 ,2,3,*, 卓嘎,1,*
,2,3,*, 卓嘎,1,*Growth habit identification and diversity and stability analysis of heading date in Tibetan barley (Hordeum vulgare L.)
Lhundrupnamgyal 1,2,3,4,5, LI Hui-Hui5, GUO Gang-Gang5, Chemiwangmo 2,3, GAO Li-Yun2,3, TANG Ya-Wei2,3, Nyematashi 2,4, Dawadondrup ,2,3,*, Dolkar ,1,*通讯作者:
收稿日期:2019-03-20接受日期:2019-06-22网络出版日期:2019-07-15
| 基金资助: |
Received:2019-03-20Accepted:2019-06-22Online:2019-07-15
| Fund supported: |
作者简介 About authors
伦珠朗杰,E-mail:lhundrupnamgyal@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (4299KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
伦珠朗杰, 李慧慧, 郭刚刚, 其美旺姆, 高丽云, 唐亚伟, 尼玛扎西, 达瓦顿珠, 卓嘎. 西藏青稞冬春性鉴定及抽穗期多样性与稳定性分析[J]. 作物学报, 2019, 45(12): 1796-1805. doi:10.3724/SP.J.1006.2019.91025
Lhundrupnamgyal , LI Hui-Hui, GUO Gang-Gang, Chemiwangmo , GAO Li-Yun, TANG Ya-Wei, Nyematashi , Dawadondrup , Dolkar .
抽穗期是作物重要的驯化和农艺性状, 抽穗期长短与作物产量和品质密切相关[1,2,3,4]。作物从起源中心向外传播和驯化选择时, 只有在恰当的时间抽穗, 才能使其正常生存和繁衍后代[5,6]。大麦存在3个抽穗期信号通路, 一是低温感知, 即春化敏感性, 将大麦分为冬性和春性; 二是光周期响应通路, 长日照促进开花; 三是早熟相关基因调控通路, 分为早熟型和晚熟型[7,8,9,10]。前2个通路受外界环境影响, 环境与基因互作调控开花, 后者主要由自身遗传决定[11]。目前, 通过图位克隆或同源克隆手段, 已经分离鉴定出抽穗期的关键基因及基因家族, 如春化基因VRN-H1、VRN-H2和光周期基因PPD-H1、PPD-H2以及FT、CO等, 这极大促进了大麦种质资源的鉴定与利用[12,13,14,15,16]。
青稞(裸大麦)是青藏高原地区藏民主要的粮食作物, 在西藏青稞播种面积和产量分别约占粮食总播种面积和产量的60%和55%, 同时也是农区饲草的主要来源[17,18,19,20]。青稞具有较好的适应性和抗逆性, 在农业生产和生态环境恶劣的西藏边境农区或海拔4700 m以上的高寒农区, 青稞是唯一能正常收获的粮食作物[20]。自1952年以来, 农业科研人员对西藏农作物种质资源多次考察和收集, 迄今共收集西藏大麦青稞种质资源7000余份, 大麦科技工作者对青稞种质进行了变种分类、多样性评价等系统研究, 经田间种植、性状观察和鉴定编目后的3602份保存于国家农作物种质资源长期库[21,22,23]。为青稞资源的遗传研究和利用奠定了基础。
西藏平均海拔在4000 m以上, 地理气候生态环境多样, 包括秋播区、冬春兼作区和春播区3个主要生态区[21], 到目前为止, 尚不明确不同来源的青稞种质在西藏各生态区的生长特性和环境适应性, 限制了多样性地方品种在提高青稞育成品种的生态适应性方面的有效利用[24]。为此, 本研究利用1605份西藏青稞地方品种、育成品种及青海、四川等周边藏区代表性育成品种, 在多种环境鉴定冬春性和抽穗期, 研究不同环境中青稞抽穗期的变化规律, 为青稞种质资源的利用与广适应性青稞品种的选育提供依据。
1 材料与方法
1.1 试验材料
本研究的青稞种质资源共1605份, 包括西藏地方品种1505份及育成品种70份、周边藏区育成品种30份(青海19份、四川甘孜5份、甘肃甘南5份, 云南迪庆1份), 育成品种中有93份春性品种和7份冬性品种。以西藏主推春青稞品种藏青2000、藏青320、喜玛拉19号作为春性品种对照, 冬青稞品种冬青18号和果洛作为冬性品种对照。地方品种覆盖西藏所有生态区, 育成品种包括近50年选育的春青稞和冬青稞代表性品种。1.2 试验方法
2017年10月和2018年3月, 于西藏农牧学院植物科学院试验基地(西藏林芝, 海拔2900 m, 94°34′E, 26°67′N)秋播和春播。在西藏农牧科学院农业研究所试验地(西藏拉萨, 海拔3660 m, 91°04′E, 29°64′N)进行冬春性鉴定分2个播期, 春播I (当地正常播期, 2018年4月12日)和春播II (比正常播期晚10 d, 2018年4月22日)。每份材料种1行, 行长1.5 m, 每行25粒, 行距为30 cm。以每行材料中1/2的穗子抽出旗叶鞘一半的时间记为抽穗期[25,26]。参照Hemming等[27]的方法, 春播出苗后100 d作为区分冬春性的抽穗期记载期限。拉萨2个播期均正常抽穗的记为春性, 春播I能够拔节或者极晚抽穗, 春播II不能抽穗, 甚至不能拔节的作为半冬性, 2个春播均不能抽穗的记为冬性[24,28](图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1拉萨冬春性鉴定表型
Fig. 1Identification of spring/winter growth habit in Lhasa
1.3 数据分析
采用Microsoft Excel 2016和R语言整理和分析数据, 采用QTL IciMapping软件中表型数据ANOVA功能分析遗传力和相关性。利用Shannon-Wiener’s遗传多样性指数(Shannon-Wiener diversity index, H')进行遗传多样性评价[29]。通过计算参试材料的平均值(μ)和标准差(σ), 把抽穗期划分为5等级, 早抽穗I (X < μ-1.5σ)、偏早抽穗II (μ-1.5σ < X < μ-0.5σ)、中间型III (μ-0.5σ < X < μ+0.5σ)、偏晚抽穗IV (μ+0.5σ < X < μ+1.5σ)和晚抽穗V (X > μ+1.5σ), X为每份材料的抽穗期。根据种质类型和生长习性, 将品种划分为春性地方品种、春性育成品种、冬性地方品种和冬性育成品种4个类型。${H}'=-\sum{{{p}_{i}}\ln {{p}_{i}}}$
式中, H'为Shannon-Wiener’s遗传多样性指数, pi为第i级中的材料份数占该类型品种总份数的百分比。采用R语言multcomp包Tukey方法进行多重比较。
对不同环境和播期的抽穗期进行标准化, 以便不同等级之间的比较。按(X-μ)/σ标准化, 式中X为不同播期某一个参试材料的抽穗期, μ为播期内参试材料的平均值, σ为标准差。
2 结果与分析
2.1 青稞种质资源冬春性鉴定与地理分布
绝大多数青稞地方品种和育成品种均为春性类型(表1)。青稞地方品种中春性共有1448份, 冬性共有57份, 分别占地方品种的96.2%和3.8%; 春性育成品种共有93份与记载吻合; 冬性育成品种有7份, 在本研究中均表现为半冬性, 冬性育成品种中未发现强冬性类型。Table 1
表1
表1材料来源及品种类型
Table 1
| 来源 Origin | 类型 Type | 品种数 No. of accessions | 合计 Total | 比例 Proportion (%) |
|---|---|---|---|---|
| 拉萨 Lhasa | 春性地方品种 Spring landrace (SL) | 125 | 155 | 9.64 |
| 冬性地方品种 Winter landrace (WL) | 3 | |||
| 春性育成品种 Spring variety (SV) | 21 | |||
| 冬性育成品种 Winter variety (WV) | 6 | |||
| 山南 Lhoka | 春性地方品种 Spring landrace (SL) | 355 | 377 | 23.45 |
| 冬性地方品种 Winter landrace (WL) | 12 | |||
| 春性育成品种 Spring variety (SV) | 10 | |||
| 日喀则 Shigatse | 春性地方品种 Spring landrace (SL) | 538 | 573 | 35.63 |
| 冬性地方品种 Winter landrace (WL) | 6 | |||
| 春性育成品种 Spring variety (SV) | 29 | |||
| 昌都 Chamdo | 春性地方品种 Spring landrace (SL) | 275 | 282 | 17.60 |
| 冬性地方品种 Winter landrace (WL) | 6 | |||
| 春性育成品种 Spring variety (SV) | 1 | |||
| 林芝 Nyingchi | 春性地方品种 Spring landrace (SL) | 114 | 147 | 9.14 |
| 冬性地方品种 Winter landrace (WL) | 30 | |||
| 春性育成品种 Spring variety (SV) | 2 | |||
| 冬性育成品种 Winter variety (WV) | 1 | |||
| 阿里 Ngari | 春性地方品种 Spring landrace (SL) | 40 | 40 | 2.49 |
| 那曲 Nagqu | 春性地方品种 Spring landrace (SL) | 1 | 1 | 0.06 |
| 西藏其他地区 Other Tibetan areas | 春性育成品种 Spring variety (SV) | 30 | 30 | 1.87 |
新窗口打开|下载CSV
春性是青稞资源的主要特征, 分布在西藏所有生态区, 冬性地方品种主要分布在林芝, 共30份; 其次在山南、昌都和日喀则, 分别有12、6和6份; 在拉萨只有3份材料。以牧业为主的阿里和那曲地区, 收集资源仅有41份, 未发现冬性类型(图2)。目前生产上推广的7份半冬性品种主要种植在拉萨、山南和林芝等海拔3700 m以下农区。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2青稞冬春性资源的地理分布
灰色线条表示雅鲁藏布江流域, 饼图大小表示在总材料中所占的比重。
Fig. 2Geographical distribution of different growth habit germplasm
Gray line indicates the Yarlung Zangbo River basin, and the pie chart size represents the proportion to the entire material.
2.2 青稞种质资源抽穗期的多样性
2.2.1 2017年林芝秋播抽穗期 林芝秋播条件下, 除116份春性地方品种和2份冬性地方品种不能正常越冬未获得数据, 其余1487份材料均能正常抽穗。春性地方品种共有1332份, 平均抽穗期为174.54± 9.33 d, 其中40份(日喀则21份、山南10份、拉萨5份、昌都4份)抽穗期最早, 为160 d, 来源于日喀则的3份(2份谢通门县、1份定结县)抽穗期最晚, 为204 d。冬性地方品种共有55份, 平均抽穗期值为182.86± 10.04 d, 其中抽穗最早的有3份(林芝2份、日喀则1份), 抽穗期为161 d, 而来自西藏林芝的品种乃那(黑青稞)抽穗期最晚, 为202 d。春性育成品种共有93份, 抽穗期平均值为174.40±6.68 d, 其中青海的2份(北青3号、北青4号)最早抽穗, 抽穗期为160 d, 林芝的紫芒抽穗期最晚, 为202 d。半冬性育成品种共有7份, 平均抽穗期为185.33±8.36 d, 冬青8号抽穗最早, 为180 d, 果洛抽穗期最晚, 为202 d。
2.2.2 2018年林芝春播 1605份青稞资源中除6份冬性地方品种、1份春性育成品种不能正常抽穗外, 其余1598份材料在林芝春播均能抽穗。春性地方品种共有1448份, 抽穗期平均值为73.16±6.33 d, 其中来源西藏日喀则的2份抽穗期最早, 为59 d, 来源林芝的扎卓骨和山南隆子县的变芒紫六棱抽穗期最晚, 为94 d。冬性地方品种共有52份, 平均抽穗期为 84.72±6.39 d, 其中来源拉萨的果洛抽穗期最早, 为64 d, 来源山南的勾芒白青稞的抽穗期最晚, 为97 d。
春性育成品种共有92份, 抽穗期平均值为75.78±5.50 d, 其中甘南的甘青1号抽穗期最早, 为62 d, 拉萨的拉萨勾芒、日喀则的喜玛拉8号、喜玛拉9号和喜玛拉10号4个品种的抽穗期最晚, 为85 d。半冬性育成品种共有6份, 平均抽穗期值80.6±7.09 d, 其中冬青1号抽穗期最早, 为71 d, 林芝的果洛抽穗期最晚, 为88 d。
2.2.3 2018年拉萨春播 1605份青稞资源中, 除56份冬性品种外, 其他均能正常抽穗。冬性品种中7份半冬性品种主茎能正常抽穗并记录抽穗期。综合两个播期的抽穗期计算其广义遗传力(H2), 为0.82。
1448份春性地方品种的抽穗期平均值为68.19± 5.62 d, 其中, 最早抽穗的来源于西藏日喀则的吉隆青稞, 抽穗期为53 d, 而来自西藏山南的左线那果(黑青稞)抽穗期最晚, 为85 d。冬性地方品种共有4份能够正常抽穗, 抽穗期平均值为79.00±9.17 d, 其中来源西藏日喀则萨迦县的紫青稞最早抽穗, 抽穗期为69 d, 而来自西藏山南市错那县的白青稞抽穗期最晚, 为87 d。春性育成品种共有93份, 抽穗期平均值为70.18±4.19d, 变幅为59~81 d; 其中, 北青7号抽穗期最早, 为59 d, 藏青7239号抽穗期最晚, 为81 d。4份半冬性育成品种冬青1号、冬青8号、冬青11号、冬青19号能够记载抽穗期, 其平均值为73.75±3.28 d, 变幅为71~78 d。
在不同环境和播期条件下, 四种类型品种的抽穗期符合正态分布(图3), 并且具有较高的多样性。总体而言, 地方品种的多样性高于育成品种, 多样性最高的是在拉萨春播环境, 其次是林芝春播环境, 而林芝秋播环境下多样性最低, 且春播环境的变异系数大于秋播环境(表2)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3青稞资源在3个环境的抽穗期分布
黑色虚线表示抽穗期平均值, SL: 春性地方品种; SV: 春性育成品种; WL: 冬性地方品种; WV:冬性育成品种。2017年林芝秋播, 2018年林芝春播, 2018年拉萨春播。
Fig. 3Heading date distribution of Tibet barley germplasm in three environments
Gray dotted line indicates the averaged value. SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
Table 2
表2
表2青稞资源的抽穗期性状变化及分布特征
Table 2
| 试验 Trial | 品种类型 Variety type | 品种数 No. of samples | 平均值 Mean±SD | 变幅 Range | 变异系数CV (%) | 多样性指数Shannon H° |
|---|---|---|---|---|---|---|
| 2017年林芝秋播2017 Nyingchi-A | 春性地方品种 Spring landrace (SL) | 1332 | 174.54±9.33 | 160.00-204.00 | 5.53 | 1.33 |
| 冬性地方品种 Winter landrace (WL) | 55 | 182.86±10.04 | 161.00-200.00 | 5.49 | 1.00 | |
| 春性育成品种 Spring variety (SV) | 93 | 174.40±6.68 | 160.00-202.00 | 3.83 | 1.13 | |
| 冬性育成品种 Winter variety (WV) | 7 | 185.33±8.36 | 180.00-202.00 | 4.51 | 0.56 | |
| 2018年林芝春播2018 Nyingchi-S | 春性地方品种 Spring landrace (SL) | 1448 | 73.16±6.33 | 59.00-94.00 | 8.65 | 1.37 |
| 冬性地方品种 Winter landrace (WL) | 52 | 84.72±6.39 | 64.00-97.00 | 7.55 | 1.25 | |
| 春性育成品种 Spring variety (SV) | 92 | 75.78±5.50 | 62.00-85.00 | 7.26 | 0.96 | |
| 冬性育成品种 Winter variety (WV) | 6 | 80.60±7.09 | 71.00-88.00 | 8.80 | 0.73 | |
| 2018年拉萨春播2018 Lhasa-S | 春性地方品种 Spring landrace (SL) | 1448 | 68.19±5.62 | 53.00-85.00 | 8.36 | 1.41 |
| 冬性地方品种 Winter landrace (WL) | 4 | 79.00±9.17 | 69.00-87.00 | 11.60 | 1.15 | |
| 春性育成品种 Spring variety (SV) | 93 | 70.18±4.19 | 59.00-81.00 | 6.04 | 0.64 | |
| 冬性育成品种 Winter variety (WV) | 4 | 73.75±3.28 | 71.00-75.00 | 4.45 | 0.69 |
新窗口打开|下载CSV
2.3 青稞种质资源抽穗期的比较分析
2.3.1 相同播种条件下不同类型青稞资源抽穗期比较 在林芝秋播条件下, 春性品种较冬性品种早抽穗, 且达到极显著水平(P<0.01), 地方品种和育成品种中具有相同的趋势。但生长习性相同的地方品种和育成品种之间无显著差异。在林芝春播条件下, 春性地方品种最早抽穗, 且与其他3个类型品种的差异达到极显著水平(P<0.01), 其次是春性育成品种, 与冬性地方品种的差异达到显著水平(P<0.01), 与冬性育成品种无显著差异, 冬性地方品种与冬性育成品种也没有显著差异。在拉萨春播条件下, 抽穗期由早到晚依次是春性地方品种、春性育成品种、冬性地方品种, 两两之间差异均达到极显著水平(P<0.01), 而冬性育成品种与其他三类品种之间没有显著差异(图4)。图4
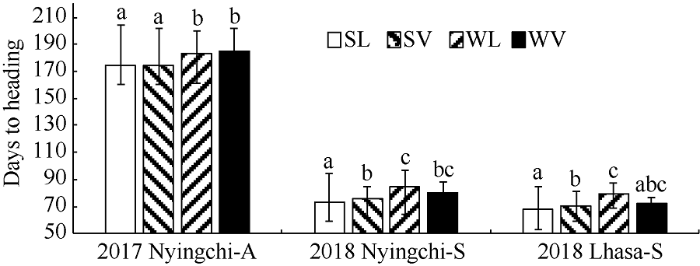 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同品种在秋播和春播条件下抽穗期的比较
SL: 春性地方品种; SV: 春性育成品种; WL: 冬性地方品种; WV: 冬性育成品种。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉萨春播。不同字母表示处理间差异显著水平(P < 0.01)。
Fig. 4Comparison of heading date of different growth habit germplasm in three sowing times
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown. Significant difference (P < 0.01).
2.3.2 不同环境条件下相同类型青稞资源抽穗期比较 在2个春播条件下, 地方品种在拉萨较在林芝更早抽穗, 且达到极显著水平(P<0.01), 春性品种和冬性品种存在相同的规律, 但是育成品种在2个春播条件下没有显著差异(图5)。
图5
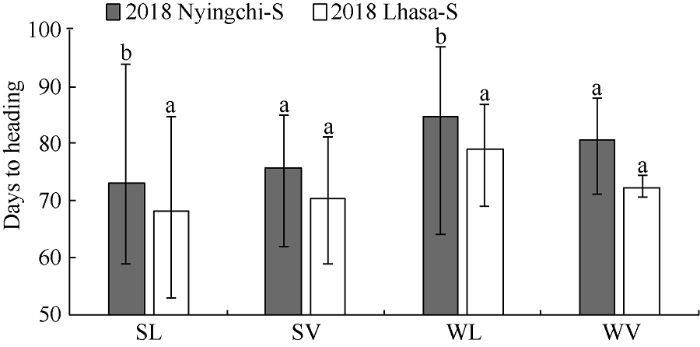 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5春播条件下不同环境间抽穗期的比较
SL: 春性地方品种; SV: 春性育成品种; WL: 冬性地方品种; WV:冬性育成品种。2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉萨春播。不同字母表示处理间差异显著水平(P<0.01)。
Fig. 5Comparison of heading date between different environments under spring sowing conditions
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown. Significant difference (P<0.01).
2.4 抽穗期在不同环境中的相关性分析
3个环境抽穗期两两之间存在极显著的正相关。其中, 2018年林芝和2018年拉萨2个春播之间相关系数为0.79, 明显高于两者与2017年林芝秋播之间的相关系数; 但2个春播与2017年秋播之间的相关系数很相近, 分别为0.40和0.43 (图6)。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同播期和环境间抽穗期的相关性
A、B、C表示播期间或环境间的相关系数。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉萨春播。
Fig. 6Correlation between heading dates among different sowing time and environments
A, B, C represents the correlation coefficient of sowing time or environments. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
2.5 品种在抽穗期各等级中的分布
抽穗期主要集中在II~IV等级内, 即偏早抽穗、中间型和偏晚抽穗类型, 占2017年林芝秋播89.87%、2018年林芝春播87.13%和2018年拉萨春播的86.13%, I和V两等级内所包含的品种量比较少, 只占10.13%、12.87%、13.87% (图7)。图7
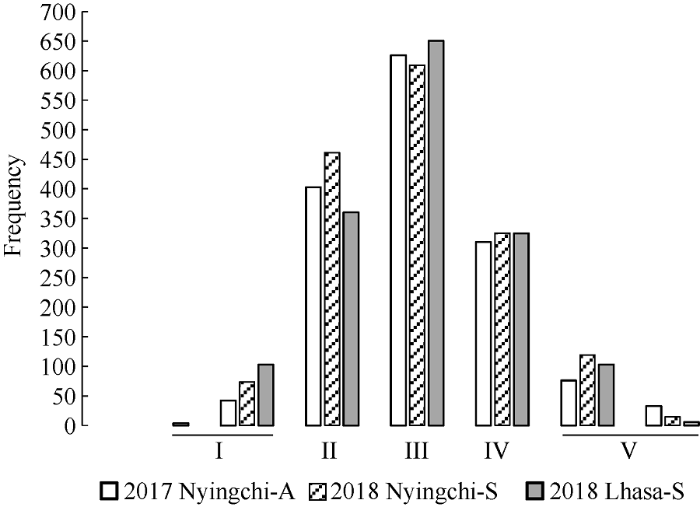 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同播期抽穗期值标准化的五个等级的变化频率
I: 早抽穗; II: 偏早抽穗; III:中间型; IV: 偏晚抽穗; V: 晚抽穗。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉萨春播。
Fig. 7Frequency of five grades after standardization of heading date values in different sowing times
I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
2.6 青稞种质资源不同环境之间的稳定性分析
2.6.1 秋播和春播抽穗期稳定性分析 青稞资源在所有条件下同一个等级内共享或稳定性较高的品种共有390份品种, 分别为I(3)、II(123)、III(199)、IV(55)、V(10); 其中, 春性地方品种中稳定性较高的品种共有368份品种, 分别为I(3)、II(122)、III(185)、IV(48)、V(10); 春性育成品种中稳定性较高的品种共有21份, 分别为II(1)、III(14)、IV(6); 冬性地方品种和冬性育成品种中只有冬青11号在所有环境中稳定在IV, 冬性地方品种无早抽穗类型(I), 冬性育成品种中未发现早抽穗类型(I)和偏早抽穗类型(II) (图8)。图8
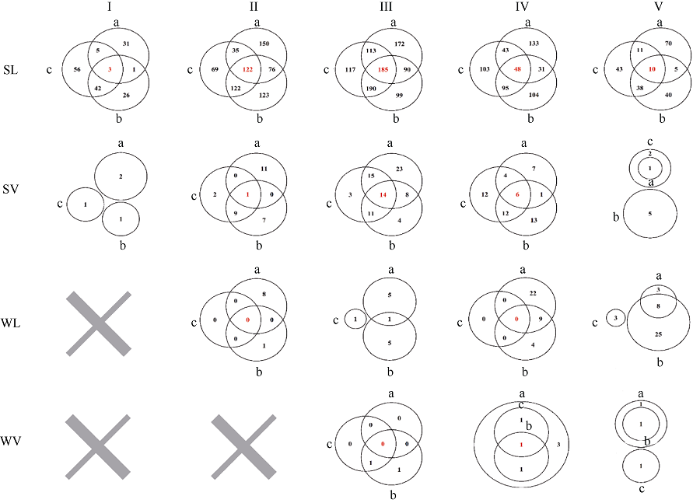 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同品种在不同播期和环境条件下抽穗期的稳定性分析
SL: 春性地方品种; SV: 春性育成品种; WL: 冬性地方品种; WV: 冬性育成品种。I: 早抽穗; II: 偏早抽穗; III: 中间型; IV: 偏晚抽穗; V: 晚抽穗。a: 2017年林芝秋播; b: 2018年林芝春播; c: 2018年拉萨春播。红色数字表示3个环境内共有的品种数。
Fig. 8Stability analysis of different varieties in heading dates across different sowing times and environmental conditions
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. a: 2017 Nyingchi-Autumn sown; b: 2018 Nyingchi-Spring sown; c: 2018 Lhasa-Spring sown. Red number indicates samples shared by the three environments.
2.6.2 青稞育成品种的稳定性评价 为评价育成品种在不同生态环境的稳定性, 选择主推青稞品种藏青320、藏青2000、喜玛拉19号、喜玛拉22号和早期青稞育种的骨干亲本昆仑1号等代表性青稞育成品种进行环境稳定性分析。藏青320和康青3号稳定性最高, 3个环境下稳定在III等级中, 其次是藏青2000、高原早1号、喜玛拉19号、喜玛拉22号、昆仑1号等品种。所有育成品种中喜玛拉9号、白玉紫芒和藏青148三个品种的环境稳定性最差(图9)。
图9
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9青稞主要育成品种在不同播期和环境之间抽穗期的稳定性分析
I: 早抽穗; II: 偏早抽穗; III: 中间型; IV: 偏晚抽穗; V: 晚抽穗。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉萨春播。
Fig. 9Stability analysis of the heading date of the main variety of Tibet barley in different sowing times and environments
I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
3 讨论
国家农作物种质库保存编目的3602份青稞地方品种中, 日喀则、山南、昌都、林芝和拉萨的资源占98%, 阿里和那曲的资源仅占2%[21]。本研究中, 地理来源和基础信息相似的材料只选代表性品种, 共选取1505份地方品种。育成品种包含自1966年开展青稞育种以来, 每个阶段具有一定推广面积和利用时间的代表性品种。用于本研究的实验材料不仅覆盖西藏所有生态区, 四省区藏区的育成品种也选自曾经或者近年多次用来改良西藏青稞的骨干品种, 能够全面反映青稞种质资源的整体情况。鉴定结果表明, 青稞地方品种绝大多数是春性类型, 这与青稞种植实际相吻合。在西藏, 生产上一直以春青稞为主, 冬青稞只在边境地区和低海拔极小面积的盆地种植, 年总面积只有267~333 hm2 [17,18]。近几年随着西藏种植业结构调整和冬青稞复种饲草的巨大优势, 冬青稞面积逐渐扩大, 年播种面积稳定在3333 hm2以上, 但是现有的冬青稞品种很难在海拔3700 m以上的高寒农区推广[21]。一般情况下,品种冬性与苗期抗寒性正相关[8]。本研究发现生产上主推的冬青稞育成品种均为半冬性类型, 这是限制现有冬青稞品种在高海拔地区大面积推广的主要因素之一。除西藏昌都和林芝有较少的冬青稞资源外, 在其他生态区冬青稞资源极为稀少。因此, 除有效利用西藏当地的强冬性资源外, 引进优异抗寒冬大麦种质以丰富和拓宽冬青稞遗传基础是未来冬青稞育种的有效途径。
本研究发现, 冬春性是影响青稞抽穗期的最主要因素, 在所有环境中, 地方品种和育成品种的冬性类型成熟均明显晚于春性类型(图4)。藏东南的生态区普遍存在春性品种秋播的现象, 这些区域的资源也往往当作冬性类型, 这种混淆大幅降低了种质资源的利用效率。因此, 冬春性的精准鉴定是有效利用青稞资源的前提。本研究鉴定的冬春性与马得泉[21]的研究结果基本一致, 但由于20世纪80年代青稞资源收集入库编目时以种植区域和种植习惯作为判断冬春性的主要依据, 即秋播方式的春性材料记为冬性, 因此, 本研究也是对青稞资源目录性状的补充鉴定。在所有环境中, 冬性育成品种和冬性地方品种之间抽穗期无明显差异, 且综合农艺性状与冬青稞骨干亲本“果洛”类似, 可能与冬青稞地方品种数目相对少且均源于西藏东南部有关, 但是对于这些冬青稞种质资源的遗传相似性有待进一步研究。
根据参试品种在不同播期或环境条件下抽穗期的变化规律, 有些品种在环境之间的波动较小, 而有些却很大。对于波动较小的品种, 环境的改变不会引起表型上大的变化, 其说这些品种对环境的改变不太敏感, 它们的环境稳定性较高。而对于波动较大的品种, 环境的改变会引起表型上较大的变化, 它们对环境的改变比较敏感, 其环境稳定性较低。因此, 根据多个品种在多种环境下的表型数据, 就能对品种的环境敏感性进行评价, 又称为稳定性分析(stability analysis)[30]。本研究通过抽穗期的标准差倍数对资源分类, 共分I~V 5个等级, 发现绝大多数地方品种和育成品种分布在偏早抽穗类型(II)、中间型(III)和偏晚抽穗类型中(IV) (图8)。品种稳定性分析发现藏青320在西藏育成品种中稳定性最高(图9), 在青稞生产实践中藏青320具有广适应性和稳定性而种植了近40年。林芝波密县白玉村的紫芒, 因当地表现良好而被选为骨干亲本, 早期推广到其他生态区, 配置了大量杂交组合。但是, 在其他地区并没有达到预期的效果, 未能成为优势品种。另外, 90年代审定的藏青148示范数年后因综合表现不稳定而未能大面积推广。本研究品种稳定性分析也显示, 白玉紫芒和藏青148稳定最差(图9)。因此, 品种稳定性能够较为准确地反映品种在实际生产中的推广情况和利用潜力。
作物品种的抽穗期是最重要的农艺性状之一, 基于抽穗期的稳定性与育成品种的生长情况完全吻合, 即稳定性与推广面积和年限正相关。因此, 广适应性青稞品种的选育应注重参考品种抽穗期的稳定性。I~V 5个等级地方品种中均存在抽穗期稳定性高的材料, 来源青海的昆仑1号是早期(1974—1986年)西藏青稞育种的一个骨干亲本[31]。本研究中来源于四川甘孜的康青3号是所有参试材料中稳定性最好的品种, 其稳定性显著高于昆仑1号, 且综合性状优良, 可作为今后青稞改良的候选骨干亲本。
4 结论
生长习性和抽穗期是决定青稞生态适应性的重要因素。青稞资源的抽穗期具有丰富的变异, 多样性分析和不同环境稳定性评价为高稳定性的青稞种的筛选及高产、广适应性春冬青稞品种的选育提供了理论依据。致谢:
感谢中国农业科学院作物科学研究所小宗作物种质资源创新研究者张京研究员提供青稞种质, 数量遗传创新研究组王建康研究员指导数据分析。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1093/aob/mcm139URL [本文引用: 1]
DOI:10.1126/science.1117619URL [本文引用: 1]
DOI:10.1093/molbev/msn167URL [本文引用: 1]
DOI:10.1371/journal.pone.0200722URL [本文引用: 1]
DOI:10.1016/j.cell.2006.12.006URL [本文引用: 1]
DOI:10.1104/pp.108.128827URL [本文引用: 1]
DOI:10.1073/pnas.1635053100URL [本文引用: 1]
DOI:10.1093/pcp/pcr046URL [本文引用: 2]

In many temperate plant species, prolonged cold treatment, known as vernalization, is one of the most critical steps in the transition from the vegetative to the reproductive stage. In contrast to recent advances in understanding the molecular basis of vernalization in Arabidopsis non-vernalization mutants or the spring growth habits of cereal crops such as wheat and barley, natural variations in winter growth habits and their geographic distribution are poorly understood. We analyzed varietal variation and the geographic distribution of the degree of vernalization requirements in germplasms of domesticated barley and wild barley collections. We found a biased geographic distribution of vernalization requirements in domesticated barley: Western regions were strongly associated with a higher degree of spring growth habits, and the extreme winter growth habits were localized to Far Eastern regions including China, Korea and Japan. Both wild accessions and domesticated landraces, the regions of distribution of which overlapped each other, mainly belonged to the moderate class of winter growth habit. As a result of quantitative evaluations performed in this study, we provide evidence that the variation in the degree of winter growth habit in recombinant inbred lines was controlled by quantitative trait loci including three vernalization genes (VRN1, VRN2 and VRN3) that account for 37.9% of the variation in vernalization requirements, with unknown gene(s) explaining the remaining two-thirds of the variation. This evidence implied that the Far Eastern accessions might be a genetically differentiated group derived for an evolutionary reason, resulting in their greater tendency towards a winter growth habit.
DOI:10.1007/s11032-012-9765-0URL [本文引用: 1]
DOI:10.1038/ng.2447URL [本文引用: 1]
DOI:10.1139/g95-074URL [本文引用: 1]
DOI:10.1073/pnas.0937399100URL [本文引用: 1]
DOI:10.1126/science.1094305URL [本文引用: 1]
DOI:10.1073/pnas.0607142103URL [本文引用: 1]
DOI:10.1534/genetics.106.069500URL [本文引用: 1]
DOI:10.1016/j.pbi.2008.12.010URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 5]
[本文引用: 5]
URL [本文引用: 1]
将我国大麦基因库划分成主基因库、二级基因库和三级基因库等不同的结构类群,分析了主基因库18839份种质资源共26个表型性状的多样性及其地理分布。总体上讲,国内大麦的多样性高于国外大麦,从野生大麦经农家到现代育成品种,多样性逐渐降低。库内各省区的大麦种质资源保存份数与多样性大小相关不显著。多样性的地区分布不均匀,主要集中在云贵、青藏高原和四川、黄河流域和长江中下游地区,表现特点与分布区生态密切相关。
URL [本文引用: 1]
将我国大麦基因库划分成主基因库、二级基因库和三级基因库等不同的结构类群,分析了主基因库18839份种质资源共26个表型性状的多样性及其地理分布。总体上讲,国内大麦的多样性高于国外大麦,从野生大麦经农家到现代育成品种,多样性逐渐降低。库内各省区的大麦种质资源保存份数与多样性大小相关不显著。多样性的地区分布不均匀,主要集中在云贵、青藏高原和四川、黄河流域和长江中下游地区,表现特点与分布区生态密切相关。
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1371/journal.pone.0113120URL [本文引用: 1]
DOI:10.3389/fpls.2017.00896URL [本文引用: 1]
DOI:10.1007/s00438-009-0449-3URL [本文引用: 1]
DOI:10.1007/s11032-016-0434-6URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}