 ,*中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程/农业部麦类生物学与遗传育种重点实验室, 北京 100081
,*中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程/农业部麦类生物学与遗传育种重点实验室, 北京 100081Cloning and functional analysis of wheat receptor-like kinase gene TaPK3A
SU Qiang, RONG Wei, ZHANG Zeng-Yan,*National Key Facility for Crop Gene Resources and Genetic Improvement/Key Laboratory of Crop Genetic and Breeding, Ministry of Agriculture/Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China通讯作者:
收稿日期:2018-09-30接受日期:2019-01-19网络出版日期:2019-05-09
| 基金资助: |
Received:2018-09-30Accepted:2019-01-19Online:2019-05-09
| Fund supported: |
作者简介 About authors
E-mail:243939427@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (1411KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
苏强, 荣玮, 张增艳. 小麦类受体蛋白激酶基因TaPK3A的克隆与抗纹枯病功能初步分析[J]. 作物学报, 2019, 45(8): 1158-1165. doi:10.3724/SP.J.1006.2019.81017
SU Qiang, RONG Wei, ZHANG Zeng-Yan.
小麦纹枯病是由禾谷丝核菌(Rhizoctonia cerealis)侵染引起的一种土传真菌病害, 在世界范围内皆有发生, 近年在我国发病区域和危害程度呈上升趋势, 已成为我国小麦生产中的主要病害之一。2012—2018年间我国病虫害测报处(中国农业信息网, http://www.agri.cn/)的数据显示, 小麦纹枯病发病面积为我国总病害发病面积的23.0%~32.5%, 一般可减产10%左右, 严重时可减产40%以上[1]。虽然已报道过一些小麦抗病资源[2-3], 但是它们农艺性状差难以被直接应用, 加之有效抗病基因与分子标记的匮乏, 严重制约了小麦抗纹枯病品种改良的进展。因此,从小麦基因组中筛选有效的抗纹枯病基因对小麦抗纹枯病分子育种和传统育种具有重要意义。
蛋白激酶识别病原效应因子或传递下游信号参与防御反应[4]。植物类受体蛋白激酶是跨膜蛋白, 由胞外的受体识别结构域、跨膜结构域和位于细胞质的激酶结构域三部分组成。类受体蛋白激酶(receptor-like protein kinase, RLK)通过胞外结构域感知刺激, 并通过其胞质激酶结构域传递信号[5,6,7]。根据胞外结构域的不同, 植物RLK可分为富含亮氨酸重复序列的受体激酶(leucine-rich repeat, LRR)、细胞壁相关的类受体激酶(wall-associated kinase, WAK/WAK-like kinase, WAKL)、含赖氨酸基序的受体激酶(lysine motif, Lys M)、类凝集素结合域的受体激酶(lectin-binding domain, LB)、具有S-结构域的受体激酶(self-incompatibility locus, S-locus)等[7,8]。植物RLK数量众多, 属于超大家族。如在拟南芥(Arabidopsis thaliana)中已鉴定出600多个受体蛋白激酶[6,10]。类受体蛋白激酶广泛存在于植物中[9,10,11]。RLK功能包括激素响应[12]、发育控制[13]、生殖调节[14]、结瘤作用[15,16]和抗病反应[17,18]。例如, 水稻Xa26基因编码一个RLK蛋白, 具有抗白叶枯病作用[19]。在拟南芥中, SA处理提高AtWAK1的转录丰度, 过表达AtWAK1增强了拟南芥对灰霉病的抗性[20]。当稻瘟病菌(Magnaporthe oryzae)侵染后, OsWAK1的转录水平受到显著诱导, 且OsWAK1过表达的转基因水稻对稻瘟病小种P007的抗性显著增强[21]。玉米ZmWAK基因可以感知丝黑穗病菌(Sporisorium reilianum)的入侵, 触发宿主防御反应, 进而抑制病原菌的生长[22]。外源凝集素受体激酶基因LecRK-VI.2可提高拟南芥对丁香假单胞菌(Pseudomonas syringae)的抵御能力[23]。水稻中, Pi-d2赋予寄主植株对稻瘟菌生理小种ZB15的抗性[24]。当水稻携带3个凝集素受体激酶基因OsLecRK1、OsLecRK2、OsLecRK3时, 具有对褐飞虱的广谱抗性[25]。具有S-结构域受体激酶基因LRK10在小麦抗叶锈病反应中起重要作用[26]。TaRLK-R1、TaRLK-R2、TaRLK-R3是小麦3个同源RLK基因, 不仅响应胁迫, 而且正向参与小麦抗条锈病的超敏反应[27]。
本研究利用小麦转录组数据发掘出一个类受体激酶类基因TaPK3A, 并从抗纹枯病小麦品系CI12633中克隆了TaPK3A基因。对TaPK3A在抗纹枯病小麦、感纹枯病小麦中的表达谱以及小麦不同组织的表达特性进行了分析, 也进行了TaPK3A响应激素的研究。重要的是利用BSMV-VIGS沉默技术分析TaPK3A对小麦纹枯病的防御功能, 旨在进一步了解小麦抗纹枯病反应的分子基础, 为小麦抗纹枯病分子育种提供候选基因。
1 材料与方法
1.1 试验材料
小麦品种温麦6号和品系CI12633, 分别由中国农业科学院作物科学研究所李立会研究员和江苏省农业科学院粮食作物研究所蔡士宾研究员惠赠。由山红麦和温麦6号衍生的RIL (recombinant inbred line)群体由中国农业科学院作物科学研究所贾继增研究员惠赠。禾谷丝核菌(Rhizoctonia cerealis)北方强毒力菌株WK207由山东农业大学于金凤教授惠赠。1.2 TaPK3A基因的分离及序列分析
对RIL系植株(F11)进行抗纹枯病鉴定, 从中选出抗、感纯合株系各3个, 于苗期接种禾谷丝核菌WK207。于接种前(0 h)、接种后4 d和10 d分别取样, 抗病系样品分别记为R0、R4和R10, 感病系样品分别记为S0、S4和S10, 首先用TRIzol Reagent提取试剂(北京百奥科利科技有限公司)提取小麦总RNA, 然后用FastKing RT Kit (含gDNase, KR116, 天根生化科技有限公司)合成第1链cDNA。分别以R0/S0、R4/S4和R10/S10的转录组数据分析小麦抗、感株系间的表达差异基因, 根据注释信息在Ensembl Plants数据库(http://plants.ensembl.org/ index.html)中得到候选基因Traes_3AS_928B35848的EST序列。使用Primer Premier 5.0软件设计2对引物, 其序列为TaPK3A-F1: 5′-TACCAGCTTCTATGCCT G-3′, TaPK3A-R1: 5′-ACACTCTCAAACCTTCCA-3′; TaPK3A-F2: 5′-GCATCTGTAAAGCTCTCATG-3′, Ta PK3A-R2: 5′-CACCTGAGGTGGGTAGAG-3′。利用TaKaRa LA Taq酶(DRR002A), 对该基因进行巢式PCR扩增。第1轮扩增程序为94°C 3 min; 30×(94°C 30 s, 50°C 30 s, 72°C 140 s); 72°C 10 min; 4°C保存。将第1轮扩增产物稀释50倍作为第2轮扩增的模板。第2轮扩增程序为94°C 3 min; 35×(94°C 30 s, 50°C 30 s, 72°C 120 s); 72°C 10 min; 4°C保存。扩增产物经1.0%琼脂糖凝胶电泳分离, 切胶回收, 并连接到pMD-18T(D101A)载体进行测序, 获得小麦基因TaPK3A序列。用Protparam (http://web.expasy.org/ protparam)分析TaPK3A编码氨基酸的理化性质, 用SignalP-4.1 Server在线工具(http://www.cbs.dtu.dk/ services/SignalP/)分析其氨基酸序列中的信号肽信息, 用SMART (http://smart.embl-heidelberg.de/)预测蛋白结构域, 使用MEGA7软件对TaPK3A及其同源序列进行进化树分析。
1.3 TaPK3A基因表达分析
1.3.1 TaPK3A在抗纹枯病小麦CI12633中的表达分析 CI12633拔节期, 用牙签法接种禾谷丝核菌WK207, 于未接种(mock)和接种后2、4、7、10、14和21 d分别取接种部位样品, 按上述方法提取总RNA, 反转录合成第1链cDNA。采用ABI PRISMR 7500实时荧光定量PCR仪进行实时荧光定量RT-PCR分析。用Primer Premier 5.0设计TaPK3A特异性引物(TaPK3A-RT-F: 5'-TCTTCTGCGGGGACGAGTA-3′; TaPK3A-RT-R: 5′-CCTGAGGTGGGTAGAGGATAAA- 3′), 以TaActin (GenBank登录号为MF405765.1, TaActin-F: 5′-AACCGGAATGGTCAAGGCTG-3′; Ta Actin-R: 5′-CTCCATGTCATCCCAGTTG-3′)为内参基因, 按试剂盒说明书设计PCR反应体系, 含SYBR Premix Ex Taq II (Tli RNaseH Plus, TaKaRa) (2×) 12.5 μL, 上、下游引物(10 μmol L-1)各0.80 μL, cDNA模板5.0 μL、ROX Reference Dye II (50×) 0.40 μL, 补RNase-free ddH2O 至20 μL。扩增程序为95°C 30 s; 40×(95°C 5 s, 58°C 15 s, 72°C 34 s); 72°C 10 min。每个样品3次技术重复, 以mock为对照, 按Livak和Schmittgen描述的方法[28]计算TaPK3A基因的相对表达量。用Microsoft Excel软件进行t检验, P<0.05为显著差异, P<0.01为极显著差异。1.3.2 TaPK3A组织表达分析 于CI12633抽穗期, 采用牙签法接种禾谷丝核菌WK207。取接种前(0 d)和接种后(4 d)的根、茎、叶、穗样品分别提取RNA, 进行RT-qPCR分析。
1.3.3 TaPK3A对不同激素处理响应的定量表达分析 使用0.7% H2O2过夜浸泡CI12633种子, 在22°C、16 h光照/8 h黑暗条件下生长至三叶一心期, 喷施外源水杨酸(SA, 1 mmol L-1)、脱落酸(ABA, 0.2 mmol L-1)、茉莉酸甲酯(MeJA, 0.1 mmol L-1)、乙烯合成前体(ACC, 0.1 mmol L-1), 在处理前(0 h)和处理后0.5、1、6和24 h取样, 取3个植株的叶片混样后提取总RNA, 进行RT-qPCR分析。
1.4 BSMV-VIGS载体的构建和实施
BSMV病毒载体α、β和γ:GFP由本实验室保存。参照Holzberg等[29]描述的方法, 根据TaPK3A的cDNA序列, 设计特异序列区段引物(TaPK3A- VIGS-F: 5°-GCGCTAGCAAGCTGTGCGTCGTTGC G-3°; TaPK3A-VIGS-1145R: 5°-GCGCTAGCCACCT GAGGTGGGTAGAGGA-3°, 下画线表示Nhe I酶切位点)。利用PCR方法获得含TaPK3A 3°端的特异片段126 bp, 构建重组载体γ:TaPK3A。用限制性内切酶Mlu I (TaKaRa)对α、γ:GFP、γ:TaPK3A载体质粒线性化, 用限制性内切酶Spe I (TaKaRa)对β质粒线性化。以线性化的质粒为模板, 按Zhu等[30]描述的方法, 使用RiboMA Large Scale RNA Production System-T7试剂盒(Promega, USA)体外转录。对三叶一心期的CI12633摩擦接种BSMV, 接种病毒10 d后提取小麦叶片总RNA, 反转录成单链cDNA作为分析的模板。利用半定量PCR对BSMV CP基因的转录表达进行分析, 反应体系为2× PCR StarMix (北京康润诚业有限公司)12.5 μL, TaActin正、反引物(10 μmol L-1)各1.0 μL, 不同材料的cDNA模板量不同, 用RNase-free ddH2O补至25 μL。扩增程序为94°C 3 min; 16× (94°C 30 s, 57°C 30 s, 72°C 30 s); 72°C 10 min。以TaActin电泳亮度调整加入模板的量, BSMV:GFP植株1.0 μL, BSMV: TaPK3A植株分别为0.75、1.5和2.0 μL。利用BSMV-CP引物(F: 5′-TGACTGCTAAGGGTGGAGG A-3′; R: 5′-CGGTTGAACATCACGAAGAGT-3′)进行PCR扩增, 26个循环, 120 V琼脂糖凝胶电泳20 min。以TaActin为内标, 对不同样品模板进行均一化处理, 然后利用TaPK3A特异引物TaPK3A-RT-F/ TaPK3A-RT-R, 对成功接种BSMV重组病毒的小麦进行RT-qPCR分析, 检测TaPK3A基因转录物的积累水平。
1.5 BSMV-VIGS小麦植株的纹枯病鉴定
在小麦拔节期, 采用牙签法对沉默植株和对照植株接种禾谷丝核菌WK207。用湿润的脱脂棉将接种部位包住保湿, 每天喷水3次, 保持相对湿度不低于100%, 持续1周。于接种纹枯病后30 d调查病情, 按5级标准[30,31]划分小麦纹枯病的病级(infection type, IT)和病情指数(disease index, DI), 以评价小麦纹枯病抗性。2 结果与分析
2.1 TaPK3A基因的发掘克隆与序列分析
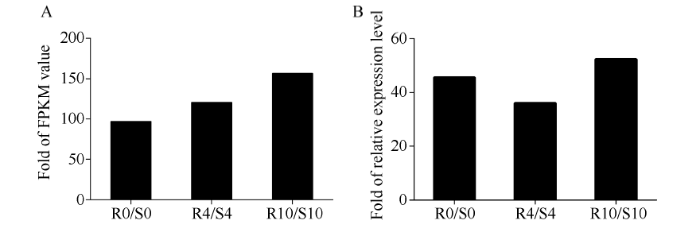
分析小麦转录组数据, 发现一个响应纹枯病侵染的基因Traes_3AS_928B35848。通过巢式PCR方法从小麦CI12633的cDNA中克隆得到TaPK3A全长编码序列(GenBank登录号为MG882067)。TaPK3A序列与Traes_3AS_928B35848的cDNA序列具有93.44%的相似性。在0、4和10 d 时间点, Traes_3AS_928B35848的表达量在抗病株系中比感病株系中上调96.7、120.8和156.6倍(图1-A)。当小麦纹枯病菌侵染时, TaPK3A的表达量上调, 并且与转录组数据趋势一致(图1-B)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1RIL中TaPK3A的表达分析
A: 抗感材料RNA-Seq 数据中TaPK3A的表达量; B: RT-qPCR验证TaPK3A的表达量; R代表RIL中抗纹枯病小麦株系, S代表RIL中感纹枯病株系; R0/S0表示在0 h时R中和S中TaPK3A的表达量的比值; R4/S4表示在4 d时R中和S中TaPK3A的表达量的比值; R10/S10表示在10 d时R中和S中TaPK3A的表达量的比值。
Fig. 1Transcriptional analyses of TaPK3A in RIL
A: expression of TaPK3A in RNA-Seq data of resistant and susceptible materials. B: RT-qPCR verification of the transcription of TaPK3A. R stands for sharp eyespot resistant wheat lines in RIL. S stands for sharp eyespot susceptible wheat lines in RIL. R0/S0 stands for the ratio of TaPK3A expression in R and S at 0 h. R4/S4 stands for the ratio of TaPK3A expression in R and S at 4 d. R10/S10 stands for the ratio of TaPK3A expression in R and S at 10 d.
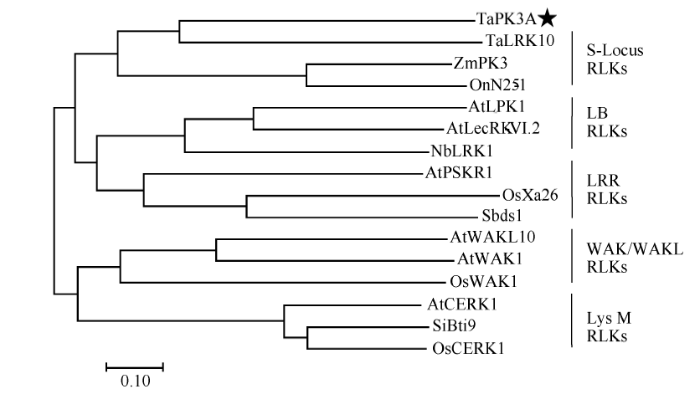
序列分析显示, TaPK3A有一个长度为1983 bp的开放阅读框, 编码660个氨基酸组成的类受体蛋白激酶TaPK3A。预测TaPK3A的分子量为73.95 kDa, 等电点是5.95。SMART软件分析表明, TaPK3A具有一个信号肽(第1~第31位氨基酸), 一个跨膜区(第287~第309位氨基酸)和丝氨酸/苏氨酸结构域(第363~第623位氨基酸)。进一步分析了TaPK3A的进化关系, 与抗病相关S-Locus RLK类的OnN25-1、ZmPK3、TaLRK10; LB RLK类的Sbds1、OsXa26、AtPSKR1; LRR RLK类的NbLRK1、AtLecRK-VI.2、AtLPK1; WAK/WAKL RLK类的AtWAK1、AtWAKL10、OsWAK1; LysM RLK类的AtCERK1、SiBti9、OsCERK1, 使用ClustalW比对, 以MEGA7软件构建系统发生树。结果显示, TaPK3A与S-locus RLK类基因在同一分支, 并且与小麦抗锈病蛋白TaLRK10亲缘关系最近(图2)。TaPK3A与TaLRK10蛋白全长氨基酸序列间一致性为32.32%。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2TaPK3A蛋白的系统进化树分析
用MEGA7对TaPK3A进化树分析, 五角星标注的是TaPK3A蛋白, 竖线内为同一亚家族蛋白。AtCERK1 (NP_566689.2)、AtLecRK-VI.2 (Q9M021.1)、AtLPK1 (NP_567233.1)、AtPSKR1 (OAP07882.1)、AtWAK1 (OAP13193.1)、AtWAKL10 (NP_178086.1)来自拟南芥; OnN25-1 (AHW98543.1)来自野生稻; NbLRK1 (BAG68210.1)来自本氏烟; OsWAK1 (BAG89532.1)、OsCERK1 (A0A0P0XII1.1)、OsXa26 (AAR08150.1)来自水稻; Sbds1 (BAM45642.1)来自高粱; SiBti9 (ADL16642.1)来自谷子; TaLRK10 (AAC49629.1)来自小麦; ZmPK3 (CAA09029.1)来自玉米。
Fig. 2Phylogenetic tree analysis of TaPK3A protein
The phylogenetic tree of TaPK3A was analyzed by MEGA7. TaPK3A protein was tagged with star. The protein in the vertical lines belongs to the same subfamily. AtCERK1 (NP_566689.2), AtLecRK-VI.2 (Q9M021.1), AtLPK1 (NP_567233.1), AtPSKR1 (OAP07882.1), AtWAK1 (OAP13193.1), AtWAKL10 (NP_178086.1) are from Arabidopsis thaliana; OnN25-1 (AHW98543.1) is from Oryza nivara; NbLRK1 (BAG68210.1) is from Nicotiana benthamiana; OsWAK1 (BAG89532.1), OsCERK1 (A0A0P0XII1.1), OsXa26 (AAR08150.1) are from Oryza sativa; Sbds1 (BAM45642.1) is from Sorghum bicolor; SiBti9 (ADL16642.1) is from Setaria italic; TaLRK10 (AAC49629.1) is from Triticum aestivum L.; ZmPK3 (CAA09029.1) is from Zea mays L.
2.2 TaPK3A基因的表达特性分析
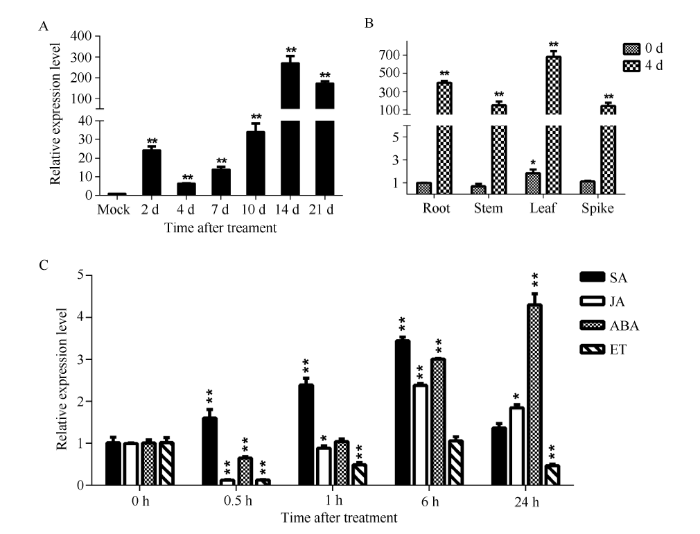
以未接种为对照, 连续检测接种禾谷丝核菌后2~21 d抗性小麦CI12633中TaPK3A的相对表达水平, 发现该基因受到病菌的强烈诱导, 在接种后14 d表达量达到最大值(图3-A), 说明TaPK3A可能在病原菌侵染过程中起作用。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3小麦品系CI12633中TaPK3A基因的表达分析
A: TaPK3A基因在抗纹枯病小麦CI12633中的表达分析, 横坐标表示禾谷丝核菌侵染小麦CI12633后0 h、2 d、4 d、7 d、10 d、14 d和21 d时的样品。B: CI12633小麦不同组织中TaPK3A受禾谷丝核菌的诱导表达分析。C: TaPK3A基因响应 SA、JA、ABA和 ET的表达分析, 横坐标表示不同植物激素处理小麦CI12633后0 h、0.5 h、1 h、6 h和24 h时的样品。t检验分析TaPK3A表达量的差异显著性, *P < 0.05; **P < 0.01。
Fig. 3Expression analysis of TaPK3A in wheat line CI12633
A: expression analysis of TaPK3A in resistant wheat line CI12633. 0 h, 2 d, 4 d, 7 d, 10 d, 14 d, and 21 d stand for the samples of wheat line CI12633 after inoculation of R. cerealis. B: expression analysis of TaPK3A induced by R. cerealis in the organs of wheat line CI12633. C: expression analysis of TaPK3A after SA, JA, ABA, and ET treatment. 0 h, 0.5 h, 1 h, 6 h, and 24 h stand for the samples of wheat line CI12633 after different phytohormones treatments, t-test was used to statistically analyze the significant difference of TaPK3A expression. * P < 0.05; ** P < 0.01.
取未受禾谷丝核菌侵染的小麦和受禾谷丝核菌侵染4 d后的小麦的不同组织, 以未接菌植株根中TaPK3A的表达量为基准, 进行表达模式的分析, 结果显示TaPK3A在小麦的不同组织都有一定程度的表达。在未接种禾谷丝核菌的情况下, 叶中的表达量最高。在接种禾谷丝核菌4 d后, 受病原菌的诱导, TaPK3A在根、茎、叶和穗中的表达量均显著升高, 且叶中的表达量最高(图3-B)。
为了研究TaPK3A基因对不同植物激素的响应, 在三叶一心期对小麦喷施SA、MeJA、ABA和ET, 分析TaPK3A基因在激素处理前(0 h)和处理后0.5~24 h的转录水平。结果显示, 激素处理后, TaPK3A最先受SA诱导表达; 处理6 h后, TaPK3A开始受JA和ABA诱导表达(图3-C)。
2.3 BSMV-VIGS分析TaPK3A基因在小麦抗纹枯病反应中的功能
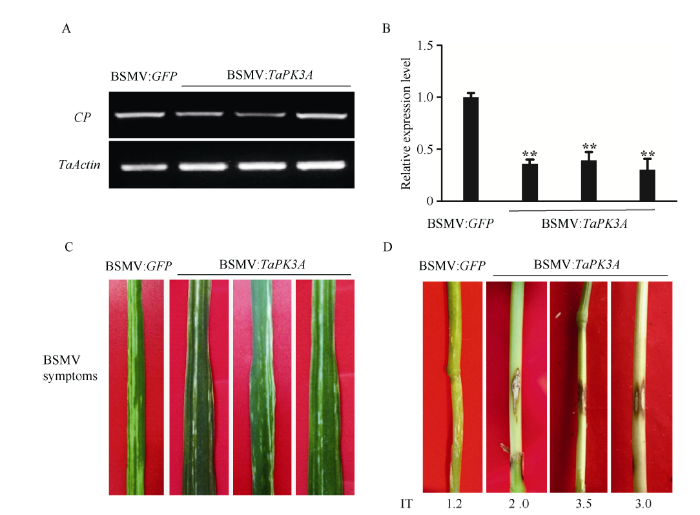
将BSMV:GFP和BSMV:TaPK3A病毒RNAs分别转染小麦品系CI12633叶片。转染病毒后6 d, 在新生叶片能观察到病毒斑, 第10天观察到明显的病毒斑(图4-C)。使用半定量PCR可以检测到编码BSMV外壳蛋白CP基因的转录表达, 说明BSMV病毒成功转染到小麦植株(图4-A)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4BSMV CP、TaPK3A基因的检测及小麦花叶和纹枯病表型
A: 半定量PCR检测CP的转录表达, 扩增26个循环; B: 对接种BSMV: GFP和BSMV: TaPK3A 10 d后的植株RT-qPCR检测TaPK3A的表达, **P < 0.01 (t检验); C: 接种BSMV后10 d, 小麦叶片呈现出BSMV病毒斑; D: 接种禾谷丝核菌后30 d小麦茎秆上纹枯病斑, IT: 病级
Fig. 4Expression analysis of CP and TaPK3A and the symptoms of BSMV and sharp eyespot in BSMV-VIGS wheat
A: transcriptional expression of CP by semi-quantitative PCR, and the amplification was 26 cycles. B: RT-qPCR analysis of the relative transcript levels of TaPK3A in the wheat plants infected by BSMV: GFP or BSMV: TaPK3A at 10 dpi. **P < 0.01 (t-test). C: BSMV virus spots on wheat leaves 10 days later after inoculation. D: sharp eyespot symptoms on wheat stems 30 days after inoculation with R. cerealis. IT: infection type.
为了检测TaPK3A基因的沉默情况, 以接种BSMV:GFP植株为对照, 对接种BSMV:TaPK3A的植株使用RT-qPCR分析TaPK3A的表达量。接种BSMV:TaPK3A的CI12633植株中TaPK3A的表达量显著降低(图4-B), 表明TaPK3A在小麦植株中被沉默。
在转染BSMV:GFP和BSMV:TaPK3A小麦CI12633拔节期, 采用牙签法接种禾谷丝核菌WK207, 分析TaPK3A沉默对小麦纹枯病抗性的影响。接种10 d后, 对照植株和BSMV:TaPK3A转染的CI12633植株的叶鞘出现明显的纹枯病症状; 接种30 d后, 所有CI12633植株茎上都有典型的纹枯病症状, 大多数转染BSMV:TaPK3A的CI12633植株的纹枯病更严重(图4-D)。转染BSMV:GFP的CI12633植株纹枯病平均病级和病情指数分别为1.27和25.40; 转染BSMV:TaPK3A的CI12633植株纹枯病平均病级和病情指数分别为2.31和46.20, 纹枯病平均病级和病情指数均达到极显著差异(表1)。沉默TaPK3A显著减弱了抗纹枯病小麦CI12633抑制禾谷丝核菌侵染的能力, 说明TaPK3A是小麦抗纹枯病反应所需的。
Table 1
表1
表1TaPK3A沉默及对照小麦的纹枯病鉴定
Table 1
| 基因型 Genotype | 病级 Infection type | 病情指数 Disease index |
|---|---|---|
| BSMV:TaPK3A | 2.31** | 46.2** |
| BSMV:GFP | 1.27 | 25.4 |
新窗口打开|下载CSV
3 讨论
本研究根据转录组数据分离克隆了小麦蛋白激酶TaPK3A全长编码序列。TaPK3A在叶中表达量最高, 且其表达受到禾谷丝核菌的诱导而上调。TaPK3A是由660个氨基酸组成的类受体蛋白激酶, 具有信号肽、跨膜域和丝氨酸/苏氨酸结构域。进化树的分析结果表明, TaPK3A蛋白与小麦的TaLRK10、玉米的ZmPK3及野生稻的OnN25-1形成一个独立的S-Locus RLK分支。已有研究表明, 小麦TaLRK10具有抗叶锈病的作用[26], 且TaPK3A与其亲缘关系最近, 但是TaPK3A与TaLRK10的全长氨基酸序列间一致性为32.32%, 说明TaPK3A是一个新的RLK功能成员。植物类受体蛋白激酶在抗病虫方面发挥着重要的作用[22-27,32-33]。S结构域RLK能引起自交不亲和性, 部分S结构域RLK能识别病原菌、病原相关分子模式和损伤[32]。Feuillet等[26]、Zhou等[27]和Brueggeman等[33]研究表明, RLK参与小麦及大麦的抗锈病反应。本研究表明, TaPK3A受禾谷丝核菌的强烈诱导, 且在接种纹枯病菌后14 d达到高峰, 暗示TaPK3A参与小麦抗纹枯病全过程。利用BSMV病毒介导的VIGS技术, 可以在大麦和小麦植物中实施高度同源的内源基因沉默, 是快速分析大麦和小麦基因功能的一种有效方法[27,29-30,34-36]。利用BSMV-VIGS沉默技术也已证明, TaBON1和TaBON3负调控小麦抗白粉病反应[37], 小麦RAR1和SGT1 在小麦抗叶锈病基因Lr21介导的免疫反应中发挥作用[38], TaAGC1和TaRCR1正向调控小麦对禾谷丝核菌的抗性反应[30,39]。本研究中, 沉默TaPK3A显著减弱了小麦品系CI12633对纹枯病的抗性, 表明TaPK3A是小麦抗纹枯病反应所需的1个基因。目前, 关于S结构域RLK在植物抗病中作用的研究相对较少, 本研究丰富了植物RLK在防御病原菌中的理解。
植物激素不仅影响植物的生长发育, 同时也在植物对病原的防御过程中起重要的作用, 植物防御激素水杨酸(SA)、茉莉酸(JA)和乙烯(ET)介导的信号途径, 是植物抗病反应的重要信号传导通路[40]。不同的植物激素在防御不同营养型的病原菌过程中起到不同的作用, SA主要在植物对活体营养型或者半活体营养型的病原菌的抗性中起主要作用, 而JA和ET主要在抗腐生营养型病原菌和虫害过程中起作用[41]。Chen等[42]发现抗白粉病相关基因TaRLK1和TaRLK2通过SA和ROS进行信号传导。本研究中, 在施加激素的前期, TaPK3A快速响应外源SA刺激而上调表达, 推测TaPK3A主要通过SA信号途径传导。在施加激素6 h后, TaPK3A同样受到JA和ABA的诱导表达, 表明TaPK3A在传递病原菌侵入信号的过程较为复杂, 需要后续的研究。值得一提的是, R. cerealis被认为是一种腐生营养型病原菌, 而本研究表明TaPK3A可能通过SA途径传递信号参与小麦对纹枯病的防御反应, 这丰富了对抗病信号传递的理解。
4 结论
克隆出一个小麦RLK基因TaPK3A, 其编码660个氨基酸组成的类受体蛋白激酶TaPK3A; TaPK3A在小麦中表达受禾谷丝核菌和外源SA的诱导而上调; 沉默TaPK3A降低了小麦对纹枯病的抗性, 表明TaPK3A是小麦防御纹枯病菌反应所需的基因。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1038/nature05286 [本文引用: 1]
DOI:10.1016/S1369-5266(03)00055-4URL [本文引用: 1]
[本文引用: 2]
DOI:10.1007/BF00016492URL [本文引用: 2]
DOI:10.1093/jxb/erp233URL [本文引用: 1]
.
[本文引用: 1]
[本文引用: 2]
DOI:10.1104/pp.011028URL [本文引用: 1]
[本文引用: 1]
DOI:10.1126/science.273.5280.1406URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1000675107URL [本文引用: 1]
DOI:10.1111/tpj.2009.60.issue-6URL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1111/tpj.2006.46.issue-5URL [本文引用: 1]
DOI:10.1038/nbt.3069URL [本文引用: 1]
URL [本文引用: 3]
DOI:10.1111/j.1365-313X.2007.03246.xURL [本文引用: 4]
DOI:10.1006/meth.2001.1262URL [本文引用: 1]
[本文引用: 2]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.2008.178.issue-3URL [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
DOI:10.1104/pp.15.01784URL [本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/1369-5266(88)80053-0URL [本文引用: 1]
DOI:10.1146/annurev.phyto.43.040204.135923URL [本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}