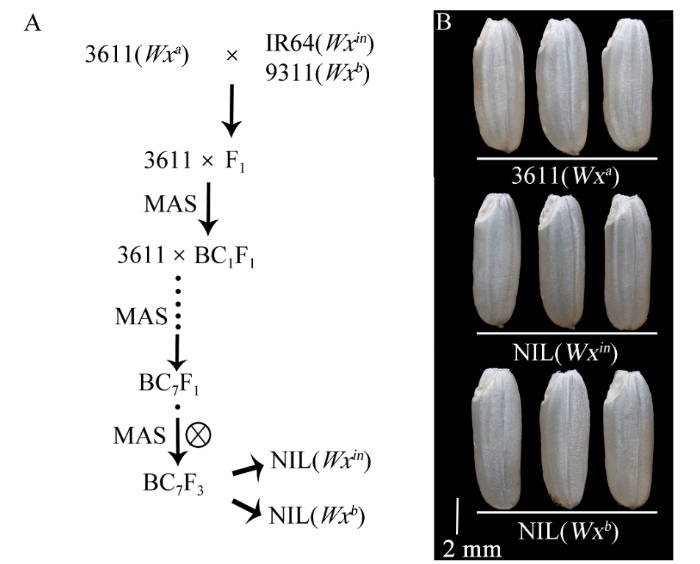
Improvement of rice eating quality and physicochemical properties by introgression of Wx in allele in indica varieties
YANG Yong1, LU Yan1,2, GUO Shu-Qing1, SHI Zhong-Hui1, ZHAO Jie1, FAN Xiao-Lei1, LI Qian-Feng1, LIU Qiao-Quan,1,*, ZHANG Chang-Quan,1,*1 Jiangsu Key Laboratory for Crop Genomics and Molecular Breeding / Key Laboratory of Plant Functional Genomics of Ministry of Education / Co-Innovation Center for Modern Production Technology of Grain Crops, Agricultural College of Yangzhou University, Yangzhou 225009, Jiangsu, China 2 Instrumental Analysis Center, Yangzhou University, Yangzhou 225009, Jiangsu, China
This study was supported by the National Key Research and Development Program of China.2016YFD0100501 the National Natural Science Foundation of China.31872860 the National Natural Science Foundation of China.31561143008 the Government of Jiangsu Province.BE2018357 the Government of Jiangsu Province.BK20160464 the Natural Science Foundation of the Jiangsu Higher Education Institutions of China.16KJB210011 the Open Research Fund of State Key Laboratory of Hybrid Rice(Hunan Hybrid Rice Research Center).2018KF04Hunan Hybrid Rice Research Center the Personnel Training Program for Undergraduates in Agricultural College of Yangzhou University
Abstract Nowadays, the Wx b allele has been widely used to improve grain quality of indica rice. However, some indica varieties carrying Wx b allele usually has a much softer texture, which is not favored by consumers in South China. So the grain quality of these varieties needs to be further improved. To understand the effect of Wx in allele on rice eating quality and physicochemical properties in indica rice, we developed two Near-Isogenic Lines (NILs) carrying Wx in and Wx b alleles by crossing an indica variety 3611 (receptor, carrying Wx a) with IR64 (carrying Wx in) and 9311 (carrying Wx b), and seven times of backcrossing based on molecular marker assistant selection (MAS). The Wx effects in controlling the synthesis of amylose, grain quality, and physicochemical properties were investigated. There were non-significant differences in the agronomic traits among the NILs. However, for grain quality characters, we found that the NIL(Wx in) rice showed significantly lower apparent amylose content (AAC) and higher gel consistency (GC), compared with the wild type 3611. Besides, the NIL(Wx b) rice showed the lowest AAC and highest GC among three lines. The NIL(Wx in) rice had a significantly higher taste value than the wild type 3611, while the NIL(Wx b) rice exhibited the highest taste value among the three samples. The granule-bound starch synthase I (GBSSI) level was the highest in 3611, moderate in NIL(Wx in) and lowest in NIL(Wx b), which showed a positive correlation with the AAC level. Also, the starch viscosity, thermal gelatinization property and crystal structure of different rice flours had a high correlation with the AAC level. To sum up, our results proved that both Wx in and Wx b allele can improve the grain quality in 3611 background, and what is more, the Wx in allele might be more useful for the improvement of grain quality in indica rice. Keywords:Oryza sativa L.;eating quality;Wx allele;apparent amylose content;molecular marker assisted selection
PDF (2760KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 杨勇, 陆彦, 郭淑青, 石仲慧, 赵杰, 范晓磊, 李钱峰, 刘巧泉, 张昌泉. 籼稻背景下导入Wx in等位基因改良稻米食味和理化品质 [J]. 作物学报, 2019, 45(11): 1628-1637. doi:10.3724/SP.J.1006.2019.82064 YANG Yong, LU Yan, GUO Shu-Qing, SHI Zhong-Hui, ZHAO Jie, FAN Xiao-Lei, LI Qian-Feng, LIU Qiao-Quan, ZHANG Chang-Quan. Improvement of rice eating quality and physicochemical properties by introgression of Wx in allele in indica varieties [J]. Acta Crops Sinica, 2019, 45(11): 1628-1637. doi:10.3724/SP.J.1006.2019.82064
A图为利用QRM190分子标记区分Wxa和Wxb的PCR检测结果, 泳道1为3611, 2为9311, 3~7为近等基因系NIL(Wxb); B图为利用基因特异分子标记区分Wxa和Wxin的PCR检测结果, 泳道1为3611, 2为IR64, 3~7为近等基因系NIL(Wxin)。 Fig. 2Detection of specific molecular markers for different Wx alleles
(A): allelic specific molecular marker QRM190 for detecting Wxa and Wxb. Lanes 1-7: 3611, 9311 and their derived NIL NIL(Wxb) (lanes 3-7), respectively. (B): allelic specific molecular marker for detecting Wxa and Wxin. Lanes 1-7: 3611, IR64 and their derived NIL NIL(Wxin) (lanes 3-7), respectively.
M: 蛋白质分子质量标准; 1~2: 3611(Wxa); 3~4: NIL(Wxin); 5~6: NIL(Wxb); 箭头所示为60 kD的GBSSI蛋白。 Fig. 3SDS-PAGE analysis of the soluble (A) and bound (B) GBSS I isolated from mature rice of different NILs
Lane M: the protein standard molecular mass; Lanes 1-2: 3611(Wxa); Lane 3-4: NIL(Wxin); Lanes 5-6: NIL(Wxb); The arrows indicate the 60 kD GBSSI.
Table 2 表2 表2不同近等基因系稻米理化和外观品质 Table 2Appearance quality and physicochemical properties of different near-isogenic lines
品系 Line
表观直链淀粉含量 Apparent amylose content (%)
胶稠度 Gel consistency (mm)
食味值 Taste value
垩白粒率 Chalkiness rate (%)
垩白度 Chalkness degree (%)
3611(Wxa)
25.51±1.48 a
42.35±4.17 c
43.15±1.35 c
18.62±0.87 a
2.89±1.05 a
NIL(Wxin)
19.85±0.27 b
78.54±2.32 b
55.09±2.51 b
13.04±0.30 b
1.93±0.06 c
NIL(Wxb)
14.78±0.55 c
87.62±3.48 a
60.51±1.87 a
13.05±0.40 b
2.03±0.32 b
Values within the same column followed by different letters are significant different at P < 0.01 and n = 3. 同列标以不同小写字母的值差异极显著(P < 0.01), n = 3。
TilmanD, BalzerC, HillJ, Befort BL . Global food demand and the sustainable intensification of agriculture Proc Natl Acad Sci USA, 2011,108:20260-20264. [本文引用: 1]
Zhang CQ, Zhao DS, Li QF, Gu MH, Liu QQ . Progresses in research on cloning and functional analysis of key genes involving in rice grain quality Sci Agric Sin, 2016,49:4267-4283 (in Chinese with English abstract). [本文引用: 2]
Zeng DL, Tian ZX, Rao YC, Dong GJ, Yang YL, Huang LC, Leng YJ, XuJ, SunC, Zhang GH, HuJ, ZhuL, Gao ZY, Hu XM, Guo LB, Xiong GS, Wang YH, Li JY, QianQ . Rational design of high-yield and superior-quality rice Nat Plants, 2017,3:17031. [本文引用: 4]
Li HY, PrakashS, Nicholson TM, Fitzgerald MA, Gilbert RG . The importance of amylose and amylopectin fine structure for textural properties of cooked rice grains Food Chem, 2016,196:702-711. [本文引用: 2]
Tian ZX, QianQ, Liu QQ, Yan MX, Liu XF, Yan CJ, Liu GF, Gao ZY, Tang SH, Zeng DL, Wang YH, Yu JM, Gu MH, Li JY . Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities Proc Natl Acad Sci USA, 2009,106:21760-21765. [本文引用: 2]
MohapatraD, BalS . Cooking quality and instrumental textural attributes of cooked rice for different milling fractions J Food Eng, 2006,73:253-259. [本文引用: 1]
He XP, Zhu CL, Liu LL, JiangL, Zhang WW, Liu YB, Wan JM . Difference of amylopectin structure among various rice genotypes differing in grain qualities and tts relation to starch physicochemical properties Acta Agron Sin, 2010,36:276-284 (in Chinese with English abstract). [本文引用: 1]
Li HY, Gilbert RG . Starch molecular structure: The basis for an improved understanding of cooked rice texture Carbohyd Polym, 2018,195:9-17. [本文引用: 1]
Tao KY, LiC, Yu WW, Gilbert RG, Li EP . How amylose molecular fine structure of rice starch affects functional properties Carbohyd Polym, 2019,204:24-31. [本文引用: 1]
Zhu JH, Zhang CQ, Gu MH, Liu QQ . Progress in the allelic variation of Wx gene and its application in rice breeding. Chin J Rice Sci, 2015,29:431-438 (in Chinese with English abstract). [本文引用: 1]
Cai XL, Wang ZY, Xing YY, Zhang JL, Hong MM . Aberrant splicing of intron 1 leads to the heterogeneous 5' UTR and decreased expression of waxy gene in rice cultivars of intermediate amylose content. Plant J, 2010,14:459-465. [本文引用: 1]
IsshikiM, MorinoK, NakajimaM, Okagaki RJ, Wessler SR, IzawaT, ShimamotoK . A naturally occurring functional allele of the rice waxy locus has a GT to TT mutation at the 5' splice site of the first intron. Plant J, 1998,15:133-138. [本文引用: 1]
SreenivasuluN, ButardoV M J, MisraG, CuevasR P, AnacletoR, Kavi KishorP B . Designing climate-resilient rice with ideal grain quality suited for high-temperature stress J Exp Bot, 2015,66:1737-1748. [本文引用: 2]
MikamiI, UwatokoN, IkedaY, YamaguchiJ, Hirano HY, SuzukiY, SanoY . Allelic diversification at the wx locus in landraces of Asian rice. Theor Appl Genet, 2008,116:979-989. [本文引用: 2]
Xiang XC, Kang CF, Xu SJ, Yang BW . Combined effects of Wx and SSIIa haplotypes on rice starch physicochemical properties. J Sci Food Agric, 2017,97:1229-1234. [本文引用: 3]
Yang BW, Xu SJ, XuL, YouH, Xiang XC . Effects of Wx and its interaction with SSIII-2 on rice eating and cooking qualities. Front Plant Sci, 2018,9:456. [本文引用: 1]
UmemotoT, HoribataT, AokiN, HiratsukaM, YanoM, InouchiN . Effects of variations in starch synthase on starch properties and eating quality of rice Plant Prod Sci, 2008,11:472-480. [本文引用: 1]
Cao XM, Sun HY, Wang CG, Ren XJ, Liu HF, Zhang ZJ . Effects of late-stage nitrogen fertilizer application on the starch structure and cooking quality of rice J Sci Food Agric, 2017,98:2332-2340. [本文引用: 1]
Tian ZX, Yan CJ, QianQ, YanS, Xie HL, WangF, Xu JF, Liu GF, Wang YH, Liu QQ, Tang SZ, Li JY, Gu MH . Development of gene-tagged molecular markers for starch synthesis-related genes in rice Chin Sci Bull, 2010,55:3768-3777. [本文引用: 1]
Murray MG, Thompson WF . Rapid isolation of high molecular weight plant DNA Nucl Acids Res, 1980,8:4321-4325. [本文引用: 1]
Liu QQ, Zhang JL, Wang ZM, Hong MM, Gu MH . A highly efficient transformation system mediated by Agrobacterium tumefaciens in rice(Oryza sativa L.). Acta Phytophysiol Sin, 1998,24:259-271 (in Chinese with English abstract). [本文引用: 1]
Zhang CQ, Zhu LJ, ShaoK, Gu MM, Liu QQ . Toward underlying reasons for rice starches having low viscosity and high amylose: physiochemical and structural characteristics J Sci Food Agric, 2013,93:1543-1551. [本文引用: 3]
Liu DR, WangW, Cai XL . Modulation of amylose content by structure-based modification of OsGBSS1 activity in rice (Oryza sativa L.). Plant Biotechnol J, 2015,12:1297-1307. [本文引用: 1]
Shu QY, Wu DX, Xia YW, Gao MW, Anna MC . Relationship between RVA profile characteristics of rice starch and edible quality Sci Agric Sin, 1998,31:25-29 (in Chinese with English abstract). [本文引用: 2]
Zhang CQ, Chen SJ, Ren XY, LuY, Liu DR, Cai XL, Li QF, Gao JP, Liu QQ . Molecular structure and physicochemical properties of starches from rice with different amylose contents resulting from modification of OsGBSSI activity J Agric Food Chem, 2017,65:2222. [本文引用: 4]
Cai JW, Man JM, HuangJ, Liu QQ, Wei WX, Wei CX . Relationship between structure and functional properties of normal rice starches with different amylose contents Carbohyd Polym, 2015,125:35-44. [本文引用: 2]
CookeD, Gidley MJ . Loss of crystalline and molecular order during starch gelatinisation: origin of the enthalpic transition Carbohyd Polym, 1992,227:103-112. [本文引用: 1]
SevenouO, Hill SE, Farhat IA, Mitchell JR . Organisation of the external region of the starch granule as determined by infrared spectroscopy Int J Biol Macromol, 2002,31:79-85. [本文引用: 1]
Chen MJ, Liu GF, YuH, WangB, Li JY . Towards molecular design of rice plant architecture and grain quality Chin Sci Bull, 2018,63:1276-1289. [本文引用: 1]
Chen MH, Bergman CJ, PinsonaS R M, FjellstromR G . Waxy gene haplotypes: associations with pasting properties in an international rice germplasm collection J Cereal Sci, 2008,48:781-788. [本文引用: 2]
HoaiT T T, MatsusakaH, ToyosawaY, SuuT D, SatohH, KumamaruT . Influence of single-nucleotide polymorphisms in the gene encoding granule-bound starch synthase I on amylose content in Vietnamese rice cultivars Breed Sci, 2014, 64:142. [本文引用: 2]
Luo JX, Jobling SA, MillarA, Morell MK, Li ZY . Allelic effects on starch structure and properties of six starch biosynthetic genes in a rice recombinant inbred line population Rice, 2015,8:15. [本文引用: 2]
Li QF, Liu XY, Zhang CQ, JiangL, Jiang MY, ZhongM, Fan XL, Gu MH, Liu QQ . Rice soluble starch synthase: I. Allelic variation, expression, function, and interaction with Waxy Front Plant Sci, 2018,9:1591. [本文引用: 1]
WangK, HasjimJ, Wu AC, Li EP, Henry RJ, Gilbert RG . Roles of GBSSI and SSIIa in determining amylose fine structure Carbohydr Polym, 2015,127:264-74. [本文引用: 1]
Zhou HJ, Wang LJ, Liu GF, Meng XB, Jing YH, Shu XL, Kong XL, Sun JA, YuH, Smith SM, Wu DX, Li JY . Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice Proc Natl Acad Sci USA, 2016,113:12844-12849. [本文引用: 1]
Fan MY, Wang XJ, SunJ, ZhangQ, Xu ZJ, XuQ . Effect of indica pedigree on eating and cooking quality in rice backcross inbred lines of indica and japonica crosses. Breed Sci, 2017,67:450-458. [本文引用: 1]
MisraG, BadoniS, Domingo CJ, CuevasR P O, LlorenteC, MbanjoE G N, SreenivasuluN . Deciphering the genetic architecture of cooked rice texture Front Plant Sci, 2018,9:1405. [本文引用: 1]
TengB, Zeng RZ, Wang YC, Liu ZQ, Zhang ZM, Zhu HT, Ding XH, Li WT, Zhang GQ . Detection of allelic variation at the Wx locus with single-segment substitution lines in rice(Oryza sativa L.) Mol Breed, 2012,30:583-595. [本文引用: 2]
TengB, ZhangY, Du SY, Wu JD, Li ZF, Luo ZH, Yang JB . Crystalline, thermal and swelling properties of starches from single-segment substitution lines with different Wx alleles in rice(Oryza sativa L.). J Sci Food Agric, 2017,97:108-114. [本文引用: 2]
Bao JS, Kong XG, Xie, J K, XuL J Analysis of genotypic and environmental effects on rice starch: 1. Apparent amylose content, pasting viscosity, and gel texture J Agric Food Chem, 2004,52:6010-6016. [本文引用: 1]
Champagne ET, Bett-GarberK L, FitzgeraldM A, GrimmC C, LeaJ, OhtsuboK I, JongdeeS, XieL H, BassinelloP Z, ResurreccionA, AhmadR, HabibiF, ReinkeR . Important sensory properties differentiating premium rice varieties Rice, 2010,3:270-281. [本文引用: 1]
InukaiT, HirayamaY . Comparison of starch levels reduced by high temperature during ripening in japonica rice lines near- isogenic for the Wx locus. J Agron Crop Sci, 2010,196:296-301. [本文引用: 1]
WangK, Zhou QF, Liu JY, Qiu FL, Angelitadela Paz M, LarazoW, YangY Z, XieF M . Genetic effects of Wx allele combinations on apparent amylose content in tropical hybrid rice Cereal Chem, 2017,94:887-891. [本文引用: 1]
 ,1,*, 张昌泉
,1,*, 张昌泉
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT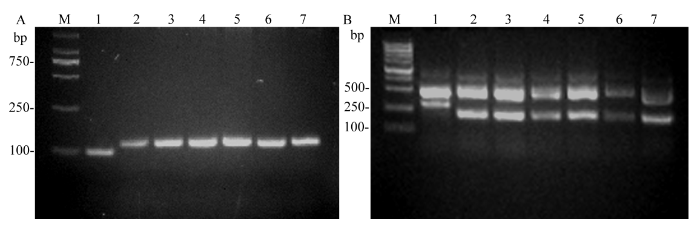 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT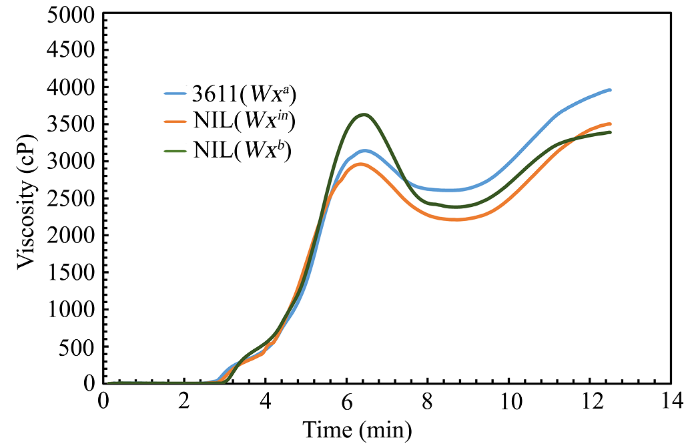 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT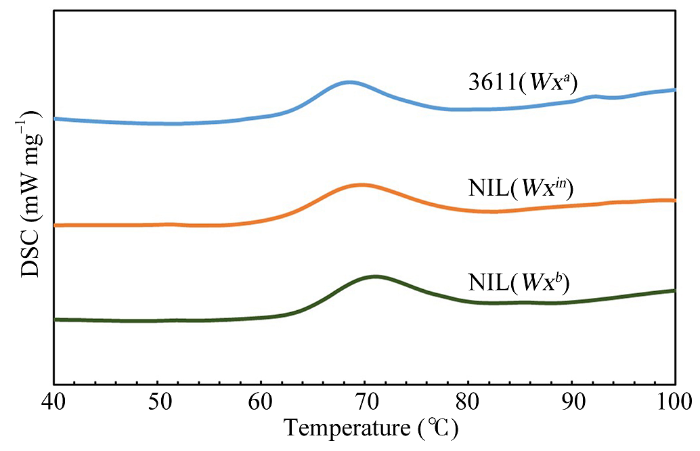 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT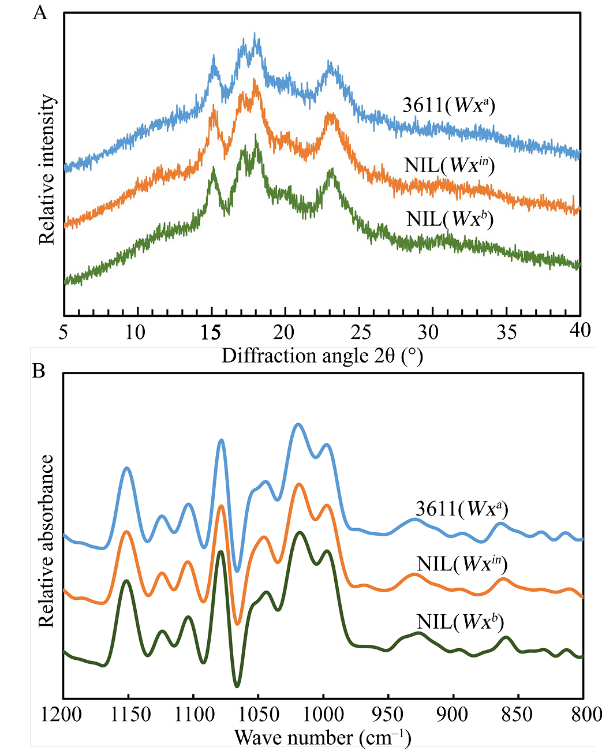 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}