瞬时表达[48,49]。这些数据提示MAPK的功能发挥与SA、MeJA及ABA等激素有密切的关系。一般认为干旱能诱导ABA含量的增加, 本文研究结果表明干旱也能诱导OsMPK17蛋白质的表达, 并增强了水稻的耐旱性。据此可推测ABA含量增加和OsMPK17的丰度增加都是干旱应答的正调控因子。但是本研究又发现, ABA处理会下调OsMPK17的丰度, 所以二者负相关。根据这些数据可以推测, 在干旱胁迫下ABA的含量会被诱导提高, 水稻应该表现耐旱性, 但ABA含量的提高能抑制OsMPK17蛋白质的表达, 如此又造成水稻的耐旱性下降, 这样两个因素在水稻中达到平衡, 其净结果表现为正常情况下水稻的耐旱性只能维持在一个特定的水平。本研究结果鉴定到OsMPK17蛋白质与干旱胁迫应答的相关性, 可能是通过激素介导响应的, 今后可在耐旱性增强机制、激素是如何发挥作用等方面开展进一步的工作。
4 结论
表达了水稻OsMPK17蛋白质, 制备了特异抗体, OsMPK17蛋白质在干旱胁迫条件下表达量升高, 在ABA和MeJA等激素处理下表达量下降。获得了过表达 OsMPK17蛋白质的转基因水稻植株, 其株高、穗长和结实率等农艺指标均低于对照。过表达OsMPK17蛋白质提高了水稻发芽期的耐旱性, 降低了幼苗期的失水率, 实际干旱恢复试验也表明过表达OsMPK17植株具有较好的耐旱性。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1126/science.1072682URLPMID:12471242 [本文引用: 1]

Multicellular organisms have three well-characterized subfamilies of mitogen-activated protein kinases (MAPKs) that control a vast array of physiological processes. These enzymes are regulated by a characteristic phosphorelay system in which a series of three protein kinases phosphorylate and activate one another. The extracellular signal-regulated kinases (ERKs) function in the control of cell division, and inhibitors of these enzymes are being explored as anticancer agents. The c-Jun amino-terminal kinases (JNKs) are critical regulators of transcription, and JNK inhibitors may be effective in control of rheumatoid arthritis. The p38 MAPKs are activated by inflammatory cytokines and environmental stresses and may contribute to diseases like asthma and autoimmunity.
DOI:10.1152/physrev.1999.79.1.143URLPMID:9922370 [本文引用: 1]
Mitogen-activated protein kinases (MAPK) are serine-threonine protein kinases that are activated by diverse stimuli ranging from cytokines, growth factors, neurotransmitters, hormones, cellular stress, and cell adherence. Mitogen-activated protein kinases are expressed in all eukaryotic cells. The basic assembly of MAPK pathways is a three-component module conserved from yeast to humans. The MAPK module includes three kinases that establish a sequential activation pathway comprising a MAPK kinase kinase (MKKK), MAPK kinase (MKK), and MAPK. Currently, there have been 14 MKKK, 7 MKK, and 12 MAPK identified in mammalian cells. The mammalian MAPK can be subdivided into five families: MAPKerk1/2, MAPKp38, MAPKjnk, MAPKerk3/4, and MAPKerk5. Each MAPK family has distinct biological functions. In Saccharomyces cerevisiae, there are five MAPK pathways involved in mating, cell wall remodelling, nutrient deprivation, and responses to stress stimuli such as osmolarity changes. Component members of the yeast pathways have conserved counterparts in mammalian cells. The number of different MKKK in MAPK modules allows for the diversity of inputs capable of activating MAPK pathways. In this review, we define all known MAPK module kinases from yeast to humans, what is known about their regulation, defined MAPK substrates, and the function of MAPK in cell physiology.
DOI:10.1023/A:1006301614690URL [本文引用: 1]
In yeast and animal cells, distinct subfamilies of mitogen-activated protein kinases (MAPKs) have evolved for transmitting different types of signals, such as the extracellular signal-regulated kinase (ERK) for mitogenic stimuli and differentiation, p38 and JUN kinase (JNK) for stress factors. Based on sequence analysis, the presently known plant MAPKs are most similar to ERKs, even though compelling evidence implies a role in various forms of biotic and abiotic stress responses. However, knowledge of their involvement in controlling proliferation is just emerging. A subgroup of the plant MAPKs, containing the alfalfa MMK3 and tobacco NTF6, are only active in mitotic cells and their localisation to the cell plate suggests a role in cytokinesis. An upstream regulator of MAPKs, the tobacco NPK1, appears to be also activated during mitosis. NPK1 might be associated and regulated by a microtubule motor protein. The localisation of NPK1 to the cell plate and its mitosis-specific activation suggest that together with NTF6 it could constitute a mitotic MAPK signalling module in tobacco. NPK1 appears to have a second role in repression of auxin-induced gene expression. MAPKs might also be involved in signalling within the meristems as suggested by the recruitement of a small G-protein to the CLAVATA 1 receptor-like protein kinase upon activation. In animal and yeast cells some of the small G-proteins relay signals from receptors to MAPK pathways.
DOI:10.1126/science.287.5454.873URLPMID:10657304 [本文引用: 1]
Genome-wide transcript profiling was used to monitor signal transduction during yeast pheromone response. Genetic manipulations allowed analysis of changes in gene expression underlying pheromone signaling, cell cycle control, and polarized morphogenesis. A two-dimensional hierarchical clustered matrix, covering 383 of the most highly regulated genes, was constructed from 46 diverse experimental conditions. Diagnostic subsets of coexpressed genes reflected signaling activity, cross talk, and overlap of multiple mitogen-activated protein kinase (MAPK) pathways. Analysis of the profiles specified by two different MAPKs-Fus3p and Kss1p-revealed functional overlap of the filamentous growth and mating responses. Global transcript analysis reflects biological responses associated with the activation and perturbation of signal transduction pathways.
DOI:10.1146/annurev-arplant-042809-112252URLPMID:20441529 [本文引用: 1]
Eukaryotic mitogen-activated protein kinase (MAPK) cascades have evolved to transduce environmental and developmental signals into adaptive and programmed responses. MAPK cascades relay and amplify signals via three types of reversibly phosphorylated kinases leading to the phosphorylation of substrate proteins, whose altered activities mediate a wide array of responses, including changes in gene expression. Cascades may share kinase components, but their signaling specificity is maintained by spaciotemporal constraints and dynamic protein-protein interactions and by mechanisms that include crossinhibition, feedback control, and scaffolding. Plant MAPK cascades regulate numerous processes, including stress and hormonal responses, innate immunity, and developmental programs. Genetic analyses have uncovered several predominant MAPK components shared by several of these processes including the Arabidopsis thaliana MAPKs MPK3, 4, and 6 and MAP2Ks MKK1, 2, 4, and 5. Future work needs to focus on identifying substrates of MAPKs, and on understanding how specificity is achieved among MAPK signaling pathways.
DOI:10.1094/MPMI.1999.12.12.1064URLPMID:10624015 [本文引用: 2]
The activation of the mitogen-activated protein (MAP) kinases by different environmental stresses has been previously observed in several dicot plant species. Here, we report the isolation of a novel MAP kinase in rice that is induced during infection by the blast fungus Magnaporthe grisea or upon mechanical wounding. The gene is designated as BWMK1 for blast- and wound-induced MAP kinase. The cDNA of BWMK1 was isolated from rice leaves challenged by the blast pathogen. Transcripts of the corresponding gene accumulated in rice leaves 4 h after blast inoculation and 30 min after mechanical wounding. This gene encodes a 506 amino acid protein that contains a new dual-phosphorylation activation motif TDY and about 150 unique amino acids on its C terminus. In-gel kinase activity and immunoprecipitation assays confirmed that BWMK1 is a functional MAP kinase. These results show that BWMK1 is a new member of the plant MAP kinase family and may mediate both defense and wound signaling in rice.
DOI:10.1016/s0006-291x(02)02868-1URLPMID:12507518 [本文引用: 1]
We report isolation of two novel rice (Oryza sativa L.) mitogen-activated protein kinases (MAPKs), OsMSRMK3 (multiple stress responsive) and OsWJUMK1 (wound- and JA-uninducible) that most likely exist as single copy genes in its genome. OsMSRMK3 and OsWJUMK1 encode 369 and 569 amino acid polypeptides having the MAPK family signature and phosphorylation activation motifs TEY and TDY, respectively. Steady state mRNA analyses of these MAPKs with constitutive expression in leaves of two-week-old seedlings revealed that OsMSRMK3 was up-regulated upon wounding (by cut), jasmonic acid (JA), salicylic acid (SA), ethylene, abscisic acid, hydrogen peroxide (H(2)O(2)), protein phosphatase inhibitors, chitosan, high salt/sugar, and heavy metals, whereas OsWJUMK1 not induced by either wounding, JA or SA, showed up-regulation only by H(2)O(2), heavy metals, and cold stress (12 degrees C). Moreover, these MAPKs were developmentally regulated. These results strongly suggest a role for OsMSRMK3 and OsWJUMK1 in both stress-signalling pathways and development in rice.
DOI:10.1111/pce.12154URL [本文引用: 1]
In rice, the Ca2+/calmodulin (CaM)-dependent protein kinase (CCaMK) OsDMI3 has been shown to be required for abscisic acid (ABA)-induced antioxidant defence. However, it is not clear how OsDMI3 participates in this process in rice. In this study, the cross-talk between OsDMI3 and the major ABA-activated MAPK OsMPK1 in ABA-induced antioxidant defence was investigated. ABA treatment induced the expression of OsDMI3 and OsMPK1 and the activities of OsDMI3 and OsMPK1 in rice leaves. In the mutant of OsDMI3, the ABA-induced increases in the expression and the activity of OsMPK1 were substantially reduced. But in the mutant of OsMPK1, the ABA-induced increases in the expression and the activity of OsDMI3 were not affected. Pretreatments with MAPKK inhibitors also did not affect the ABA-induced activation of OsDMI3. Further, a transient expression analysis in combination with mutant analysis in rice protoplasts showed that OsMPK1 is required for OsDMI3-induced increases in the activities of antioxidant enzymes and the production of H2O2. Our data indicate that there exists a cross-talk between OsDMI3 and OsMPK1 in ABA signalling, in which OsDMI3 functions upstream of OsMPK1 to regulate the activities of antioxidant enzymes and the production of H2O2 in rice.
DOI:10.1042/BJ20111792URLPMID:22248149 [本文引用: 1]
MAPK (mitogen-activated protein kinase) pathways have been implicated in stress signalling in plants. In the present study, we performed yeast two-hybrid screening to identify partner MAPKs for OsMKK (Oryza sativa MAPK kinase) 6, a rice MAPK kinase, and revealed specific interactions of OsMKK6 with OsMPK3 and OsMPK6. OsMPK3 and OsMPK6 each co-immunoprecipitated OsMKK6, and both were directly phosphorylated by OsMKK6 in vitro. An MBP (myelin basic protein) kinase assay of the immunoprecipitation complex indicated that OsMPK3 and OsMPK6 were activated in response to a moderately low temperature (12°C), but not a severely low temperature (4°C) in rice seedlings. A constitutively active form of OsMKK6, OsMKK6DD, showed elevated phosphorylation activity against OsMPK3 and OsMPK6 in vitro. OsMPK3, but not OsMPK6, was constitutively activated in transgenic plants overexpressing OsMKK6DD, indicating that OsMPK3 is an in vivo target of OsMKK6. Enhanced chilling tolerance was observed in the transgenic plants overexpressing OsMKK6DD. Taken together, our data suggest that OsMKK6 and OsMPK3 constitute a moderately low-temperature signalling pathway and regulate cold stress tolerance in rice.
DOI:10.1105/tpc.008714URLPMID:12615946 [本文引用: 3]
Mitogen-activated protein kinase (MAPK) cascades play an important role in mediating stress responses in eukaryotic organisms. However, little is known about the role of MAPKs in modulating the interaction of defense pathways activated by biotic and abiotic factors. In this study, we have isolated and functionally characterized a stress-responsive MAPK gene (OsMAPK5) from rice. OsMAPK5 is a single-copy gene but can generate at least two differentially spliced transcripts. The OsMAPK5 gene, its protein, and kinase activity were inducible by abscisic acid as well as various biotic (pathogen infection) and abiotic (wounding, drought, salt, and cold) stresses. To determine its biological function, we generated and analyzed transgenic rice plants with overexpression (using the 35S promoter of Cauliflower mosaic virus) or suppression (using double-stranded RNA interference [dsRNAi]) of OsMAPK5. Interestingly, suppression of OsMAPK5 expression and its kinase activity resulted in the constitutive expression of pathogenesis-related (PR) genes such as PR1 and PR10 in the dsRNAi transgenic plants and significantly enhanced resistance to fungal (Magnaporthe grisea) and bacterial (Burkholderia glumae) pathogens. However, these same dsRNAi lines had significant reductions in drought, salt, and cold tolerance. By contrast, overexpression lines exhibited increased OsMAPK5 kinase activity and increased tolerance to drought, salt, and cold stresses. These results strongly suggest that OsMAPK5 can positively regulate drought, salt, and cold tolerance and negatively modulate PR gene expression and broad-spectrum disease resistance.
DOI:10.1016/j.devcel.2017.11.016URLPMID:29257952 [本文引用: 1]
Improvement of chilling tolerance is a major target in rice breeding. The signaling pathways regulating chilling consist of complex networks, including key transcription factors and their targets. However, it remains largely unknown how transcription factors are activated by chilling stress. Here, we report that?the transcription factor OsbHLH002/OsICE1 is phosphorylated by OsMAPK3 under chilling stress. The osbhlh002-1 knockout mutant and antisense transgenic plants showed chilling hypersensitivity, whereas OsbHLH002-overexpressing plants exhibited enhanced chilling tolerance. OsbHLH002 can directly target OsTPP1, which encodes a key enzyme for trehalose biosynthesis. OsMAPK3 interacts with OsbHLH002 to prevent its ubiquitination by the E3 ligase OsHOS1. Under chilling stress, active OsMAPK3 phosphorylates OsbHLH002, leading to accumulation of phospho-OsbHLH002, which promotes OsTPP1 expression and increases trehalose content and resistance to chilling damage.?Taken together, these results indicate that OsbHLH002 is phosphorylated by OsMAPK3, which enhances OsbHLH002 activation to its target OsTPP1 during chilling stress.
DOI:10.1016/j.plantsci.2014.08.007URL [本文引用: 1]
Mitogen-activated protein kinase (MAPK) pathways have been implicated in signal transduction of both biotic and abiotic stresses in plants. In this study, we found that the transcript of a rice (Oryza sativa) MAPKK (OsMKK1) was markedly increased by salt stress. By examining the survival rate and Na+ content in shoot, we found that OsMKK1-knockout (osmkk1) mutant was more sensitive to salt stress than the wild type. OsMKK1 activity in the wild-type seedlings and protoplasts was increased by salt stress. Yeast two-hybrid and in vitro and in vivo kinase assays revealed that OsMKK1 targeted OsMPK4. OsMPK4 activity was increased by salt, which was impaired in osmkk1 plants. In contrast, overexpression of OsMKK1 increased OsMPK4 activity in protoplasts. By comparing the transcription factors levels between WT and osmkk1 mutant, OsMKK1 was necessary for salt-induced increase in OsDREB2B and OsMYBS3. Taken together, the data suggest that OsMKK1 and OsMPK4 constitute a signaling pathway that regulates salt stress tolerance in rice. (C) 2014 Elsevier Ireland Ltd.
DOI:10.1104/pp.111.174334URL [本文引用: 1]
We examined ways in which the Brown planthopper induced008a (Bphi008a; AY256682) gene of rice (Oryza sativa) enhances the plant's resistance to a specialist herbivore, the brown planthopper (BPH; Nilaparvata lugens). Measurement of the expression levels of ethylene synthases and of ethylene emissions showed that BPH feeding rapidly initiated the ethylene signaling pathway and up-regulated Bphi008a transcript levels after 6 to 96 h of feeding. In contrast, blocking ethylene transduction (using 1-methylcyclopropene) reduced Bphi008a transcript levels in wild-type plants fed upon by BPH. In vitro kinase assays showed that Bphi008a can be phosphorylated by rice Mitogen-activated Protein Kinase5 (OsMPK5), and yeast two-hybrid assays demonstrated that the carboxyl-terminal proline-rich region of Bphi008a interacts directly with this kinase. Furthermore, bimolecular fluorescence complementation assays showed that this interaction occurs in the nucleus. Subsequently, we found that Bphi008a up-regulation and down-regulation were accompanied by different changes in transcription levels of OsMPK5, OsMPK12, OsMPK13, and OsMPK17 in transgenic plants. Immunoblot analysis also showed that the OsMPK5 protein level increased in overexpressing plants and decreased in RNA interference plants after BPH feeding. In transgenic lines, changes in the expression levels of several enzymes that are important components of the defenses against the BPH were also observed. Finally, yeast two-hybrid screening results showed that Bphi008a is able to interact with a b-ZIP transcription factor (OsbZIP60) and a RNA polymerase polypeptide (SDRP).
DOI:10.1016/j.molp.2018.04.004URLPMID:29702261 [本文引用: 1]
Grain size is one of the key agronomic traits that determine grain yield in crops. However, the mechanisms underlying grain size control in crops remain elusive. Here we demonstrate that the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway positively regulates grain size and weight in rice. In rice, loss of OsMKKK10 function results in small and light grains, short panicles, and semi-dwarf plants, while overexpression of constitutively active OsMKKK10 (CA-OsMKKK10) results in large and heavy grains, long panicles, and tall plants. OsMKKK10 interacts with and phosphorylates OsMKK4. We identified an OsMKK4 gain-of-function mutant (large11-1D)?that produces large and heavy grains. OsMKK4A227T encoded by the large11-1D allele has stronger kinase activity than OsMKK4. Plants overexpressing a constitutively active form of OsMKK4 (OsMKK4-DD) also produce large grains. Further biochemical and genetic analyses revealed that OsMKKK10, OsMKK4, and OsMAPK6 function in a common pathway to control grain size. Taken together, our study establishes an important genetic and molecular framework for OsMKKK10-OsMKK4-OsMAPK6 cascade-mediated control of grain size and weight in rice.
DOI:10.1104/pp.006072URLPMID:12177502 [本文引用: 2]
Rice (Oryza sativa) anther development is easily damaged by moderately low temperatures above 12 degrees C. Subtractive screening of cDNA that accumulated in 12 degrees C-treated anthers identified a cDNA clone, OsMEK1, encoding a protein with features characteristic of a mitogen-activated protein (MAP) kinase kinase. The putative OsMEK1 protein shows 92% identity to the maize (Zea mays) MEK homolog, ZmMEK1. OsMEK1 transcript levels were induced in rice anthers by 12 degrees C treatment for 48 h. Similar OsMEK1 induction was observed in shoots and roots of seedlings that were treated at 12 degrees C for up to 24 h. It is interesting that no induction of OsMEK1 transcripts was observed in 4 degrees C-treated seedlings. In contrast, rice lip19, encoding a bZIP protein possibly involved in low temperature signal transduction, was not induced by 12 degrees C treatment but was induced by 4 degrees C treatment. Among the three MAP kinase homologs cloned, only OsMAP1 displayed similar 12 degrees C-specific induction pattern as OsMEK1. A yeast two-hybrid system revealed that OsMEK1 interacts with OsMAP1, but not with OsMAP2 and OsMAP3, suggesting that OsMEK1 and OsMAP1 probably function in the same signaling pathway. An in-gel assay of protein kinase activity revealed that a protein kinase (approximately 43 kD), which preferentially uses myelin basic protein as a substrate, was activated by 12 degrees C treatment but not by 4 degrees C treatment. Taken together, these results lead us to conclude that at least two signaling pathways for low temperature stress exist in rice, and that a MAP kinase pathway with OsMEK1 and OsMAP1 components is possibly involved in the signaling for the higher range low-temperature stress.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s12038-011-9002-8URLPMID:21451255 [本文引用: 1]
Mitogen-activated protein kinases (MAPK) signalling cascades are activated by extracellular stimuli such as environmental stresses and pathogens in higher eukaryotic plants. To know more about MAPK signalling in plants, aMAPK cDNA clone, OsMAPK33, was isolated from rice. The gene is mainly induced by drought stress. In phylogenetic analysis, OsMAPK33 (Os02g0148100) showed approximately 47-93% identity at the amino acid level with other plant MAPKs. It was found to exhibit organ-specific expression with relatively higher expression in leaves as compared with roots or stems, and to exist as a single copy in the rice genome. To investigate the biological functions of OsMAPK33 in rice MAPK signalling, transgenic rice plants that either overexpressed or suppressed OsMAPK33 were made. Under dehydration conditions, the suppressed lines showed lower osmotic potential compared with that of wild-type plants, suggesting a role of OsMAPK33 in osmotic homeostasis. Nonetheless, the suppressed lines did not display any significant difference in drought tolerance compared with their wild-type plants. With increased salinity, there was still no difference in salt tolerance between OsMAPK33-suppressed lines and their wild-type plants. However, the overexpressing lines showed greater reduction in biomass accumulation and higher sodium uptake into cells, resulting in a lower K+/Na+ ratio inside the cell than that in the wild-type plants and OsMAPK33-suppressed lines. These results suggest that OsMAPK33 could play a negative role in salt tolerance through unfavourable ion homeostasis. Gene expression profiling of OsMAPK33 transgenic lines through rice DNA chip analysis showed that OsMAPK33 altered expression of genes involved in ion transport. Further characterization of downstream components will elucidate various biological functions of this novel rice MAPK.
DOI:10.1074/jbc.M110999200URLPMID:11927577 [本文引用: 1]
The rice disease resistance gene, Xa21, encodes a receptor kinase-like protein consisting of leucine-rich repeats in the putative extracellular domain and a serine/threonine kinase in the putative intracellular domain. The putative XA21 kinase domain was expressed as maltose-binding and glutathione S-transferase fusion proteins in Escherichia coli. The fusion proteins are capable of autophosphorylation. Phosphoamino acid analysis of the glutathione S-transferase fusion protein indicates that only serine and threonine residues are phosphorylated. The relative phosphorylation rate of the XA21 kinase against increasing enzyme concentrations follows a first-order rather than second-order kinetics, indicating an intramolecular phosphorylation mechanism. Moreover, the active XA21 kinase cannot phosphorylate a kinase-deficient mutant of XA21 kinase. The enzymatic activity of the XA21 kinase in a buffer containing Mn(2+) is at least 15 times higher than that with Mg(2+). The K(m) and V(max) of XA21 kinase for ATP are 0.3 microm and 8.4 nmol/mg/min, respectively. Tryptic phosphopeptide mapping reveals that multiple sites on the XA21 kinase are phosphorylated. Finally, our data suggest that the region of XA21 kinase corresponding to the RD kinase activation domain is not phosphorylated, revealing a distinct mode of action compared with the tomato Pto serine/threonine kinase conferring disease resistance.
DOI:10.1007/s12033-010-9276-3URL [本文引用: 1]
A number of primer design programs have been developed for diverse applications. However, none of these programs can be used to design primers for gene cloning aimed at expressing protein. Here we report the design of PrimerCE, which can be used to cover the whole process of gene cloning and expression. The main features of PrimerCE include inspection of restriction enzyme recognition sequence, open reading frame verification, stop codon inspection, base adjustment, primer optimization, sequence assembly and protein analysis. In addition to this, the program can be modified based on the different needs of users, e.g. new vector sequence and restriction enzyme recognition sequences can be integrated. With the use of PrimerCE, a pair of primers can be designed within minutes. The program has been proven to be efficient in designing primers in our high throughput cloning and gene expression experiments. The software is freely available at http://tch.hebau.edu.cn/shengm/download/down.html.
DOI:10.3864/j.issn.0578-1752.2017.19.001URL [本文引用: 1]
【目的】制备抗CAS9蛋白质的单克隆抗体,建立转基因水稻中CAS9蛋白质的免疫印迹检测方法,了解CAS9蛋白质在转基因水稻中的表达特征。【方法】以带Cas9的质粒DNA为模板,PCR扩增Cas9 5′端810 bp片段后克隆到pET30a载体中,将酶切验证且序列正确的重组质粒转入表达菌Condon Plus中,经IPTG诱导表达CAS9蛋白质(N端270氨基酸),用纯化的重组蛋白质作为免疫原免疫小鼠,制备单克隆抗体,用免疫印迹分析筛选特异性和灵敏度高的抗体细胞株。用特异性引物PCR扩增转基因水稻的Cas9,用免疫印迹检测转基因水稻中的CAS9蛋白质。将重组CAS9蛋白质和带有CAS9的水稻苗期叶片蛋白质进行平行免疫印迹分析,用Image J软件采集信号绘制CAS9蛋白质的标准曲线,进而对水稻叶片中的CAS9蛋白质进行定量分析。提取单粒稻米的总蛋白质,稀释后用免疫印迹分析CAS9蛋白质的检测下限,提取多个时期、部位的水稻材料的总蛋白质,包括苗期的地上部和地下部,分蘖期的茎、茎节、叶鞘、叶片等,用SDS-PAGE分离后免疫印迹检测比较CAS9蛋白质的表达特征。【结果】通过体外克隆、诱导表达获得了纯化的重组CAS9蛋白质N端片段,免疫小鼠后得到42株阳性杂交瘤细胞株,经筛选鉴定到#12D2细胞株对水稻样品的检测具有较好的特异性和灵敏度。用该抗体通过免疫印迹检测了转基因水稻,所建立的免疫印迹方法对重组CAS9蛋白质的检测下限约为0.25 ng。在水稻苗期叶片中,CAS9蛋白质约占鲜重的0.00005%,可检出单粒稻米8%样品(约2 mg)中的CAS9蛋白质。在苗期地上部CAS9蛋白质的表达丰度高于地下部,分蘖期茎和叶片中表达量较高,根和叶鞘表达量较低。【结论】获得特异性强、灵敏度高的抗CAS9单克隆抗体,建立了检测转基因水稻中CAS9蛋白质的免疫印记方法,揭示了CAS9蛋白质在水稻不同部位的表达特征,并展示了在其他植物中的应用潜力。
DOI:10.3864/j.issn.0578-1752.2017.19.001URL [本文引用: 1]
【目的】制备抗CAS9蛋白质的单克隆抗体,建立转基因水稻中CAS9蛋白质的免疫印迹检测方法,了解CAS9蛋白质在转基因水稻中的表达特征。【方法】以带Cas9的质粒DNA为模板,PCR扩增Cas9 5′端810 bp片段后克隆到pET30a载体中,将酶切验证且序列正确的重组质粒转入表达菌Condon Plus中,经IPTG诱导表达CAS9蛋白质(N端270氨基酸),用纯化的重组蛋白质作为免疫原免疫小鼠,制备单克隆抗体,用免疫印迹分析筛选特异性和灵敏度高的抗体细胞株。用特异性引物PCR扩增转基因水稻的Cas9,用免疫印迹检测转基因水稻中的CAS9蛋白质。将重组CAS9蛋白质和带有CAS9的水稻苗期叶片蛋白质进行平行免疫印迹分析,用Image J软件采集信号绘制CAS9蛋白质的标准曲线,进而对水稻叶片中的CAS9蛋白质进行定量分析。提取单粒稻米的总蛋白质,稀释后用免疫印迹分析CAS9蛋白质的检测下限,提取多个时期、部位的水稻材料的总蛋白质,包括苗期的地上部和地下部,分蘖期的茎、茎节、叶鞘、叶片等,用SDS-PAGE分离后免疫印迹检测比较CAS9蛋白质的表达特征。【结果】通过体外克隆、诱导表达获得了纯化的重组CAS9蛋白质N端片段,免疫小鼠后得到42株阳性杂交瘤细胞株,经筛选鉴定到#12D2细胞株对水稻样品的检测具有较好的特异性和灵敏度。用该抗体通过免疫印迹检测了转基因水稻,所建立的免疫印迹方法对重组CAS9蛋白质的检测下限约为0.25 ng。在水稻苗期叶片中,CAS9蛋白质约占鲜重的0.00005%,可检出单粒稻米8%样品(约2 mg)中的CAS9蛋白质。在苗期地上部CAS9蛋白质的表达丰度高于地下部,分蘖期茎和叶片中表达量较高,根和叶鞘表达量较低。【结论】获得特异性强、灵敏度高的抗CAS9单克隆抗体,建立了检测转基因水稻中CAS9蛋白质的免疫印记方法,揭示了CAS9蛋白质在水稻不同部位的表达特征,并展示了在其他植物中的应用潜力。
DOI:10.1093/jxb/err084URL [本文引用: 1]
Studies of rice protein expression have increased considerably with the development of rice functional genomics. In order to obtain reliable expression results in western blotting, information on appropriate reference proteins is necessary for data normalization. To date, no published study has identified and systematically validated reference proteins suitable for the investigation of rice protein expression. In this study, nine candidate proteins were selected and their specific antibodies were obtained through immunization of rabbits with either recombinant proteins expressed in Escherichia coli or synthesized peptides. Western blotting was carried out to detect the expression of target proteins in a set of 10 rice samples representing different rice tissues/organs at different developmental stages. The expression stability of the proteins was analysed using geNorm and Microcal Origin 6.0 software. The results indicated that heat shock protein (HSP) and elongation factor 1-alpha (eEF-1 alpha) were the most constantly expressed among all rice proteins tested throughout all developmental stages, while the proteins encoded by conventional internal reference genes fluctuated in amount. Comparison among the profiling of translation and transcription [expressed sequence tags (EST) and massively parallel signature sequencing (MPSS)] revealed that a correlation existed. Based on the standard curves derived from the antigen-antibody reaction, the concentrations of HSP and eEF-1 alpha proteins in rice leaves were similar to 0.12%. Under the present experimental conditions, the lower limits of detection for HSP and eEF-1 alpha proteins in rice were 0.24 ng and 0.06 ng, respectively. In conclusion, the reference proteins selected in this study, and the corresponding antibodies, can be used in qualitative and quantitative analysis of rice proteins.
DOI:10.3864/j.issn.0578-1752.2014.14.002URL [本文引用: 1]
【目的】建立转基因水稻中GUS蛋白质的免疫学检测方法,并了解花椰菜花叶病毒(CaMV)35S启动子驱动的GUS蛋白质在转基因水稻中的表达特征。【方法】以细菌基因组DNA为模板,PCR扩增GUS基因后克隆到表达载体pET30a中,测序验证的重组子转入大肠杆菌表达菌BL21中,IPTG诱导获得重组表达的GUS蛋白质,用HIS-tag beads纯化后作为免疫原免疫小鼠制备GUS蛋白质特异的抗体,通过免疫印迹分析筛选高特异性的单克隆抗体,用Broadford法对重组的GUS蛋白质进行定量,对不同浓度的GUS蛋白质进行免疫印迹分析,绘制检测GUS蛋白质的标准曲线,通过与标准曲线的比较对水稻叶片中GUS蛋白质进行定量分析。提取不同时期、不同部位的水稻总蛋白质,包括苗期的地上部、地下部,分蘖期的茎、茎节、叶鞘、叶枕、叶片上部、叶片中部和叶片下部,孕穗期的茎、穗轴、叶鞘、叶枕、叶片、幼穗(长度分别为1、2、10和20 cm),开花期的茎、穗轴、叶鞘、叶片、穗子,成熟期的茎、叶片、授粉后不同时期的种子(分别为授粉后10、20、30和40 d)、乳熟期的胚、胚乳和颖壳、成熟种子的全种子、胚、胚乳和颖壳以及不同时期的叶片和根部材料等。SDS-PAGE分离后用抗体检测其GUS蛋白质的丰度。【结果】筛选获得了高特异性的抗GUS单克隆抗体(编号为#27),用该抗体检测转基因水稻中及重组的GUS蛋白质均呈现特异条带,没有可见的背景信号,用本研究建立的免疫印迹方法对重组GUS蛋白质的检测下限约为4 ng,可检出转基因水稻单粒大米2.5%样品中(约0.6 mg)的GUS蛋白质。在不同时期的转基因水稻叶片中GUS蛋白质的表达丰度基本稳定,而在水稻根部的GUS丰度随生长急剧减少,5叶期根中的表达量不到3叶期的三分之一,到6叶期检测不到GUS蛋白质。在水稻苗期叶片中,GUS蛋白质约占鲜重的0.02‰。另外,除分蘖期以后的根部之外,GUS蛋白质几乎在所有的水稻组织部位中呈组成型表达,只是不同组织中的表达量略有差异,如在孕穗期和开花期的茎及颖壳中的表达量较低。【结论】建立了具有应用价值的对转基因水稻中GUS蛋白质丰度检测的免疫印记方法。该方法特异性高、样品用量少、不依赖于GUS蛋白质的酶活性、测定结果易于在不同实验室间比较。证明了35S启动子驱动的GUS蛋白质在转基因水稻中基本呈组成型表达。
DOI:10.3864/j.issn.0578-1752.2014.14.002URL [本文引用: 1]
【目的】建立转基因水稻中GUS蛋白质的免疫学检测方法,并了解花椰菜花叶病毒(CaMV)35S启动子驱动的GUS蛋白质在转基因水稻中的表达特征。【方法】以细菌基因组DNA为模板,PCR扩增GUS基因后克隆到表达载体pET30a中,测序验证的重组子转入大肠杆菌表达菌BL21中,IPTG诱导获得重组表达的GUS蛋白质,用HIS-tag beads纯化后作为免疫原免疫小鼠制备GUS蛋白质特异的抗体,通过免疫印迹分析筛选高特异性的单克隆抗体,用Broadford法对重组的GUS蛋白质进行定量,对不同浓度的GUS蛋白质进行免疫印迹分析,绘制检测GUS蛋白质的标准曲线,通过与标准曲线的比较对水稻叶片中GUS蛋白质进行定量分析。提取不同时期、不同部位的水稻总蛋白质,包括苗期的地上部、地下部,分蘖期的茎、茎节、叶鞘、叶枕、叶片上部、叶片中部和叶片下部,孕穗期的茎、穗轴、叶鞘、叶枕、叶片、幼穗(长度分别为1、2、10和20 cm),开花期的茎、穗轴、叶鞘、叶片、穗子,成熟期的茎、叶片、授粉后不同时期的种子(分别为授粉后10、20、30和40 d)、乳熟期的胚、胚乳和颖壳、成熟种子的全种子、胚、胚乳和颖壳以及不同时期的叶片和根部材料等。SDS-PAGE分离后用抗体检测其GUS蛋白质的丰度。【结果】筛选获得了高特异性的抗GUS单克隆抗体(编号为#27),用该抗体检测转基因水稻中及重组的GUS蛋白质均呈现特异条带,没有可见的背景信号,用本研究建立的免疫印迹方法对重组GUS蛋白质的检测下限约为4 ng,可检出转基因水稻单粒大米2.5%样品中(约0.6 mg)的GUS蛋白质。在不同时期的转基因水稻叶片中GUS蛋白质的表达丰度基本稳定,而在水稻根部的GUS丰度随生长急剧减少,5叶期根中的表达量不到3叶期的三分之一,到6叶期检测不到GUS蛋白质。在水稻苗期叶片中,GUS蛋白质约占鲜重的0.02‰。另外,除分蘖期以后的根部之外,GUS蛋白质几乎在所有的水稻组织部位中呈组成型表达,只是不同组织中的表达量略有差异,如在孕穗期和开花期的茎及颖壳中的表达量较低。【结论】建立了具有应用价值的对转基因水稻中GUS蛋白质丰度检测的免疫印记方法。该方法特异性高、样品用量少、不依赖于GUS蛋白质的酶活性、测定结果易于在不同实验室间比较。证明了35S启动子驱动的GUS蛋白质在转基因水稻中基本呈组成型表达。
DOI:10.3864/j.issn.0578-1752.2018.19.001URL [本文引用: 3]
【目的】建立水稻幼苗胁迫处理的蛋白质样品资源库RiceS-A300,应用RiceS-A300调查内参蛋白质HSP82的表达特征。【方法】粳稻TP309种子在30℃条件下浸泡3 d露白,土培用蛭石土(土壤与蛭石1﹕1)培养,30℃、光周期L12 h/D12 h培养5 d,进行冷(4℃)、热(44℃、48℃)、淹和恒光、恒暗处理,水培播种在纱网上,以30℃、光周期L12 h/D12 h培养5 d,进行PEG 6000(20%)和NaCl(0.2 mol·L-1)处理。以30℃、光周期L12 h/D12 h非胁迫处理为对照。在不同时间点观察、拍照并取材,测定各种处理条件下的株高和鲜重,提取总蛋白质建立水稻幼苗胁迫处理的蛋白质样品资源库,SDS-PAGE分离总蛋白质后进行考染,以此评价蛋白质质量。通过免疫印迹(WB)技术分析样品中内参蛋白质HSP82的表达特征。【结果】调查各种胁迫处理过程中9个时间点的幼苗表型、株高、鲜重和总蛋白质含量,发现:冷(4℃)、热(44℃、48℃)温度胁迫在2 d内即表现出对株高的抑制作用,淹胁迫能促进株高伸长,恒暗和恒光处理均能抑制株高,恒暗处理使幼苗变黄,PEG与盐胁迫也能明显抑制株高的伸长,且盐胁迫在5 d后可见明显叶尖干枯。对鲜重的调查结果表明,冷(4℃)、热(44℃、48℃)温度胁迫、PEG胁迫和盐胁迫均会降低幼苗鲜重,淹胁迫对鲜重的影响较小,恒暗处理会降低鲜重而恒光处理对鲜重影响不明显。3种温度胁迫都使总蛋白质含量提高,恒暗胁迫则降低总蛋白质含量,其他胁迫对总蛋白质含量影响不大。累计收集近300份蛋白质样品,建立了水稻蛋白质样品资源库(RiceS-A300),每份样品体积为2 mL,可供200次WB分析,由于试验所采用的条件比较规范且容易控制,可根据需要重复扩大样品的规模,预期不同的实验室采用相同的条件获得的蛋白质样品也能获得可重复的试验结果,利用RiceS-A300调查HSP82蛋白质的表达特征,显示热胁迫明显提高了HSP82的表达,恒光、PEG和盐胁迫也对HSP82的表达有一定影响,而淹胁迫和恒暗处理对HSP82的影响不大。对RiceS-A300资源库的评价扩展了HSP82内参蛋白质的用途。【结论】提出了蛋白质样品资源库建设和评价的流程,建立了第一个版本的胁迫处理条件下的水稻幼苗蛋白质样品资源库(RiceS-A300);在此基础上,通过WB调查了HSP82蛋白质的表达特征,发现其在热胁迫下有特异诱导表达。
DOI:10.3864/j.issn.0578-1752.2018.19.001URL [本文引用: 3]
【目的】建立水稻幼苗胁迫处理的蛋白质样品资源库RiceS-A300,应用RiceS-A300调查内参蛋白质HSP82的表达特征。【方法】粳稻TP309种子在30℃条件下浸泡3 d露白,土培用蛭石土(土壤与蛭石1﹕1)培养,30℃、光周期L12 h/D12 h培养5 d,进行冷(4℃)、热(44℃、48℃)、淹和恒光、恒暗处理,水培播种在纱网上,以30℃、光周期L12 h/D12 h培养5 d,进行PEG 6000(20%)和NaCl(0.2 mol·L-1)处理。以30℃、光周期L12 h/D12 h非胁迫处理为对照。在不同时间点观察、拍照并取材,测定各种处理条件下的株高和鲜重,提取总蛋白质建立水稻幼苗胁迫处理的蛋白质样品资源库,SDS-PAGE分离总蛋白质后进行考染,以此评价蛋白质质量。通过免疫印迹(WB)技术分析样品中内参蛋白质HSP82的表达特征。【结果】调查各种胁迫处理过程中9个时间点的幼苗表型、株高、鲜重和总蛋白质含量,发现:冷(4℃)、热(44℃、48℃)温度胁迫在2 d内即表现出对株高的抑制作用,淹胁迫能促进株高伸长,恒暗和恒光处理均能抑制株高,恒暗处理使幼苗变黄,PEG与盐胁迫也能明显抑制株高的伸长,且盐胁迫在5 d后可见明显叶尖干枯。对鲜重的调查结果表明,冷(4℃)、热(44℃、48℃)温度胁迫、PEG胁迫和盐胁迫均会降低幼苗鲜重,淹胁迫对鲜重的影响较小,恒暗处理会降低鲜重而恒光处理对鲜重影响不明显。3种温度胁迫都使总蛋白质含量提高,恒暗胁迫则降低总蛋白质含量,其他胁迫对总蛋白质含量影响不大。累计收集近300份蛋白质样品,建立了水稻蛋白质样品资源库(RiceS-A300),每份样品体积为2 mL,可供200次WB分析,由于试验所采用的条件比较规范且容易控制,可根据需要重复扩大样品的规模,预期不同的实验室采用相同的条件获得的蛋白质样品也能获得可重复的试验结果,利用RiceS-A300调查HSP82蛋白质的表达特征,显示热胁迫明显提高了HSP82的表达,恒光、PEG和盐胁迫也对HSP82的表达有一定影响,而淹胁迫和恒暗处理对HSP82的影响不大。对RiceS-A300资源库的评价扩展了HSP82内参蛋白质的用途。【结论】提出了蛋白质样品资源库建设和评价的流程,建立了第一个版本的胁迫处理条件下的水稻幼苗蛋白质样品资源库(RiceS-A300);在此基础上,通过WB调查了HSP82蛋白质的表达特征,发现其在热胁迫下有特异诱导表达。
DOI:10.1006/bbrc.2000.3114URLPMID:10903912 [本文引用: 1]
A novel rice acidic pathogenesis-related (PR) class 1 cDNA (OsPR1a) was isolated from jasmonic acid (JA)-treated rice seedling leaf. The OsPR1a cDNA is 830 bp long and contains an open reading frame of 507 nucleotides encoding 168 amino acid residues with a predicted molecular mass of 17,560 and pI of 4.4. The deduced amino acid sequence of OsPR1a has a high level of identity with acidic and basic PR1 proteins from plants. Southern analysis revealed that OsPR1a is a member of a multigene family. The OsPR1a gene was found to be cut-inducible, whereas the phytohormones JA, salicylic acid (SA), 3-indoleacetic acid, gibberellin, and ethylene (using ethylene generator ethephon, ET) enhanced accumulation of OsPR1a transcript, as well as the protein phosphatase inhibitors cantharidin (CN) and endothall (EN). Induced expression of OsPR1a gene by JA, CN or EN, and ET was light/dark- and dose-dependent and was almost completely inhibited by cycloheximide. Dark downregulated CN-, EN-, and ET-induced OsPR1a gene expression, whereas it was further enhanced with JA. SA and abscisic acid blocked JA-induced OsPR1a transcript. Simultaneous application of staurosporine (ST) enhances CH- or EN-induced OsPR1a transcript, but not with JA. This is the first report on cloning of a rice acidic PR1 gene (OsPR1a), which is regulated by phytohormones, phosphorylation/dephosphorylation event(s), and light.
[本文引用: 1]
[本文引用: 1]
DOI:10.1002/cpmb.89URLPMID:31237422 [本文引用: 1]
Genetic tools for specific perturbation of endogenous gene expression are highly desirable for interrogation of plant gene functions and improvement of crop traits. Synthetic transcriptional activators derived from the CRISPR/Cas9 system are emerging as powerful new tools for activating the endogenous expression of genes of interest in plants. These synthetic constructs, generated by tethering transcriptional activation domains to a nuclease-dead Cas9 (dCas9), can be directed to the promoters of endogenous target genes by single guide RNAs (sgRNAs) to activate transcription. Here, we provide a detailed protocol for targeted transcriptional activation in plants using a recently developed, highly potent dCas9 gene activator construct referred to as dCas9-TV. This protocol covers selection of sgRNA targets, construction of sgRNA expression cassettes, and screening for an optimal sgRNA using a protoplast-based promoter-luciferase assay. Finally, the dCas9-TV gene activator coupled with the optimal sgRNA is delivered into plants via Agrobacterium-mediated transformation, thereby enabling robust upregulation of target gene expression in transgenic Arabidopsis and rice plants. ? 2019 by John Wiley &amp; Sons, Inc.
DOI:10.1007/s00299-012-1275-3URL [本文引用: 1]
A number of Agrobacterium-mediated rice transformation systems have been developed and widely used in numerous laboratories and research institutes. However, those systems generally employ antibiotics like kanamycin and hygromycin, or herbicide as selectable agents, and are used for the small-scale experiments. To address high-throughput production of transgenic rice plants via Agrobacterium-mediated transformation, and to eliminate public concern on antibiotic markers, we developed a comprehensive efficient protocol, covering from explant preparation to the acquisition of low copy events by real-time PCR analysis before transplant to field, for high-throughput production of transgenic plants of Japonica rice varieties Wanjing97 and Nipponbare using Escherichia coli phosphomannose isomerase gene (pmi) as a selectable marker. The transformation frequencies (TF) of Wanjing97 and Nipponbare were achieved as high as 54.8 and 47.5 %, respectively, in one round of selection of 7.5 or 12.5 g/L mannose appended with 5 g/L sucrose. High-throughput transformation from inoculation to transplant of low copy events was accomplished within 55-60 days. Moreover, the Taqman assay data from a large number of transformants showed 45.2 % in Wanjing97 and 31.5 % in Nipponbare as a low copy rate, and the transformants are fertile and follow the Mendelian segregation ratio. This protocol facilitates us to perform genome-wide functional annotation of the open reading frames and utilization of the agronomically important genes in rice under a reduced public concern on selectable markers.
Key message We describe a comprehensive protocol for large scale production of transgenic Japonica rice plants using non-antibiotic selectable agent, at simplified, cost- and labor-saving manners.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00299-014-1626-3URL [本文引用: 1]
OsiSAP1, an A20/AN1 zinc-finger protein, confers water-deficit stress tolerance at different stages of growth by affecting expression of several endogenous genes in transgenic rice.
Transgenic lines have been generated from rice constitutively expressing OsiSAP1, an A20/AN1 zinc-finger containing stress-associated protein gene from rice, driven by maize UBIQUITIN gene promoter and evaluated for water-deficit stress tolerance at different stages of growth. Their seeds show early germination and seedlings grow better under water-deficit stress compared to non-transgenic (NT) rice. Leaves from transgenic seedlings showed lesser membrane damage and lipid peroxidation under water-deficit stress. Relatively lower rate of leaf water loss has been observed in detached intact leaves from transgenic plants during late vegetative stage. Delayed leaf rolling and higher relative water content were also observed in transgenic plants under progressive water-deficit stress during reproductive developmental stage. Although reduction in grain yield is observed under unstressed condition, the relative water-deficit stress-induced yield losses are lower in transgenic rice vis-A -vis NT plants thereby resulting in yield loss protection. Transcriptome analysis suggests that overexpression of OsiSAP1 in transgenic rice results in altered expression of several endogenous genes including those coding for transcription factors, membrane transporters, signaling components and genes involved in metabolism, growth and development. A total of 150 genes were found to be more than twofold up-regulated in transgenic rice of which 43 genes are known to be involved in stress response. Our results suggest that OsiSAP1 is a positive regulator of water-deficit stress tolerance in rice.
DOI:10.3389/fpls.2017.00993URLPMID:28659944 [本文引用: 1]
SNF 1-RELATED PROTEIN KINASE 2 (SnRK2) is a family of plant-specific protein kinases which is the key regulator of hyper-osmotic stress signaling and abscisic acid (ABA)-dependent development in various plants. Among the rice subclass-I and -II SnRK2s, osmotic stress/ABA-activated protein kinase 2 (SAPK2) may be the primary mediator of ABA signaling. However, SAPK2 has not been comprehensively characterized. In this study, we elucidated the functional properties of SAPK2 using loss-of-function mutants produced with the CRISPR/Cas9 system. The SAPK2 expression level was strongly upregulated by drought, high-salinity, and polyethylene glycol (PEG) treatments. The sapk2 mutants exhibited an ABA-insensitive phenotype during the germination and post-germination stages, suggesting that SAPK2 had a pivotal role related to ABA-mediated seed dormancy. The sapk2 mutants were more sensitive to drought stress and reactive oxygen species (ROS) than the wild-type plants, indicating that SAPK2 was important for responses to drought conditions in rice. An additional investigation revealed that SAPK2 increased drought tolerance in the following two ways: (i) by reducing water loss via the accumulation of compatible solutes, promoting stomatal closure, and upregulating the expression levels of stress-response genes such as OsRab16b, OsRab21, OsbZIP23, OsLEA3, OsOREB1 and slow anion channel (SLAC)-associated genes such as OsSLAC1 and OsSLAC7; (ii) by inducing the expression of antioxidant enzyme genes to promote ROS-scavenging abilities that will ultimately decrease ROS damages. Moreover, we also observed that SAPK2 significantly increased the tolerance of rice plants to salt and PEG stresses. These findings imply that SAPK2 is a potential candidate gene for future crop improvement studies.
DOI:10.3389/fpls.2017.01102URLPMID:28694815 [本文引用: 1]
Drought is one of the major abiotic stresses threatening rice (Oryza sativa) production worldwide. Drought resistance is controlled by multiple genes, and therefore, a multi-gene genetic engineering strategy is theoretically useful for improving drought resistance. However, the experimental evidence for such a strategy is still lacking. In this study, a few drought-responsive genes from rice were assembled by a multiple-round site-specific assembly system, and the constructs were introduced into the rice cultivar KY131 via Agrobacterium-mediated transformation. The transgenic lines of the multi-gene and corresponding single-gene constructs were pre-evaluated for drought resistance. We found that the co-overexpression of two genes, encoding a constitutively active form of a bZIP transcription factor (OsbZIP46CA1) and a protein kinase (SAPK6) involved in the abscisic acid signaling pathway, showed significantly enhanced drought resistance compared with the single-gene transgenic lines and the negative transgenic plants. Single-copy lines of this bi-gene combination (named XL22) and the corresponding single-gene lines were further evaluated for drought resistance in the field using agronomical traits. The results showed that XL22 exhibited greater yield, biomass, spikelet number, and grain number under moderate drought stress conditions. The seedling survival rate of XL22 and the single-gene overexpressors after drought stress treatment also supported the drought resistance results. Furthermore, expression profiling by RNA-Seq revealed that many genes involved in the stress response were specifically up-regulated in the drought-treated XL22 lines and some of the stress-related genes activated in CA1-OE and SAPK6-OE were distinct, which could partially explain the different performances of these lines with respect to drought resistance. In addition, the XL22 seedlings showed improved tolerance to heat and cold stresses. Our results demonstrate that the multi-gene assembly in an appropriate combination may be a promising approach in the genetic improvement of drought resistance.
[本文引用: 1]
[本文引用: 1]
In: Wang G L, Valent B, eds. Advances in Genetics, Genomics and Control of Rice Blast Disease.
[本文引用: 1]
DOI:10.1104/pp.109.152702URLPMID:20130100 [本文引用: 1]
The plant hormone abscisic acid (ABA) is involved in an array of plant processes, including the regulation of gene expression during adaptive responses to various environmental cues. Apart from its well-established role in abiotic stress adaptation, emerging evidence indicates that ABA is also prominently involved in the regulation and integration of pathogen defense responses. Here, we demonstrate that exogenously administered ABA enhances basal resistance of rice (Oryza sativa) against the brown spot-causing ascomycete Cochliobolus miyabeanus. Microscopic analysis of early infection events in control and ABA-treated plants revealed that this ABA-inducible resistance (ABA-IR) is based on restriction of fungal progression in the mesophyll. We also show that ABA-IR does not rely on boosted expression of salicylic acid-, jasmonic acid -, or callose-dependent resistance mechanisms but, instead, requires a functional Galpha-protein. In addition, several lines of evidence are presented suggesting that ABA steers its positive effect on brown spot resistance through antagonistic cross talk with the ethylene (ET) response pathway. Exogenous ethephon application enhances susceptibility, whereas genetic disruption of ET signaling renders plants less vulnerable to C. miyabeanus attack, thereby inducing a level of resistance similar to that observed on ABA-treated wild-type plants. Moreover, ABA treatment alleviates C. miyabeanus-induced activation of the ET reporter gene EBP89, while derepression of pathogen-triggered EBP89 transcription via RNA interference-mediated knockdown of OsMPK5, an ABA-primed mitogen-activated protein kinase gene, compromises ABA-IR. Collectively, these data favor a model whereby exogenous ABA enhances resistance against C. miyabeanus at least in part by suppressing pathogen-induced ET action in an OsMPK5-dependent manner.
.
DOI:10.1093/pcp/pcf111URLPMID:12198199 [本文引用: 1]
Mitogen-activated protein kinase (MAPK) cascades play important roles in signal transduction of extracellular stimuli in eukaryotes. However, stimulatory signals for plant MAPKs have not been well elucidated. Here, a cDNA clone, termed Oryza sativa MAPK4 (OsMAPK4), from rice encoding a protein that showed homology with the eukaryotic MAPKs was isolated. According to the phylogenetic analysis, OsMAPK4 belongs to subgroup IV MAPK in plants. OsMAPK4 transcripts were expressed strongly in mature leaves and weakly in young leaves and panicles. The gene was also differentially expressed in roots at different developmental stages. In addition, the mRNA level of OsMAPK4 was up-regulated under sugar starvation, high salinity and cold treatments. These results suggest that this OsMAPK4 functions not only in developmental programs but also in stress-signaling pathways.
DOI:10.1111/j.1365-313X.2005.02415.xURLPMID:15941394 [本文引用: 1]
Elicitor-triggered transient membrane potential changes and Ca2+ influx through the plasma membrane are thought to be important during defense signaling in plants. However, the molecular bases for the Ca2+ influx and its regulation remain largely unknown. Here we tested effects of overexpression as well as retrotransposon (Tos17)-insertional mutagenesis of the rice two-pore channel 1 (OsTPC1), a putative voltage-gated Ca(2+)-permeable channel, on a proteinaceous fungal elicitor-induced defense responses in rice cells. The overexpressor showed enhanced sensitivity to the elicitor to induce oxidative burst, activation of a mitogen-activated protein kinase (MAPK), OsMPK2, as well as hypersensitive cell death. On the contrary, a series of defense responses including the cell death and activation of the MAPK were severely suppressed in the insertional mutant, which was complemented by overexpression of the wild-type gene. These results suggest that the putative Ca(2+)-permeable channel determines sensitivity to the elicitor and plays a role as a key regulator of elicitor-induced defense responses, activation of MAPK cascade and hypersensitive cell death.
DOI:10.1146/annurev.arplant.59.032607.092740URLPMID:18257711 [本文引用: 1]
Seed dormancy provides a mechanism for plants to delay germination until conditions are optimal for survival of the next generation. Dormancy release is regulated by a combination of environmental and endogenous signals with both synergistic and competing effects. Molecular studies of dormancy have correlated changes in transcriptomes, proteomes, and hormone levels with dormancy states ranging from deep primary or secondary dormancy to varying degrees of release. The balance of abscisic acid (ABA):gibberellin (GA) levels and sensitivity is a major, but not the sole, regulator of dormancy status. ABA promotes dormancy induction and maintenance, whereas GA promotes progression from release through germination; environmental signals regulate this balance by modifying the expression of biosynthetic and catabolic enzymes. Mediators of environmental and hormonal response include both positive and negative regulators, many of which are feedback-regulated to enhance or attenuate the response. The net result is a slightly heterogeneous response, thereby providing more temporal options for successful germination.
DOI:10.1016/s0006-291x(02)02944-3URLPMID:12559953 [本文引用: 1]
Mitogen-activated protein kinase (MAPK) cascade(s) is important for plant defense/stress responses. Though MAPKs have been identified and characterized in rice (Oryza sativa L.), a monocot cereal crop research model, the first upstream component of the kinase cascade, namely MAPK kinase kinase (MAPKKK) has not yet been identified. Here we report the cloning of a novel rice gene encoding a MAPKKK, OsEDR1, designated based on its homology with the Arabidopsis MAPKKK, AtEDR1. OsEDR1, a single copy gene in the genome of rice, encodes a predicted protein with molecular mass of 113046.13 and a pI of 9.03. Using our established two-week-old rice seedling in vitro model system, we show that OsEDR1 has a constitutive expression in seedling leaves and is further up-regulated within 15 min upon wounding by cut, treatment with the global signals jasmonic acid (JA), salicylic acid (SA), ethylene (ethephon, ET), abscisic acid, and hydrogen peroxide. In addition, protein phosphatase inhibitors, fungal elicitor chitosan, drought, high salt and sugar, and heavy metals also dramatically induce its expression. Moreover, OsEDR1 expression was altered by co-application of JA, SA, and ET, and required de novo synthesized protein factor(s) in its transient regulation. Furthermore, using an in vivo system we also show that OsEDR1 responds to changes in temperature and environmental pollutants-ozone and sulfur dioxide. Finally, OsEDR1 expression varied significantly in vegetative and reproductive tissues. These results suggest a role for OsEDR1 in defense/stress signalling pathways and development.
DOI:10.1242/jcs.00175URLPMID:12432076 [本文引用: 1]
The phytohormone abscisic acid (ABA) plays important regulatory roles in many plant developmental processes including seed dormancy, germination, growth, and stomatal movements. These physiological responses to ABA are in large part brought about by changes in gene expression. To study genome-wide ABA-responsive gene expression we applied massively parallel signature sequencing (MPSS) to samples from Arabidopsis thaliana wildtype (WT) and abi1-1 mutant seedlings. We identified 1354 genes that are either up- or downregulated following ABA treatment of WT seedlings. Among these ABA-responsive genes, many encode signal transduction components. In addition, we identified novel ABA-responsive gene families including those encoding ribosomal proteins and proteins involved in regulated proteolysis. In the ABA-insensitive mutant abi1-1, ABA regulation of about 84.5% and 6.9% of the identified genes was impaired or strongly diminished, respectively; however, 8.6% of the genes remained appropriately regulated. Compared to other methods of gene expression analysis, the high sensitivity and specificity of MPSS allowed us to identify a large number of ABA-responsive genes in WT Arabidopsis thaliana. The database given in our supplementary material (http://jcs.biologists.org/supplemental) provides researchers with the opportunity to rapidly assess whether genes of interest may be regulated by ABA. Regulation of the majority of the genes by ABA was impaired in the ABA-insensitive mutant abi1-1. However, a subset of genes continued to be appropriately regulated by ABA, which suggests the presence of at least two ABA signaling pathways, only one of which is blocked in abi1-1.
DOI:10.1016/j.cell.2006.05.050URLPMID:16901781 [本文引用: 1]
Small-molecule hormones govern every aspect of the biology of plants. Many processes, such as growth, are regulated in similar ways by multiple hormones, and recent studies have revealed extensive crosstalk among different hormonal signaling pathways. These results have led to the proposal that a common set of signaling components may integrate inputs from multiple hormones to regulate growth. In this study, we tested this proposal by asking whether different hormones converge on a common set of transcriptional targets in Arabidopsis seedlings. Using publicly available microarray data, we analyzed the transcriptional effects of seven hormones, including abscisic acid, gibberellin, auxin, ethylene, cytokinin, brassinosteroid, and jasmonate. A high-sensitivity analysis revealed a surprisingly low number of common target genes. Instead, different hormones appear to regulate distinct members of protein families. We conclude that there is not a core transcriptional growth-regulatory module in young Arabidopsis seedlings.
DOI:10.1007/s10142-002-0070-6URLPMID:12444421 [本文引用: 1]
Full-length cDNAs are essential for functional analysis of plant genes. Recently, cDNA microarray analysis has been developed for quantitative analysis of global and simultaneous analysis of expression profiles. Microarray technology is a powerful tool for identifying genes induced by environmental stimuli or stress and for analyzing their expression profiles in response to environmental signals. We prepared an Arabidopsis full-length cDNA microarray containing around 7,000 independent full-length cDNA groups and analyzed the expression profiles of genes. The transcripts of 245, 299, 54 and 213 genes increased after abscisic acid (ABA), drought-, cold-, and salt-stress treatments, respectively, with inducibilities more than fivefold compared with those of control genes [corrected]. The cDNA microarray analysis showed that many ABA-inducible genes were induced after drought- and high-salinity-stress treatments, and that there is more crosstalk between drought and ABA responses than between ABA and cold responses. Among the ABA-inducible genes identified, we identified 22 transcription factor genes, suggesting that many transcriptional regulatory mechanisms exist in the ABA signal transduction pathways.
DOI:10.1021/acschembio.9b00453URLPMID:31497942 [本文引用: 1]
The plant hormone abscisic acid (ABA) regulates the development of various plant organs including seeds, roots, and fruits, and significantly contributes to abiotic stress responses, especially to drought. Since recent climate changes are adversely affecting crop cultivation, enhancement of plant stress tolerance by regulation of ABA signaling would be an important strategy. In the plant genome, ABA receptors are encoded by multiple genes constituting three subfamilies; however, functional differences among them remain unclear. To enhance desired effects of ABA, the biological functions of the receptor family warrant clarification. This study aimed to determine the functional differences among ABA receptors in plants. We screened small-molecule ligands binding to specific receptors, using a chemical array. In vitro evaluation of hit compounds using 11 Arabidopsis ABA receptors revealed that (+)-3'-alkyl ABAs served as agonists for different receptors depending on the length of their 3'-alkyl chains. Combinatorial in vitro and physiological effects of these compounds on the stomata, seeds, and seedlings indicated that, along with subfamily III, receptors of subfamily II are important to induce strong drought responses. Among (+)-3'-alkyl ABAs assessed herein, (+)-3'-butyl ABA induced a transcriptional response and stomatal closure but only slightly inhibited seed germination and growth, suggesting that it enhances drought tolerance. In silico docking simulation and site-directed mutagenesis revealed the amino acid residues contributing to the selective agonist activity of the (+)-3'-alkyl ABAs. These results provide novel insights into the structure and biological effects of 3'-derivatives of ABA and a basis for agrochemical development.
DOI:10.1016/s0092-8674(01)00606-7URLPMID:11747807 [本文引用: 1]
The plant hormone abscisic acid (ABA) regulates the aperture of the stomatal pore. The recent identification of new intermediates involved in ABA signaling suggests that this complex pathway is organized as a module-based network.
DOI:10.1093/jxb/erq243URLPMID:20693409 [本文引用: 1]
Brassinosteroids (BRs) have been shown to induce hydrogen peroxide (H(2)O(2)) accumulation, and BR-induced H(2)O(2) up-regulates antioxidant defence systems in plants. However, the mechanisms by which BR-induced H(2)O(2) regulates antioxidant defence systems in plants remain to be determined. In the present study, the role of ZmMPK5, a mitogen-activated protein kinase, in BR-induced anitioxidant defence and the relationship between the activation of ZmMPK5 and H(2)O(2) production in BR signalling were investigated in leaves of maize (Zea mays) plants. BR treatment activated ZmMPK5, induced apoplastic and chloroplastic H(2)O(2) accumulation, and enhanced the total activities of antioxidant enzymes. Such enhancements were blocked by pre-treatment with mitogen-activated protein kinase kinase (MAPKK) inhibitors and H(2)O(2) inhibitors or scavengers. Pre-treatment with MAPKK inhibitors substantially arrested the BR-induced apoplastic H(2)O(2) production after 6 h of BR treatment, but did not affect the levels of apoplastic H(2)O(2) within 1 h of BR treatment. BR-induced gene expression of NADPH oxidase was also blocked by pre-treatment with MAPKK inhibitors and an apoplastic H(2)O(2) inhibitor or scavenger after 120 min of BR treatment, but was not affected within 30 min of BR treatment. These results suggest that the BR-induced initial apoplastic H(2)O(2) production activates ZmMPK5, which is involved in self-propagation of apoplastic H(2)O(2) via regulation of NADPH oxidase gene expression in BR-induced antioxidant defence systems.
DOI:10.1111/j.1365-313X.2008.03433.xURLPMID:18248592 [本文引用: 1]
Catalase controls cellular H(2)O(2) and plays important roles in the adaptation of plants to various stresses, but little is known about the signaling events that lead to the expression of CAT1 and the production of H(2)O(2). Here we report the dependence of CAT1 expression and H(2)O(2) production on a mitogen-activated protein kinase (MAPK) cascade. CAT1 transcript was induced in an ABA-dependent way and the induction was abolished in the T-DNA insertion mutant mkk1 (SALK_015914), while AtMKK1 overexpression significantly enhanced the ABA-induced CAT1 expression and H(2)O(2) production. AtMPK6, another component in the MAPK cascade, was also involved: mpk6 mutant blocked and overexpressing AtMPK6 enhanced the ABA-dependent expression of CAT1 and H(2)O(2) production. The activity of AtMPK6 was increased by ABA in an AtMKK1-dependent manner. These data clearly suggest an ABA-dependent signaling pathway connecting CAT1 expression through a phosphorelay including AtMKK1 and AtMPK6. In further support of this view, mkk1 mutant reduced both the sensitivity to ABA during germination and the drought tolerance of seedlings, whereas the AtMKK1 overexpression line showed the opposite responses when compared with the wild type. The data suggest AtMKK1-AtMPK6 to be a key module in an ABA-dependent signaling cascade causing H(2)O(2) production and stress responses.
DOI:10.1073/pnas.0907205106URLPMID:19910530 [本文引用: 1]
Reactive oxygen species (ROS) mediate abscisic acid (ABA) signaling in guard cells. To dissect guard cell ABA-ROS signaling genetically, a cell type-specific functional genomics approach was used to identify 2 MAPK genes, MPK9 and MPK12, which are preferentially and highly expressed in guard cells. To provide genetic evidence for their function, Arabidopsis single and double TILLING mutants that carry deleterious point mutations in these genes were isolated. RNAi-based gene-silencing plant lines, in which both genes are silenced simultaneously, were generated also. Mutants carrying a mutation in only 1 of these genes did not show any altered phenotype, indicating functional redundancy in these genes. ABA-induced stomatal closure was strongly impaired in 2 independent RNAi lines in which both MPK9 and MPK12 transcripts were significantly silenced. Consistent with this result, mpk9-1/12-1 double mutants showed an enhanced transpirational water loss and ABA- and H(2)O(2)-insensitive stomatal response. Furthermore, ABA and calcium failed to activate anion channels in guard cells of mpk9-1/12-1, indicating that these 2 MPKs act upstream of anion channels in guard cell ABA signaling. An MPK12-YFP fusion construct rescued the ABA-insensitive stomatal response phenotype of mpk9-1/12-1, demonstrating that the phenotype was caused by the mutations. The MPK12 protein is localized in the cytosol and the nucleus, and ABA and H(2)O(2) treatments enhance the protein kinase activity of MPK12. Together, these results provide genetic evidence that MPK9 and MPK12 function downstream of ROS to regulate guard cell ABA signaling positively.
DOI:10.1007/s00425-008-0848-4URL [本文引用: 1]
Mitogen-activated protein kinase (MAPK) cascades are involved in biotic and abiotic stress responses. In plants, MAPKs are classified into four groups, designated A–D. Information about group C MAPKs is limited, and, in particular, no data from maize are available. In this article, we isolated a novel group C MAPK gene, ZmMPK7, from Zea mays. Exogenous abscisic acid (ABA) and hydrogen peroxide (H2O2) induced calcium-dependant transcription of ZmMPK7. Induction of this gene in response to ABA was blocked by several reactive oxygen species (ROS) manipulators such as imidazole, Tiron, and dimethylthiourea (DMTU). This result indicates that endogenous H2O2 may be required for ZmMPK7-mediated ABA signaling. Expression of ZmMPK7 in Nicotonia tobaccum caused less H2O2 to accumulate and alleviated ROS-mediated injuries following submission of the plants to osmotic stress. The enhanced total peroxidase (POD) activity in transgenic tobacco plants may contribute to removal of ROS. Finally, we have shown that the ZmMPK7 protein localizes in the nucleus. These results broaden our knowledge regarding plant group C MAPK activity in response to stress signals.
DOI:10.1105/tpc.9.5.809URLPMID:9165755 [本文引用: 1]
The involvement of phosphorylation/dephosphorylation in the salicylic acid (SA) signal transduction pathway leading to pathogenesis-related gene induction has previously been demonstrated using kinase and phosphatase inhibitors. Here, we show that in tobacco suspension cells, SA induced a rapid and transient activation of a 48-kD kinase that uses myelin basic protein as a substrate. This kinase is called the p48 SIP kinase (for SA-Induced Protein kinase). Biologically active analogs of SA, which induce pathogenesis-related genes and enhanced resistance, also activated this kinase, whereas inactive analogs did not. Phosphorylation of a tyrosine residue(s) in the SIP kinase was associated with its activation. The SIP kinase was purified to homogeneity from SA-treated tobacco suspension culture cells. The purified SIP kinase is strongly phosphorylated on a tyrosine residue(s), and treatment with either protein tyrosine or serine/threonine phosphatases abolished its activity. Using primers corresponding to the sequences of internal tryptic peptides, we cloned the SIP kinase gene. Analysis of the SIP kinase sequence indicates that it belongs to the MAP kinase family and that it is distinct from the other plant MAP kinases previously implicated in stress responses, suggesting that different members of the MAP kinase family are activated by different stresses.
DOI:10.1111/j.1365-313X.2006.03003.xURLPMID:17253983 [本文引用: 1]
In tobacco (Nicotiana tabacum), wounding causes rapid activation of two mitogen-activated protein kinases (MAPKs), wound-induced protein kinase (WIPK) and salicylic acid (SA)-induced protein kinase (SIPK), and the subsequent accumulation of jasmonic acid (JA). Our previous studies suggested that activation of WIPK is required for the production of wound-induced JA. However, the exact role of WIPK remains unresolved. We generated transgenic tobacco plants in which either WIPK or SIPK were silenced using RNA interference to define the roles of WIPK and SIPK in the wound response. In addition, transgenic tobacco plants were generated in which both WIPK and SIPK were silenced to examine the possibility that they have redundant roles. Wound-induced JA production was reduced compared with non-silenced plants in all of the WIPK-, SIPK- and WIPK/SIPK-silenced plants. Transgenic plants over-expressing NtMKP1, a gene encoding tobacco MAPK phosphatase, which inactivates WIPK and SIPK, also exhibited reduced JA production in response to wounding. In both WIPK/SIPK-silenced and NtMKP1-over-expressing plants, wounding resulted in an abnormal accumulation of both SA and transcripts for SA-responsive genes. These results suggest that WIPK and SIPK play an important role in JA production in response to wounding, and that they function cooperatively to control SA biosynthesis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}